











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkBotanical excursions over several years into central Veracruz along the transition zone between cloud forest and tropical forest at around 800-1,800 m elevation have resulted in the location of a number of Ceratozamia Brongn. populations. The taxonomic status of one of these cycads, C. brevifrons Miq. that was considered synonymous with C. mexicana Brongn. has been solved by applying numerical analyses to leaflet anatomical characters by Vovides et al. (2012). The objective of this study was to elucidate the taxonomic status of the cycads in two of these populations considered as C. mexicana that separated significantly in the analyses; at El Esquilón north of the city of Xalapa and at El Mirador to the north east of Huatusco. Details of the other taxa studied, apart from one appearing in the analysis will not be considered further here and can be consulted in Vovides et al. (2012). Living specimens were collected and cultivated at the Jardín Botánico Francisco Javier Clavijero (JBC) for several years in order to observe plasticity upon later comparison with habitat plants. Herbarium specimens at XAL, MEXU, NY and especially the type of C. mexicana at the herbarium of the National Museum of Natural History, Paris (P) were also examined and compared. Table 1 lists the species and vouchers of the specimens used for the anatomical study and analysis deposited at XAL, MEXU and NY.
Table 1 El Esquilón and El Mirador, Veracruz populations of Ceratozamia mexicana vouchers deposited at XAL of the specimens used for the anatomical analysis.

Brongniart (1846) described Ceratozamia mexicana from a cultivated plant in Paris as having coriaceous, oblong-lanceolate, subfalcate, entire, acuminate smooth leaflets with partly sunken parallel veins that corresponds to the holotype in Paris (P!, Figure 1A) Brongniart s.n. This was collected by Ghiesbrecht in Mexico from El Mirador (Schuster, 1932) also mentioned by Thiselton-Dyer (1884), and an illustration in Brongniart (1846, Figure 1B), see also Stevenson and Sabato (1986). Thiselton-Dyer (1884) described C. mexicana var. tenuis Dyer very briefly with no illustration referring to Mexico as the provenance, but at the time of typification by Stevenson and Sabato (1986) no extant specimens were known so they typified with the description (no longer legal in the International Code of Nomenclature for algae, fungi, and plants). Thus, the true nature of this taxon was enigmatic until Stevenson in 1999 discovered oversized sheets of cycads at the Kew herbarium that were far removed from the normal-sized collections. In those collections are three sheets of a single leaf of a specimen labelled C. mexicana var. tenuis collected from a cultivated plant at Kew (no longer alive) in 1881 (K-3 sheets!, Figure 2) that we consider to be the lectotype. Thus, with an established type, we have a better concept of the morphology of this taxon. Coupled with this discovery, Vovides and Avendaño visited the El Mirador population northeast of Huatusco and it does correspond to the holotype of C. mexicana; thus, we refer to this as the El Mirador population. Chamberlain (1919) located a population about 11 km north of the city of Xalapa in a broad valley on the road to Naolinco that he considered C. mexicana, and subsequently was recognized as C. mexicana var. mexicana in the Flora of Veracruz by Vovides et al. (1983) and C. mexicana by Whitelock (2002). There are still stands of this cycad in this area in localities such as El Esquilón, and it is more abundant in the surrounding forests near Coacoatzintla. Ceratozamia mexicana sensu Chamberlain, which we refer to as the El Esquilón population, differs from the type specimen (El Mirador) in that the El Esquilón population has linear lanceolate, falcate to subfalcate leaflets whereas the leaflets of the type specimen of C. mexicana are oblong-lanceolate, subfalcate (Brongniart, 1846; Figure 1). The Esquilón population has leaflet morphology consistent with the cultivated plant of C. mexicana var. tenuis as represented by the lectotype (Figure 2). Thiselton-Dyer (1884) comments that C. mexicana var. tenuis leaflets resemble those of C. kuesteriana Regel, but the general habit is different. Ceratozamia kuesteriana has consistently reddish brown emergent leaves (a character that is lost on herbarium vouchers), elongate linear-lanceolate leaflets that are strongly caniculate adaxially, mentioned in the German description shown in Figure 22 of Tafel 4 by Regel (1857a) and can be seen in the illustration in Figure 2 of Moretti et al., (1982), in Figure 2 of Vovides (1985), and most obviously in Figures 24-25 Tafel 186 of Regel (1857b). Leaflets in C. mexicana from El Mirador are consistently reddish brown emergent and flat whereas those in C. mexicana from El Esquilón are consistently light green emergent and only slightly caniculate.
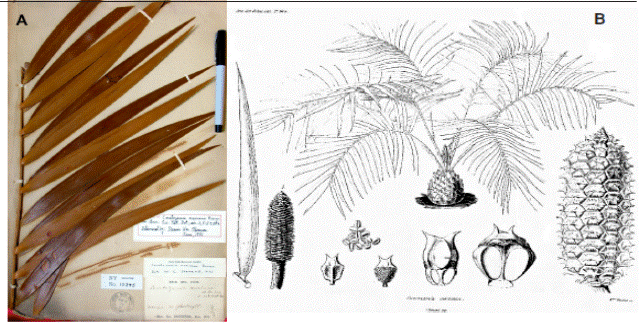
Figure 1 Ceratozamia mexicana. A) Holotype of C. mexicanaBrongniart. B) Illustration from Brongniart’s original 1846 paper describing C. mexicana.

Figure 2 Holotype of Ceratozamia mexicana var. tenuis. A) Sheet 2 of 3. B) Sheet 3 of 3, insert above, enlarged portion of text at base of Sheet 3. C) Sheet 1 of 3. D) Enlargement of letter attached to Sheet 2 in Thiselton-Dyer’s handwriting.
Methods for anatomical study
Leaflets for used for sectioning were taken from plants growing in habitat as well as those cultivated in the Jardín Botánico Francisco Javier Clavijero over several years. The central portion of leaflets taken from the median leaflets of leaves from five individuals of each taxon from each of the populations were fixed in FAA and hand sectioned with the aid of a sliding microtome. Fixed sections were subjected to the phloroglucinol HCl test for lignin also permanent preparations were made, double-stained with safranin and fast green, cleared and mounted according to Purvis et al. (1966). Photomicrographs taken with a digital camera using clear field light microscopy. Cross-sectional measurements were taken from 11 characters (Table 2) using a calibrated eyepiece micrometer; where cells were not isodiametric the measurements were expressed as length and width. Twentyfive replicate measurements were taken for each character from each of the five leaflet samples of the two taxa as described in Vovides et al. (2012), the additional species C. robusta Miq. was included order to satisfy requirements for a discriminant re-analysis of the data on three taxa. Analysis of variance (ANOVA) and Tukey multiple range analyses were done using JMP version 3.2 statistical software. Discriminant analyses (McCune & Mefford, 1997) were done with Statgraphics software version 2.0. Data were transformed to log10 and Mahalanobis distances were obtained.

Results
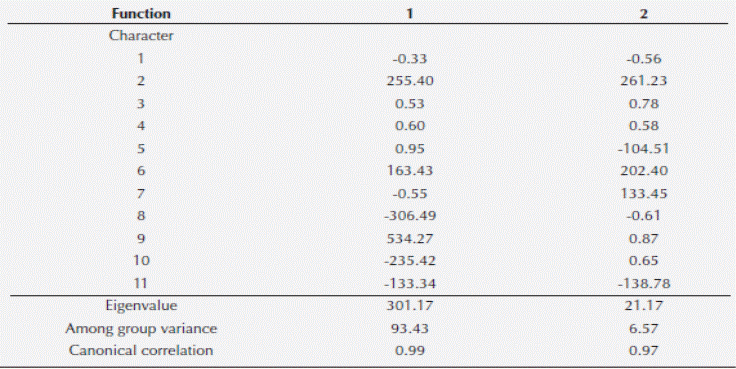
Two qualitative morphological and one anatomical variable separate Ceratozamia mexicana, El Mirador and C. mexicana, El Esquilón (Table 3). Furthermore the ANOVA test on the11 anatomical variables of the leaflets of the Ceratozamia species tested were significantly different at P < 0.001 (Table 4) except for macrolumen epidermal cell and palisade parenchyma cell length (characters 2 and 5 respectively). The Tukey multiple range test showed that the number of perivascular fibres (character 10) were significantly different for the two C. mexicana populations, separating the groups with no overlap. Ceratozamia robusta is separated from the two C. mexicana populations by the number of intervascular fibres character (character 11). Data derived from the discriminant function analysis of anatomical characters separate the three populations; two of C. mexicana (Esquilón and El Mirador populations) and C. robusta coordinately in bi-dimensional space and do not present any overlapping between groups (Figure 3). The Wilks Lambda (λ) test was highly significant (Wilk’s λ = 0.00015 P < 0.0091) (Table 5), thus showing that all the species were classified correctly. Additionally, the squared Mahalanobis distances (Table 6) were statistically significant in all cases (P < 0.05). Of the 11 variables included in the standardized discrete canonical function, the two variables with the highest values in discriminant functions 1 and 2 were the cross sectional length of the thin walled macrolumen epidermal cells (character 2) and the cross sectional width of intervascular fibers (character 9) for function 1. The first and second canonic variable showed that 93.43 % of the variation is largely due to anatomical characters. The positive correlations (Table 7) of all the variables show differences between species. Additional to this analysis microscopic examination of leaflet cross sections revealed that C. mexicana (El Mirador) presents incipient girder sclerenchyma (not used in analysis) whereas those of El Esquilón do not (Figure 4).

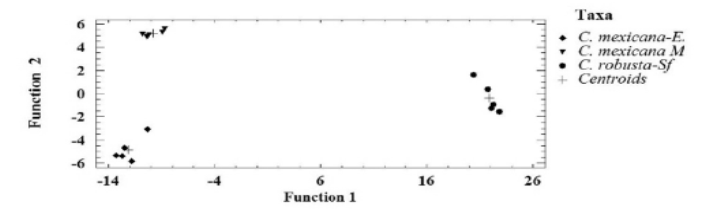
Figure 3 Scatter plot score derived from the functions produced by stepwise discriminant analysis of 11 anatomical characters occurring in Ceratozamia tenuis from El Esquilón, C. mexicana from El Mirador, and C. robusta from San Fernando, Chiapas.
Table 4 Summary of analysis of variance of the 11 anatomical characters R2 (correlation coefficient), F (F value), P (Probability).
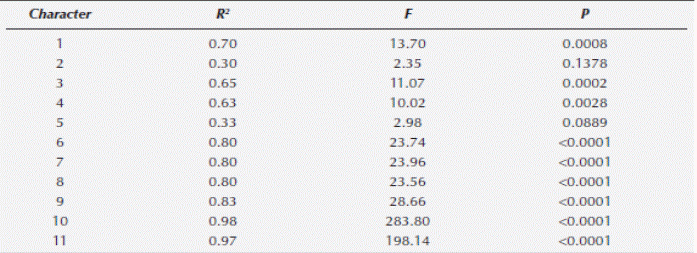
Table 5 Summary of the discriminant analysis results from the analysis of three taxa in Ceratozamia χ2 = Chi square test, DF: Degrees of freedom, P = Probability
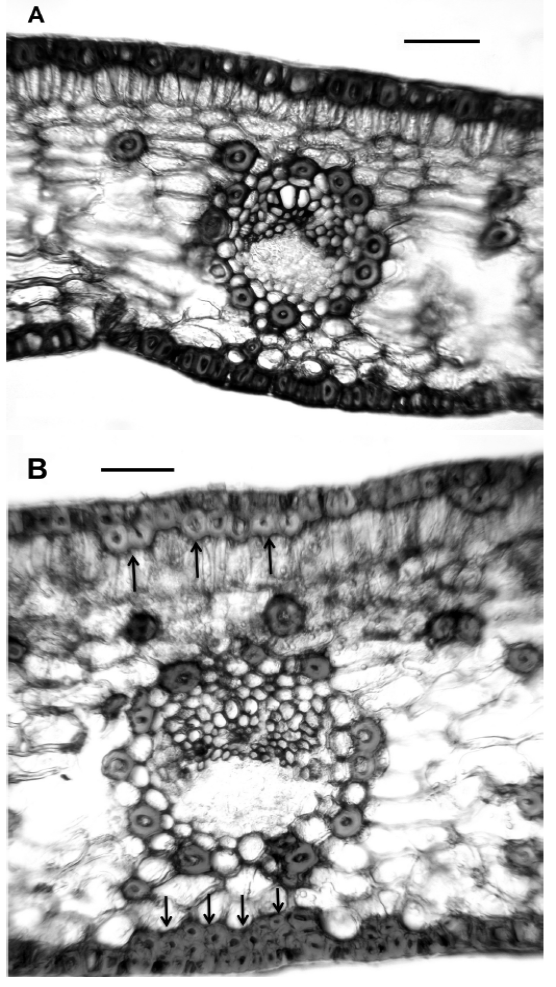
Figure 4 Transverse sections of leaflets treated with phloroglucinol HCl. A) Ceratozamia tenuis from El Esquilón. B) Ceratozamia mexicana from El Mirador with girder sclerenchyma (arrows). Scale Bars = 100 μm.
Table 6 Summary of the squared Mahalanobis distance (above asterisk), F values (below asterisk) and statistical significance (P). Esq = Esquilón population, Mir = Mirador population. P < 0.05 for all cases.
Table 7 Standardized discriminant function values for each of three factors used in the analysis of taxa for 11 leaflet anatomic characters.
We therefore consider the plants from El Esquilón to match Thiselton-Dyer’s concept of Ceratozamia mexicana var. tenuis, and consider this taxon distinct enough to merit species rank. In following the guidelines for cycad classification proposed in Walters and Osborne (2004) that discourages the use of infraspecific units, we propose a change in rank of C. mexicana var. tenuis to C. tenuis, and based upon handwriting and origin a lectotype was assigned for this species. Epitypes were assigned for C. tenuis (AV-018) and for C. mexicana [JR-1688 (XAL, MEXU, NY)].
Description
Ceratozamia tenuis (Dyer) D. W. Stev. et Vovides stat. and comb. nov. (Figure 5A) Basionym: Ceratozamia mexicana var. tenuis Dyer Biol. Cent.-Amer., Bot. 3: 193. 1884. Type: Hort. Kew Palm House Thistleton-Dyer s.n. 1881 (Lectotype, here designated, K! 3 sheets). Epitype, here designated A. Vovides 018 (XAL, NY).

Trunk epigeous, globose becoming cylindrical with age, up to one meter tall or more in very old plants, up to 20 cm diameter or more when mature. Leaves 1-15 forming a terminal crown, pinnate, 80-280 cm long, 50-100 cm wide; leaflets 20-30 pairs, linear-lanceolate to lanceolate, falcate to subfalcate, 22-50 cm long, 1.3-1.8 cm wide but up to 2.5 cm or more wide in juveniles, apex acuminate to sub-acuminate, base attenuate; petiole and rachis ascending to descending, armed with short to long stout prickles. Pollen strobili cylindrical to narrowly conical, erect, 17-25 cm long or more, 2.5-4.5 cm diameter; peduncle short, tomentose; ovulate strobili olive green, cylindrical, 10-31 cm long, 5-10 cm diameter; peduncles 9-13 cm long, 1-2 cm diameter, erect, tomentose. Seeds angular to ovoid, 1.7-2 cm long, 1.4-1.6 cm diameter, sarcotesta fleshy, creamy white turning brown at maturity. Chromosome number 2n = 16 (this study).
Other vouchers examined:
MEXICO: Veracruz; Cuacuatzintla, Castillo-Campos 118 (XAL); Naolinco, Chamberlain 13 (F); Chiconquiaco, Hernández 385A, 1507 (MEXU, XAL); Jilotepec, El Esquilón Ortega 525 (MEXU, XAL), El Esquilón Rees 1620 (XAL), El Esquilón A. Vovides 018, 470, 471, 635, 735 (XAL).
Ceratozamia mexicana Brong. (Figure 5B) Ann. Sci. Nat. Bot., ser. 3, 5: 7-8, Tab. 1. 1846.Type: Mexico, from Ghiesbrecht, cult. in Hort. Bot. Parisiensis, 1845, Brongniart s.n. (Holotype, P!). Epitype (here designated): J. Rees 1688 (XAL).
Trunk partially subterranean to epigeous, globose to ovoid, becoming cylindrical with age, up to one meter tall or more in very old plants, up to 35 cm diameter or more when mature. Leaves 1-16 or more forming a terminal crown, pinnate, 80-300 cm long, 50-90 cm wide; leaflets 20-50 pairs or more, leaflets oblong-lanceolate, subfalcate, entire, acuminate, smooth 25.8-45.5 cm long, 2-3.4 cm wide, apex acuminate, base attenuate; petiole and rachis ascending to descending, armed with short to long stout prickles. Pollen strobili cylindrical to narrowly conical, erect, 17-25 cm long or more, 2.5-4.5 cm diameter; peduncle short, tomentose; ovulate strobili olive green, cylindrical, 20-45 cm long, 5-10 cm diameter; peduncles becoming decumbent when ovulate strobilus matures, 9-13 cm long, 1.5-2.5 cm diameter, tomentose. Seeds angular to ovoid, 1.7-2 cm long, 1.4-1.6 cm diameter, sarcotesta fleshy, creamy white turning brown at maturity. Chromosome number 2n = 16 (this study).
Other vouchers examined:
MEXICO: Veracruz; Totutla, El Mirador A. Vovides 730, 731, 732, 733 (XAL), Walters 2-1, 2-3, 2-5 (XAL, FTG); Totutla, Zacuapan C. A. Purpus 6362 (XAL).
Key to species:
Leaflets linear lanceolate to lanceolate, shallowly caniculate, 1.3-1.8 cm wide…....….C. tenuis
Leaflets oblong lanceolate, flat, 2-3.4 cm wide……………..…………………...…C. mexicana
Discussion
Recent botanical explorations at mid elevations (800 to 1,800 masl) in central Veracruz on the Transvolcanic Mexican Belt revealed populations of related Ceratozamia spp. including C. tenuis that we consider to form part of the C. mexicana species complex that is comprised of C. morettii Vázq. Torres and Vovides, C. brevifrons Miq., C. decumbens Vovides, Avendaño, Pérez-Farr.& Gonz.-Astorga, C. delucana Vázq.Torres, A.Moretti & Carvajal-Hern. Ceratozamia tenuis differs from its congeners within the complex by having long, narrow, linear-lanceolate, slightly caniculate leaflets and occurs in small, restricted populations in the Xalapa region, although plants may be locally abundant within these populations. Molecular phylogenetic studies by González and Vovides (2002, 2012) and an overall review by Vovides et al. (2004) reported that the C. mexicana species complex may be of recent speciation along with the C. kuesteriana species complex in northeastern México above the Transvolcanic Mexican Belt also known as the Mexican transition zone (Contreras-Medina et al., 2007). Molecular phylogenetic relationships among closely related plant species are problematic, both nuclear ribosomal DNA ITS and chloroplast DNA trnL-F non-coding region (González and Vovides, 2002) and a modification to the Sequence Characterized Amplified Region (SCAR) method (González and Vovides, 2012) on the genus Ceratozamia showed a similar and unresolved topology in the clade grouping both the C. mexicana and the C. zaragozae species complexes at and to the north of the Mexican Transvolcanic Mexican Belt respectively. However the morphological and anatomical data were found to be useful in elucidating the species, rather than the molecular studies alone that showed clear geographical differences but not interspecific. That modern cycad species are of recent speciation (Miocene) has been demonstrated by Morettii et al. (1993) and Nagalingum et al. (2011) and is in agreement with our hypothesis (González and Vovides, 2002, 2012). Furthermore, we regard populations of C. mexicana, C. tenuis, C. morettii, and C. brevifrons as forming the Ceratozamia mexicana species complex that may be of hybrid origin similar to that found by Johnson (1963) for the Macrozamia communis L.A.S. Johnson complex. Further research combining morphological, anatomical and molecular data may throw more light on the speciation processes in Ceratozamia.

