










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBotanical Sciences
versão On-line ISSN 2007-4476versão impressa ISSN 2007-4298
Bot. sci vol.91 no.2 México Jun. 2013
Botánica estructural
Karyotype characterization of eight Mexican species of Eleocharis (Cyperaceae)
Caracterización de los cariotipos de ocho especies mexicanas de Eleocharis (Cyperaceae)
Jorge Alberto Tena-Flores1,4, María Socorro González-Elizondo1, Yolanda Herrera-Arrieta1, Norma Almaraz-Abarca1, Netzahualcóyotl Mayek-Pérez2, Carlos Roberto Maximiano Da Silva3 and André Luís Laforga Vanzela3
1 Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional (CIIDIR) Durango, Instituto Politécnico Nacional, Durango, Durango, México.
2 Centro de Biotecnología Genómica, Instituto Politécnico Nacional, Reynosa, Tamaulipas, México.
3 Laboratório de Biodiversidade e Restauração de Ecossistemas, Departamento de Biologia Geral, CCB, Universidade Estadual de Londrina, Londrina, Paraná, Brasil.
4 Autor para correspondencia: jorge@tena-flores.com
Received: March 15th, 2012
Accepted: June 18th, 2012
Abstract
Karyotypes of 49 populations belonging to eight species of Mexican Eleocharis (Cyperaceae) are described. Chromosome numbers for Eleocharis densa, E. reznicekii, and E. rostellata are reported for the first time and new numbers are reported for E. macrostachya, E. xyridiformis, and the E. montevidensis complex. Numbers ranged from 2n = 10 to 2n = 60. Dysploidy was the most common mechanism of karyotype variation, which has been detected in four species (E. densa, E. macrostachya, E. reznicekii, and E. xyridiformis). Two species are diploid (Eleocharis parishii and E. cf. montevidensis) and three are polyploid (E. acicularis, E. montevidensis, and E. rostellata). Except for specimens of E. montevidensis complex, no intraspecific variation in chromosome number was found. However, differences in the chromosome sizes were found among populations of that complex and in E. rostellata. Mean lengths of diploid set ranged from 12.96 μm in E. montevidensis to 178.25 μm in E. rostellata and the average of chromosomes sizes varied from 0.97 μm in E. montevidensis to 6.01 μm in E. xyridiformis. These two taxa presented an extreme interchromosomal asymmetry A2: 0.12 and 0.43. Absence of primary constrictions was confirmed. Taxonomical implications of the karyological data are discussed.
Key words: chromosome, cytotaxonomy, dysploidy, holocentric, sedges.
Resumen
Se describen los cariotipos de 49 poblaciones de ocho especies de Eleocharis (Cyperaceae) de México. Se reportan por primera vez números cromosómicos para Eleocharis densa, E. reznicekii y E. rostellata, así como nuevos números para E. macrostachya, E. xyridiformis y plantas del complejo de E. montevidensis. Los números cromosómicos van de 2n = 10 a 2n = 60. El mecanismo más común de variación cariotípica es la disploidía, presente en la mitad de las especies (E. densa, E. macrostachya, E. reznicekii y E. xyridiformis). Dos especies son diploides (E. parishii y E. cf. montevidensis) y tres son poliploides (E. acicularis, E. montevidensis y E. rostellata). No se encontró variación intraespecífica en cuanto a números cromosómicos excepto para plantas del complejo de E. montevidensis, pero se encontraron diferencias en tamaño entre poblaciones de ese mismo complejo y en E. rostellata. Las longitudes medias del cariotipo van de 12.96 a 178.25 μm (en una variante de E. montevidensis y en E. rostellata, respectivamente); los promedios de longitud de los cromosomas van de 0.97 μm en E. montevidensis a 6.01 μm en E. xyridiformis, especies que también presentan los extremos de asimetría intercromosomal A2: 0.12 y 0.43, respectivamente. Se confirma la ausencia de constricciones primarias. Se discuten las implicaciones taxonómicas de los datos cariológicos.
Palabras clave: ciperáceas, citotaxonomía, cromosoma, disploidía, holocéntrico.
Cyperaceae is the third largest family of monocots, with about 5,400 species in 106 genera (Govaerts et al., 2007) to 5,500 species in 109 genera (Muasya et al., 2009). Known as sedges, its representatives occur in a variety of habitats, being most common in moist areas. It includes several worldwide distributed genera, e.g., Carex L., with more than 2,000 species (Reznicek, 1990) and Eleocharis R.Br., which comprises more than 270 species (González-Elizondo, unpubl. data). Many species of Eleocharis are important forage for livestock, a few are used as human food, and several have potential use in aquatic weed management and in pollution abatement (Catling and Hay, 1993). The use of native plants that have a high capacity to accumulate metals and remove them from soil and water (phytoremediation) is a very convenient approach (González-Elizondo et al., 2005).
Eleocharis is distinguished by having unbranched stems, leaves reduced to basal, tubular sheaths, inflorescence reduced to a simple terminal spikelet, floral traits very reduced in number and size, and achene with a persistent stylobase (González-Elizondo and Peterson, 1997; González-Elizondo and Tena-Flores, 2000). In spite of the easy recognition of Eleocharis as a genus and its prominent delimitation within Cyperaceae (Kukkonen, 1990), the species are difficult to identify and classify because of the limited number of morphological features (Smith et al., 2002). This limitation is increased because several traits are strongly variable among related species and, in some cases, the convergence is common and many morphological features are not phylogenetically informative (González-Elizondo and Tena-Flores, 2000). The supraspecific classification of Eleocharis has been revised and modified by Kukkonen (1990) and González-Elizondo and Peterson (1997) on the basis of the classification of Svenson (1929). Four subgenera are recognized but recent phylogenies (Roalson and Friar, 2000; Roalson et al., 2010) reveal that subgenus Eleocharis is paraphyletic.
Cyperaceae possess a unique combination of cytogenetical features: holokinetic ("holocentric") chromosomes, possibility of inverted meiosis, and pseudomonad development (asymmetric tetrads). Holokinetic condition favors karyotype differentiation for agmatoploidy (fission), symploidy (fusion), and polyploidy (Luceño and Guerra, 1996; Da Silva et al., 2005, 2008b).
The ability of holokinetic chromosomes to migrate parallel in the cell divisions is due to kinetic activity distributed throughout the chromosome, which favors the maintenance of chromosome rearrangements, such as fissions and fusions, and chromosomes viability after most rearrangements. For this reason, chromosomes evolve more dynamically in sedges than in any other group of flowering plants (Hipp et al., 2009).
Since holokinetic chromosomes have no primary constriction, the main feature useful to identify the chromosome morphology, the options for analysis based on morphology of the karyotype are greatly reduced in sedges. Because the lack of centromere, karyotype parameters, such as intrachromosomal asymmetry, can only be estimated using the chromosome length. However, the high variation in chromosome number, the interchromosomal asymmetry index, and the presence or absence of nucleolar constrictions had been useful for cytotaxonomical diagnosis (e.g., Da Silva et al., 2008b, 2010).
The use of karyological data in taxonomy, traditionally referred to as cytotaxonomy or karyosystematics, contributes to evaluate the genetic relationships among species or populations and to a better understanding of the way they diverged from each other (Guerra, 2008). The importance of cytogenetical studies to contribute to the knowledge of Mexican plants has been addressed by Palomino (2000). Some examples of karyotypical and cytogenetical analyses of Mexican plants are those of Flores-Maya et al. (2010), Martínez and Palomino (1996), Mercado et al. (1989), Mercado-Ruaro y Delgado-Salinas (1998, 2000, 2009), Palomino and Heras (2001), Tapia-Pastrana and Gómez-Acevedo (2005), Tapia-Pastrana (2010), Tapia-Pastrana and Jiménez-Salazar (2011), and Tapia-Pastrana et al. (2004, 2012). Chromosomes of Mexican sedges are almost entirely unknown. Chromosome numbers have been reported for Carex peucophila Holm (Beaman et al., 1962) and Fimbristylis mexicana Palla (Kral, 1971).
A considerable variation in the chromosome number in Cyperaceae has been recorded, from 2n = 4 for Rhynchospora tenuis (Vanzela et al., 1996) to 2n = 216 for Eleocharis dulcis (Roalson, 2008). Chromosome counting in Eleocharis palustris dates from 1924, by Piech (Da Silva, 2010) and cytological studies in the genus have been made by Strandhede (1965a,b, 1967), Hoshino et al. (2000), Bureš et al. (2004), Yano et al. (2004), and Da Silva et al. (2005, 2008a, b, 2010), among others. However, most members of the family (about 84%) remain cytologically unexplored (Roalson, 2008). The aim of this study was to contribute to the knowledge of Mexican Eleocharis. The karyotypes of 49 populations of eight species of Eleocharis from north-central Mexico were analyzed and the taxonomical implications of the karyological data are discussed.
Materials and methods
Mitotic metaphase chromosomes were studied from root meristematic cells. Forty-nine samples representing eight species of Eleocharis were collected in 35 different localities of north-central Mexico, most of them in the state of Durango. Voucher specimens were deposited in herbarium CIIDIR (Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Instituto Politécnico Nacional). Data of the studied taxa, localities of collection and voucher specimens are given in Table 1. Plants were raised in the nursery of CIIDIR in Durango, Mexico, from materials transplanted from the wild. Root-tips were taken from actively growing plants. The root tips were pre-treated in 2mM 8-hydroxyquinoline for 24 h, fixed in ethanol : acetic acid (3:1, v:v) for 24 h, and stored at -20 °C or immediately used. For the conventional staining, the root tips were softened in 4% cellulase plus 40% pectinase at 37 °C for 1 h, hydrolyzed in 1M HCl for 10 min at 60 °C, and squashed in a drop of 45% acetic acid. Slides were stained in 4% hematoxylin and mounted with Entellan (Merck). Chromosome counts were made in at least 20 cells per sample. Chromosome measurements were made using the freeware computer application MicroMeasure version 3.3. For each sample, from 5 to 10 metaphase spreads with similar condensation were measured. Mean lengths of the karyotype (the total diploid length) and of the shortest and longest chromosome of the complement were calculated. Slides were acquired with a microscope (Carl Zeiss AxioImager.Z2) equipped with an Axiocam Hrc camera, objective Plan-Apochromat 100x/1.4 Oil, and AxioVs40 Rel.4.8.2 software. Ideograms were draw from the chromosome measurements. Interchromosomal asymmetry was calculated using the Romero (1986) index based on Pearson's dispersion coefficient (the ratio between the standard deviation and the mean of chromosome length for each sample): A2 = s /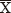 .
.
Results
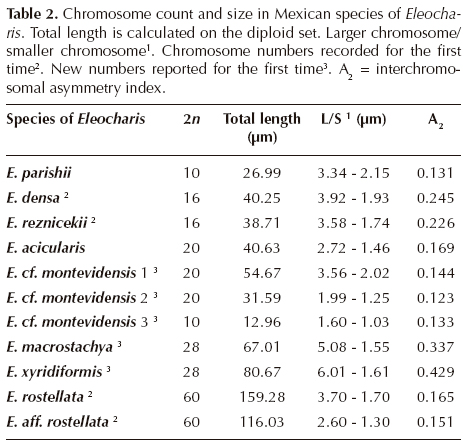
Karyotypes of eight species of Eleocharis from Mexico present holokinetic chromosomes, without primary constactions (Figure 1), and only nucleolar constrictions were observed, as in E. densa (Figure 1B). Chromosome number, mean of diploid set length, the highest and the lowest average chromosome length, and interchromosomal asymmetry index are presented in Table 2. The chromosome number is registered here for the first time for E. densa (2n = 16), E. reznicekii (2n = 16), E. rostellata (2n = 60), and a variant of E. montevidensis (E. cf. montevidensis 3) (2n = 10). New numbers are reported for E. macrostachya (2n = 28) and E. xyridiformis (2n = 28). Numbers range from 2n = 10 to 2n = 60 (Figure 1). Mean length of the karyotype (diploid set) ranges from 12.96 μm (E. cf. montevidensis 3) to 178.24 μm (E. rostellata). The lowest average chromosome length was 1.03 μm (E. cf. montevidensis 3) and the highest 6.01 μm for E. xyridiformis (Table 2).
Ideograms of the haploid complement showed variable karyotypes in which most species exhibit chromosomes decreasing gradually in size, independently of the chromosome numbers (Figure 2). Karyotypes of four species have a low A2 interchromosomal asymmetry index (< 0.17), with chromosomes of about the same size, decreasing gradually. Eleocharis densa and E. reznicekii showed A2 = 0.23-0.25, with two medium and six gradually decreasing small pairs of chromosomes, whereas E. macrostachya and E. xyridiformis present a higher A2 (0.34-0.43), with two large and twelve pairs of medium to small chromosomes gradually decreasing.
Discussion
Karyotypes differ among the studied species in a combination of traits including chromosome number, total length of the diploid set, chromosomes length, asymmetry indices, and mechanisms of variation.
Chromosome numbers. A wide range of chromosome numbers have been registered for Eleocharis, from 2n = 6 for E. subarticulata (Da Silva et al., 2005) to 2n = 216 for E. dulcis (Roalson, 2008). In this study, chromosome numbers ranged from 2n = 10 to 2n = 60, a broad spectrum considering the low number of taxa analyzed (Table 2). Results in this work, with chromosome numbers that are multiples of 5 or that suggesting dysploidy from multiples of that number, are consistent with x = 5 as the basic number for Eleocharis.
For the first time, numbers are given for Eleocharis densa, E. reznicekii, E. rostellata, and a variant of E. montevidensis (E. cf. montevidensis 3). New numbers are reported for E. macrostachya and E. xyridiformis, as well as for plants of the E. montevidensis complex. Chromosomal counts confirm previous reports for E. acicularis with 2n = 20 (Yano et al., 2004; Roalson, 2008), as well as 2n = 20 for E. montevidensis (Roalson, 2008), and 2n = 10 for E. parishii (Roalson, 2008).
Karyotype length. Chromosomes exhibited from small to medium-small sizes, considering the Stebbins (1938) standards, ranging from 1.03 to 6.01 μm, with only two medium-large pairs (> 5 μm) in Eleocharis xyridiformis and one medium-large pair in E. macrostachya. In these two hexaploids with 2n = 28 the medium-large chromosomes could have been originated by fusion of four chromosomes (two-by-two) from the original 2n = 30. The smallest chromosomes were found in the diploid E. cf. montevidensis 3, with 2n = 10. A more fine division in the small chromosomes category for Eleocharis was proposed by Yano et al. (2004), who recognized very small (< 1.1 μm) and larger (1.4-4.3 μm) chromosomes. The first case applies to species of the section Limnochloa and the second one to species of sections Pauciflorae and Eleocharis [subgenera Zinserlingia p.p. and Eleocharis according González-Elizondo and Peterson, 1997]. Very small and numerous chromosomes distinguishing Limnochloa have been confirmed by Da Silva et al. (2008b, 2010). In the present study no species of Limnochloa were included, but some chromosomes lower than 1.4 μm are present in E. montevidensis s.l. and in E. aff. rostellata, species that belong to the subgenus Eleocharis.
Interchromosomal asymmetry A2 (Romero, 1986) describes the variation in chromosome length in a complement. In general terms, the karyotypes found in this study have a low interchromosomal A2 index, with chromosomes decreasing gradually in size (Figure 2, Table 2). Eleocharis densa and E. reznicekii with 2n = 16 could be considered dysploids in relation to basic number x = 5 (see Da Silva et al., 2008b, 2010) with six small and two small-medium pairs of chromosomes (A2 = 0.23-0.25), a karyotype that seems derived by chromosome fusions from a 2n = 20 (Figure 2B, C). Comparatively, the most asymmetric karyotypes, with the highest difference between the longest and the shortest chromosomes were found in E. xyridiformis (6.01/1.61) and E. macrostachya (5.08/1.55), with A2 indices of 0.43 and 0.34, respectively (Table 2). No association between total karyotype length and asymmetry was found.
Intraspecific variation. Variation in chromosome number among populations of the same taxon was found only in Eleocharis montevidensis s.l., in which, most of the studied plants are 2n = 20 and only one is 2n = 10. A similar example of polyploidy has been reported for E. geniculata 2n = 10, 20 (Sanyal and Sharma, 1972). Intraspecific variations in chromosome number involving few chromosomes have been recorded for species of Eleocharis elsewere, e.g., for E. acicularis 2n = 36-38, 50-58 (Hicks, 1929), E. acicularis f. longiseta 2n = 20, 21 (Yano et al., 2004), E. maculosa 2n = 10, 8, 7, 6 (Da Silva et al., 2008a), E. palustris 2n = 14-17, 38, 39, and E. uniglumis 2n = 46, 78-82 (Strandhede, 1965a,b, 1967), E. palustris 2n = 15-42 (Bureš et al., 2004), and E. xyridiformis 2n = 18-20 (Harms, 1968). All these examples reinforce that mechanisms of karyotype differentiation in the genus, which include dysploidy (fission and/or fusion) and polyploidy.
Given the morphological diversification in the plants identified as Eleocharis montevidensis and several differences with the "typical" E. montevidensis, the plants studied here are considered as part of a complex. Three cytotypes are recognized: E. cf. montevidensis 1 (Figures 1G, 2E), with 2n = 20 and large karyotype (54.67 μm), E. cf. montevidensis 2 (Figures 1F, 2F), with 2n = 20 and medium size karyotype (31.59 μm), and E. cf. montevidensis 3 (Figures 1F, 2G), with 2n = 10 and small karyotype (12.96 μm). Although the karyotype of E. cf. montevidensis 3 has the same diploid number as E. parishii, a closely related species, it differs in having chromosomes half sized in relation of those of E. parishii: longer chromosome 1.60 μm vs 3.34 μm, shorter 1.03 μm vs 2.15 μm, and total length of the complement 12.96 μm vs 26.99 μm (Table 2). Morphological traits of the plants allow to separating to E. cf. montevidensis 3 from E. parishii: cuspid at apex of upper sheath short and thick (vs long and thin); spikelets ovate (vs ovate-lanceolate to lanceolate); glumes ovate, almost black, broadly hyalino marginated (vs lanceolate, paler); and a thicker, obovate achene with short pyramidal stylobase (vs narrowly pyramidal to lanceolate stylobase). As currently circumscribed, E. montevidensis includes at least three elements that need further study and could represent undescribed taxa or taxa that have been included as synonyms of E. montevidensis. The complex, distributed from the United States to southern South America, needs taxonomic revision.
Intraspecific differences in size were also found in Eleocharis rostellata. All the studied samples are 2n = 60, but among those that can be confidently identified as E. rostellata two groups of chromosomes were found: small-medium, 1.90-4.30 μm with a total length of the complement of 178 μm (Figures 1J, 2J) and small to small-medium (1.503.10 μm); besides, a smaller variant with chromosomes 1.30-2.60 μm and a total length of μm 116 μm (Figures 1K, 2K) differs in some morphological external features, such as filiform culms and thinner and darker glumes, which possibly representing an incipient species which is called here E. aff. rostellata.
Karyotypic variation. The most common mechanism of karyotype variation in the species studied here was dysploidy, occurring in four of the eight species (Eleocharis densa, E. macrostachya, E. reznicekii, and E. xyridiformis). Another two taxa are diploid (E. parishii and E. cf. montevidensis 3), and three polyploid: E. acicularis and E. cf. montevidensis, which are tetraploids, and E. rostellata, a dodecaploid.
Eleocharis displays a large variation in karyotype and genome sizes (Zedek et al., 2010) and the occurrence of polyploidy and agmatoploidy/symploidy have been well documented (Da Silva et al., 2008a, b). Despite its variability, most species of Eleocharis have numbers multiple of 5 (Da Silva et al., 2008a, b), the number proposed as the basic number for the family by Löve et al. (1957). Polyploidy has been found as an important mechanism of evolution in this genus (Hoshino, 1987; Yano et al., 2004; Da Silva et al. 2008b) as in angiosperms in general (Stebbins, 1971; Soltis and Soltis, 1999). In some groups, chromosome evolution has proceeded from higher to lower numbers, as found by Hipp et al. (2007) for Carex sect. Ovales and confirmed by Mayrose et al. (2010) using probabilistic models. However, for E. dysploidy has been as important as polyploidy.
The highest ploidy level (12-ploid) and the longest length in this study were found in Eleocharis rostellata. No previous data had been published on the chromosome number nor karyotype structure for this highly variable and widely distributed species known from North America and South America. The series Rostellatae accommodates species characterized by firm and shiny sheaths, culms 10-220 cm long, flattened, wiry, sometimes arching to decumbent; spikelets ovoid-lanceolate to spindle-shaped, acute, often proliferous, and achene obtusely trigonous to plano-convex, prolongated at the apex and continuous with the conic to lanceolate stylobase (González-Elizondo and Peterson, 1997). The chromosome number and karyotype length confirm the distinctiveness of this species or species complex. The important role that karyotypes may have in the acclimation of plants has been pointed out by Mayrose et al. (2010) and Wang et al. (2011), who indicate that when plants are exposed to a large variety of abiotic stresses, their karyotypes or genomes tend to evolve to polyploidy suitable for adverse environments. The dodecaploid karyotype of E. rostellata reflects the wide adaptability of this species both to acid and to strongly alkaline habitats, being the last the most common habitat for the populations of E. rostellata studied here.
Eleocharis macrostachya and E. xyridiformis were both found to be 2n = 28. Different numbers have been registered for both species: E. macrostachya with unstable polyploid numbers ranging around 2n = 38 and E. xyridiformis with 2n = 18-20, the cytotype 19-chromosome is trisomic for one of the long chromosomes (Harms, 1968). Cytotaxonomical and morphological studies suggested that E. macrostachya may be a diploid-polyploid complex, at least partly of hybrid origin (Smith et al., 2002) with 2n = 10, 16, 18, 19, and 38.
Eleocharis macrostachya is an extremely variable taxon and E. xyridiformis has been synonymized under it or has been recognized as a species with karyotypical and morphological differences (Harms, 1968). Smith et al. (2002) noted that E. xyridiformis (treated by them as variant a of E. macrostachya) "almost certainly deserves taxonomic recognition, perhaps as a species". They differ in several morphological features and, for the plants revised by them, in chromosome numbers: 2n = 18 for variant a and 2n = 38 for variant b or typical E. macrostachya. Although under both criteria, species boundaries are diffuse among the plants treated in this study as E. macrostachya and E. xyridiformis, more information including a broader sampling and using different taxonomic approaches is needed to better understand the limits of E. macrostachya.
Eleocharis acicularis, which belongs to subgenus Scirpidium (González-Elizondo and Peterson, 1997), has a karyotype very similar to some plants of the E. montevidensis complex, with the same diploid number (2n = 20) and similar general aspect. No differences were found between subgenera in this study. Our data are in accordance to phylogenetic analyses (Roalson and Friar, 2000; Yano et al., 2004) that indicate that Scirpidium is a monophyletic group nested into a paraphyletic subgen. Eleocharis and that it could be considered a sister group to the rest of this clade (Roalson and Hinchcliff, 2007; Roalson et al., 2010). These similarities could suggest a karyotype conservation for this species. According to Guerra (2008), the chromosome number can be a plesiomorphic characteristic of a large clade or a recurrent trait which arose independently in two or more clades. Reports of E. acicularis 2n = 56 (36-38, 50-58; Hicks, 1929) suggest the highly polymorphic nature of the complex identified worldwide as E. acicularis. The plants analyzed during this study have relatively coarse rhizomes and culms (0.3-0.4 mm wide) and spikelets more than 12-flowered but other plants that also key to E. acicularis (which died in the nursery and were not analyzed during this study) have slender rhizomes, capillary culms and few-flowered spikelets. Recognition of varieties in E. acicularis is premature pending a worldwide taxonomic revision of subg. Scirpidium (Smith et al., 2002), and the plants studied in this work are considered as part of that complex.
Interspecific hybridization may be a widely overlooked evolutionary phenomenon in Eleocharis and may have a significant role in its diversification (Košnar et al., 2010). Karyological data in the present study do not support the hypothesis of the hybrid origin of E. reznicekii suggested by González-Elizondo et al. (2007). At least, no intermediate chromosome numbers were found between the putative parents, as in Bureš (1998) studies. In this study, E. reznicekii and one of its putative parents (E. densa) are 2n = 16, whereas E. macrostachya and E. xyridiformis (that also were suggested as putative parents) are both 2n = 28. To test the hybrid origin hypothesis, molecular and/or cytogenetical analyses are required (González-Elizondo et al., 2007). Because of the perennial mat-forming habit, long, horizontal rhizomes; mostly bifid styles; and biconvex, blunt angled, yellow to brown achenes almost smooth at 30x, E. reznicekii is classified into Eleocharis subg. Eleocharis, sect. Eleocharis (González-Elizondo and Peterson, 1997), which also includes the "E. palustris complex". Eleocharis palustris has been recorded with 2n = 16 (Roalson, 2008), as in E. densa and E. reznicekii.
The karyotype features examined are useful to distinguish among the taxa studied. Chromosome structure and ploidy status as well as the interspecific variation of karyotypes (cytotypes) provide indicators of the genetic similarity between populations or species (Palomino, 2000), but caution should be applied in the interpretation of the results. As Hipp et al. (2010) have noticed for Carex, different chromosome rearrangements not necessarily represent monophyletic 'races' or infraspecies, in spite that karyotype evolution is a potential player in the speciation in that genus.
Karyotype features can be considered a good tool to distinguish species in sedges and karyotypical differences have an excellent potential to be used in evolutionary studies; however, they shall be interpreted in combination with other taxonomic characters. Additional karyotype studies as well as cytogenetical analyses are needed for Eleocharis, along with morphological and field studies to better understand the taxonomy and the evolutionary relationships of this complex genus.
Acknowledgments
We are indebted to Octavio Rosales Carrillo and Claudia Silva Salas for their help in the field and the laboratory, to Martha González Elizondo, Lorena López Enriquez, Jorge Noriega Villa, and Vanessa Mancuso for assistance, and to COFAA and EDI for stimuli for research. A licence from COTEPABE is gratefully acknowledged by the first author. Two anonymous reviewers provided helpful suggestions to improve the manuscript.
Literature cited
Beaman J.H., De Jong D.C.D. and Stoutamire W.P. 1962. Chromosome studies in the alpine and subalpine floras of Mexico and Guatemala. American Journal of Botany 49:41-50. [ Links ]
Bureš P 1998. A high polyploid Eleocharis uniglumis S.L. (Cyperaceae) from central and southeastern Europe. Folia Geobotanica 33:429-439. [ Links ]
Bureš P., Rotreklová O., Stoneberg Holt S.D. and Pikner R. 2004. Cytogeographical survey of Eleocharis subser. Eleocharis in Europe 1. Eleocharis palustris. Folia Geobotanica 39:235-257. [ Links ]
Catling P.M. and Hay S.G. 1993. The hybrid origin of Eleocharis macounii. Rhodora 95:85-96. [ Links ]
Da Silva C.R.M., González-Elizondo M.S. and Vanzela A.L.L. 2005. Reduction of chromosome number in Eleocharis subarticulata (Cyperaceae) by multiple translocations. Botanical Journal of the Linnean Society 149:457-464. [ Links ]
Da Silva C.R.M., González-Elizondo M.S. and Vanzela A.L.L. 2008a. Chromosome reduction in Eleocharis maculosa (Cyperaceae). Cytogenetic and Genome Research 122:175-180. [ Links ]
Da Silva C.R.M., González-Elizondo M.S., Rego L.N.A.A., Domingues T.J.M. and Vanzela A.L.L. 2008b. Cytogenetical and cytotaxonomical analysis of some Brazilian species of Eleocharis (Cyperaceae). Australian Journal of Botany 56:82-90. [ Links ]
Da Silva C.R.M., Trevisan R., González-Elizondo M.S., Maldonado F.J. and Vanzela A.L.L. 2010. Karyotypic diversification and its contribution to the taxonomy of Eleocharis (Cyperaceae) from Brazil. Australian Journal of Botany 58:49-60. [ Links ]
Flores-Maya S., Moreno-Ramírez J.C., Romero-Rangel S. and Rojas-Zenteno C. 2010. Análisis morfológico y cariológico de Agave karwinskii Zucc. y Agave macroacantha Zucc. en el municipio de Zapotitlán Salinas, Puebla. Biocyt 3:181-196. [ Links ]
González-Elizondo M.S. and Peterson P.M. 1997. A classification of and key to the supraspecific taxa in Eleocharis (Cyperaceae). Taxon 46:433-449. [ Links ]
González-Elizondo M.S. and Tena-Flores J.A. 2000. Eleocharis (Cyperaceae) in the New World. In: Wilson K.L. and Morrison D.A. Eds. Monocots: Systematics and Evolution, pp. 637-643, CSIRO, Melbourne. [ Links ]
González-Elizondo M.S., Tena-Flores J.A., Alarcón-Herrera M.T., Flores-Tavizón E. and Barajas-Acosta N. 2005. An arsenic-tolerant new species of Eleocharis (Cyperaceae) from Chihuahua, Mexico. Brittonia 57:150-154. [ Links ]
González-Elizondo M.S., Rosen D.J., Carter R. and Peterson P.M. 2007. Eleocharis reznicekii (Cyperaceae), a new species from the Mexican High Plateau. Acta Botanica Mexicana 81:35-43. [ Links ]
Govaerts R., Simpson D.A. and Bruhl J.J. 2007. World Checklist of Cyperaceae Sedges. Royal Botanic Gardens, Kew Publishing, Kew. [ Links ]
Guerra M. 2008. Chromosome numbers in plant cytotaxonomy: concepts and implications. Cytogenetic and Genome Research 120:339-350. [ Links ]
Harms L.J. 1968. Cytotaxonomic studies in Eleocharis subser. Palustres: Central United States taxa. American Journal of Botany 55:966-974. [ Links ]
Hicks, G. C. 1929. Cytological studies in Cyperus, Eleocharis, Dulichium, and Eriophorum. Botanical Gazette (Chicago) 88: 132-150. [ Links ]
Hipp A.L., Rothrock P.E., Reznicek A.A. and Berry PE. 2007. Chromosome number changes associated with speciation in sedges: a phylogenetic study in Carex section Ovales (Cyperaceae) using AFLP data. Aliso 23:193-203. [ Links ]
Hipp A.L., Rothrock P.E. and Roalson E.H. 2009. The evolution of chromosome arrangements in Carex (Cyperaceae). The Botanical Review 75:96-109. [ Links ]
Hipp A.L., Rothrock P.E., Whitkus R. and Weber J.A. 2010. Chromosomes tell half of the story: the correlation between karyotype rearrangements and genetic diversity in sedges, a group with holocentric chromosomes. Molecular Ecology 19:3124-3138. [ Links ]
Hoshino T. 1987. Karyomorphological studies on 6 taxa of Eleocharis in Japan. Bulletin of the Okayama University of Science 22:305-312. [ Links ]
Hoshino T., Rajbhandari K. R. and Ohba H. 2000. Cytological studies of eleven species of Cyperaceae collected from Central Nepal. Cytologia 65:219-224. [ Links ]
Košnar J., Košnar J., Herbstová M., Macek P, Rejmánková E. and Stech M. 2010. Natural hybridization in tropical spikerushes of Eleocharis subgenus Limnochloa (Cyperaceae): Evidence from morphology and DNA markers. American Journal of Botany 97:1229-1240. [ Links ]
Kral R. 1971. A treatment of Abildgaardia, Bulbostylis and Fimbristylis (Cyperaceae) for North America. Sida 4:57-227. [ Links ]
Kukkonen I. 1990. On the genus Eleocharis (Cyperaceae) in the Flora Iranica area, with revised infrageneric classification and nomenclature. Annales Botanici Fennici 27:109-117. [ Links ]
Löve A., Löve D. and Raymond M. 1957. Cytotaxonomy of Carex section Capillares Canandian Journal of Botany 35:715-761. [ Links ]
Luceño M. and Guerra M. 1996. Numerical variation in species exhibiting holocentric chromosomes: a nomenclatural proposal. Caryologia 49:301-309. [ Links ]
Martínez J. and Palomino G. 1996. Karyotype analysis in three new species of Echeandia (Liliaceae) and cytotypes of E. reflexa. Cytologia 61:215-223. [ Links ]
Mayrose I., Barker M.S. and Otto S.P 2010. Probabilistic models of chromosome number evolution and the inference of polyploidy. Systematic Biology 59:132-144. [ Links ]
Mercado P., Ramamoorthy T.P. and Palomino G. 1989. Karyotypes of five mexican species of Salvia subgenus Calosphace (Lamiaceae). Cytologia 54:605-608. [ Links ]
Mercado-Ruaro P. and Delgado-Salinas A. 1998. Karyotypic studies on species of Phaseolus (Fabaceae: Phaseolinae). American Journal of Botany 85:1-9. [ Links ]
Mercado-Ruaro P. and Delgado-Salinas A. 2000. Cytogenetic studies in Phaseolus L. (Fabaceae). Genetics and Molecular Biology 23:985-987. [ Links ]
Mercado-Ruaro P. and Delgado-Salinas A. 2009. Karyotypic analysis in six species of Phaseolus L. (Fabaceae). Caryologia 62:167-170. [ Links ]
MicroMeasure version 3.3, available on the Internet at http://www.colostate.edu/Depts/Biology/MicroMeasure [ Links ]
Muasya A.M., Simpson D.A., Verboom G.A., Goetghebeur P., Naczi R.F.C., Chase M.W. and Smets E. 2009. Phylogeny of Cyperaceae based on DNA sequence data: current progress and future prospects. The Botanical Review 75:2-21. [ Links ]
Palomino G. 2000. Genome analysis of Mexican Flora. Geneticsand Molecular Biology 23:921-924. [ Links ]
Palomino G. and Heras H.M. 2001. Karyotypic studies in Opuntia cochinera, O. hyptiacantha, and O. streptacantha (Cactaceae). Caryologia 54:147-154. [ Links ]
Reznicek A.A. 1990. Evolution in sedges (Carex, Cyperaceae). Canadian Journal of Botany 68:1409-1432. [ Links ]
Roalson E.H. 2008. A synopsis of chromosome number variation in the Cyperaceae. The Botanical Review 74:209-393. [ Links ]
Roalson E.H. and Friar E.A. 2000. Infrageneric classification of Eleocharis (Cyperaceae) revisited: Evidence from the internal transcribed spacer (ITS) region of nuclear ribosomal DNA. Systematic Botany 25:323-336. [ Links ]
Roalson E.H. and Hinchliff C. 2007. Phylogenetic relationships in Eleocharis R.Br. (Cyperaceae): comparisons with classification, morphology, biogeography, and physiology. In: Barbosa L.M. and dos Santos Jr. N.A. Eds. A Botânica no Brasil: pesquisa, ensino e políticas públicas ambientais. 58° Congresso Nacional de Botânica, pp. 304-307, Sociedade Botânica do Brasil. Saõ Paulo. [ Links ]
Roalson E.H., Hinchliff C.E., Trevisan R. and da Silva C.R.M. 2010. Phylogenetic relationships in Eleocharis (Cyperaceae): C4 photosynthesis origins and patterns of diversification in the spikerushes. Systematic Botany 35:257-271. [ Links ]
Romero Z.C. 1986. A new method for estimating caryotype asymmetry. Taxon 35:526-530. [ Links ]
Sanyal, B. and Sharma, B. 1972. Cytological studies in Indian Cyperaceae I. Tribe Scirpeae. Cytologia 37:13-32. [ Links ]
Smith S.G., Bruhl J.J., González-Elizondo M.S. and Menapace F.J. 2002. Eleocharis. In: Flora of North America Editorial Committee, Ed. Cyperaceae. Flora of North America North of Mexico, Vol. 23, pp. 60-120, Oxford University Press, New York. [ Links ]
Soltis D.E. and Soltis P.S. 1999. Polyploidy: recurrent formation and genome evolution. Trends in Ecology and Evolution 14:348-352. [ Links ]
Stebbins G.L. 1938. Cytological characteristics associated with the different growth habits in the Dicotyledons. American Journalof Botany 25:189-198. [ Links ]
Stebbins G.L. 1971. Chromosomal Evolution in Higher Plants. Edward Arnold, London. [ Links ]
Strandhede S.O. 1965a. Chromosome studies in Eleocharis, subser. Palustres. I. Meiosis in some forms with 15 chromosomes. Hereditas 53:47-62. [ Links ]
Strandhede S.O. 1965b. Chromosome studies in Eleocharis, subser. Palustres. III. observations on western european taxa. Opera Botanica a Societate Botanica Lundensi 9:1-86. [ Links ]
Strandhede S.O. 1967. Eleocharis, subser. Eleocharis in North America. Taxonomical comments and chromosome numbers. Botaniska Notiser 120:355-368. [ Links ]
Svenson H.K. 1929. Monographic studies in the genus Eleocharis I. Rhodora 31:121-242. [ Links ]
Tapia-Pastrana F. 2010. Citogenética de Calochortus barbatus (Liliaceae: Calochortoideae) de la Reserva Ecológica del Pedregal de San Ángel, en México, Distrito Federal. Revista Mexicana de Biodiversidad 81:649-654. [ Links ]
Tapia-Pastrana F. and Gómez-Acevedo S.L. 2005. El cariotipo de Pithecellobium dulce (Mimosoideae-Leguminosae). Darwiniana 43:52-56. [ Links ]
Tapia-Pastrana F. and Jiménez-Salazar A. 2011. Los cariotipos de Cologania grandiflora y Erythrina americana (Leguminosae-Papilionoideae-Phaseoleae) de la Reserva Ecológica del Pedregal de San Ángel, México. Revista Mexicana de Biodiversidad 82:776-781. [ Links ]
Tapia-Pastrana F., Fernández-Nava R., Gómez-Acevedo S. y Mercado-Ruaro P. 2004. Estudio cromosómico en tres especies de Karwinskia (Rhamnaceae) endémicas de México. Anales del Instituto de Biología. Serie Botánica 75:1-10. [ Links ]
Tapia-Pastrana F., Mercado-Ruaro P. y Gómez-Acevedo S. 2012. Contribución a la citogenética de Tamarindus indica (Leguminosae: Caesalpinioideae). Acta Botanica Mexicana 98:99-110. [ Links ]
Vanzela A.L.L., Guerra M. and Luceño M. 1996. Rhynchospora tenuis Link (Cyperaceae): a species with the lowest number of holocentric chromosomes (n = 2). Cytobios 88:219-228. [ Links ]
Wang X., Zhang T., Wen Z., Xiao H., Yang Z., Chen G. and Zhao X. 2011. The chromosome number, karyotype and genome size of the desert plant diploid Reaumuria soongorica (Pall.) Maxim. Plant Cell Reports 30:955-964. [ Links ]
Yano O., Katsuyama T., Tsubota H. and Hoshino T. 2004. Molecular phylogeny of Japanese Eleocharis (Cyperaceae) based on ITS sequence data, and chromosomal evolution. Journal of Plant Research 117:409-419. [ Links ]
Zedek F., Smerda J., Smarda P. and Bureš P. 2010. Correlated evolution of LTR retrotransposons and genome size in the genus Eleocharis. BMC Plant Biology 10:265. [ Links ]

