











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Habanero pepper (Capsicum chinense Jacq.) is one of the horticultural crops of greater culinary, social and economical importance in the Yucatan Peninsula. In 2010, the Mexican Institute of Intellectual Property (IMPI) gave the appellation of origin “Chile habanero de la Península de Yucatán” (DOF, 2010), this increased its demand in fresh as well as for the industry (Zavala et al., 2013). However, the loss of viability and low germination of the seed, as well as the heterogeneity in seedlings emergence, are the main issues presented by this crop (Garruña-Hernandez et al., 2014); some of the factors to which these issues are attributed are the state of ripeness of the fruit at the moment of harvest, the position of the fruit in the plant and the inhibitory substances in the seed’s cover (Randle & Honma, 1981). Other factors, like the extraction process, the content of moisture and the seed selection, cause physiological and biochemical alterations that have an effect on the physiological quality of the seeds and seedlings (Doijode, 2001; Vidigal et al., 2009). In this sense, the ripeness of fruits from the Capsicum genus is known to affect germination of the seeds (Vidigal et al., 2006; Zavala et al., 2015). Some studies realized in Capsicum annuum demonstrated that storing the fruits before extracting the seeds allows to complete the ripeness of the embryo and to increase the germination and vigor of the seeds (Dias et al., 2006; Dos-Santos et al., 2016). Nonetheless, in the case of Capsicum chinense, the following question was laid out: Do the ripeness of the fruit and post-harvest storage time increase percentage and rate of seeds germination as well as seedlings emergence? Based on the aforementioned, the objective of this study was to identify the proper ripening stage of the fruit to obtain seeds of quality and to determine the effect of the post-harvest storage of fruits on germination and emergence.
Material and Methods
Seeds production
A habanero pepper (Capsicum chinense Jacq.) seedbed was established in Motul, Yucatan. Seeds from the variety Mayapan were used, provided by the INIFAP Mococha experimental field, they were sown in germinating trays with Canadian moss-based substrate (Sunshine, Proveedora Agricola, Guadalajara, Mexico). Seedlings were established in a 200 m2 plot at the INIFAP Mococha experimental field, to 35 days after sowing (das). The distance of sowing between plants was 0.30 m and 1.50 m between furrows. The agronomic handling was performed according to Tun (2001).
At around 45 das, flowering started, at this stage flowers in anthesis were labelled. Fruits were harvested at unripe (green), half-ripe (half green and half orange) and ripe (orange) stages, 28, 35 and 40 days post-anthesis (dpa), respectively. Seeds were immediately extracted from a portion of harvested fruits (control: fruits without storage), like local producers of seedlings traditionally do it. Another portion of harvested fruits (unripe, half-ripe, and ripe) was stored during 7 and 14 days post-harvest, at a temperature of 25 ± 2 ºC and 60 % of relative humidity (RH). After the corresponding period or storage, the seeds were extracted from the fruits with an apple corer, disinfected with chlorine at 2 % during three minutes and were rinsed three times with distilled water. Finally, the seeds were dried in the shade until reaching a moisture content within a range of 6-10 % (SNICS, 2014), the moisture content test was performed in accordance with the International Seed Testing Association (ISTA, 2004). The seeds were stored in silvered polyethylene bags and kept in refrigeration at 10 ºC until evaluation (7 days later). The treatments were V = green fruit, not-stored, V7 = green fruit, stored for 7 days, V14 = green fruit, stored for 14 days, P = half-ripe fruit, not-stored, P7 = half-ripe fruit, stored for 7 days, P14 = half-fruit, stored for 14 days, M = ripe fruit, not-stored, M7 = ripe fruit, stored for 7 days, and M14 = ripe fruit, stored for 14 days.
Evaluated variables
All the evaluations of germination, emergence and growth were performed in the laboratorio de Biotecnologia y Fisiologia Vegetal del Instituto Tecnologico de Conkal, Mexico.
In order to determine the moisture content (MC), five samples of 0.5 g of seeds from each treatment were dried in a forced-air convection furnace at 103 ºC until obtaining a constant dry weight (approximately 17 h). Initial fresh weight (FW) and final dry weight (DW) were used to calculate the moisture content of seeds according to the ISTA (2004) with the following formula:
Electric conductivity (EC) was determined by submerging during 24 hours a set of 50 seeds into 50 mL of deionized distilled water at 25 ºC, the readings were realized with a conductometer (Consort C931, Debruyne Instruments, Wichelen, Belgium). Four repetitions of 50 seeds per treatment were evaluated in accordance with Vidigal et al. (2009).
The percentage of germination (%G) was evaluated by sowing 20 seeds per Petri dish, dishes with 90 mm in diameter were used and humidified paper tissues with 4 mL of deionized distilled water as a substrate, five Petri dishes per treatment were established, each dish represented a repetition. The growth chamber where the germination was evaluated had a temperature of 22 ± 1 ºC, a relative humidity of 60 % and was in total darkness. The germination was counted during 14 days and a germinated seed was considered when the radicular protrusion was observed. The following formula was used:
Where %G = percentage of germination, n = total number of germinated seeds at the end of the evaluation and N = number of seeds sown.
Seedling emergence was evaluated in germinating trays with 200 cavities during 7 days. Twenty seeds were sown per repetition and five repetitions per treatment. Canadian moss was used as a substrate (Sunshine, Proveedora Agricola, Guadalajara, Mexico). The same formula used to calculate %G was used to calculate the percentage of emergence (%E).
So as to obtain the germination rate (GR), the germination was counted during 14 days and was calculated with the formula for seeds germination rate proposed by the ISTA (2004):
Where GR = germination rate, ni = number of germinated seeds during the interval Ti, Ti = time from the sowing until the counting day.
For the emergence rate (ER) the number of emerged seedlings was counted daily until 7 das and the same formula was used as for calculating GR.
Root length was evaluated 7 and 14 das, pictures of 10 seedlings per treatment were taken and processed with ImageJ software (National Institute of Health, Maryland, United States) to calculate root length.
With root length data, the relative growth rate of root length was calculated, according to the formula proposed by Hunt et al. (2002):
Where RGRRL = relative growth rate of root length; InL1 = natural logarithm of root length at7 das; InL2 = natural logarithm of root length at 14 das; C1 = day on which the first measurement was performed (7 das); C2 = day on which the second measurement was performed (14 das).
Experimental design and statistical analysis
The experimental design was completely randomized, with bi-factorial adjustment, one factor was the ripeness state of the fruits (unripe, half-ripe and ripe) and another factor was the period of post-harvest storage of the fruits (7 and 14 days post-harvest; control = not-stored). Data in percentage were arcsine square root transformed; an analysis of variance (ANOVA, p≤0.05) was performed on all data. A comparison of means test (Tukey’s multiple comparison test, α = 0.05) was carried out on the variables when significant differences were detected among treatments. The analyses were performed with Statistica 7 software (Statsoft, Tulsa, Ok, EE. UU. A.).
Results and Discussion
In the analysis of variance, significant differences were found in all the variables. Post-harvest storage and fruit ripeness factors had highly significant statistical differences (p≤0.01). In the interaction of factors (storage x ripeness), the electrical conductivity and germination parameters (%G and GR) were highly significant (p≤0.01); while emergence parameters (%E and ER) and radicle growth (RL7, RL14 and RGRRL) had significant differences (p≤0.05) (Table 1).
Table 1 F-value of moisture content (MC), electrical conductivity (EC), germination (%G), emergence (%E), germination rate (GR), emergence rate (ER), root length at 7 (RL7) and 14 (RL14) days after sowing and relative growth rate of RL (RGRRL) of seeds from habanero pepper fruits harvested in different stages of maturation and post-harvest storage.
| Treatments | MC | EC | %G | %E | GR | ER | RL7 | RL14 | RGRRL |
|---|---|---|---|---|---|---|---|---|---|
| APdeF | 0.22ns | 224.14** | 73.05** | 45.32** | 310.42** | 137.69** | 167.43** | 93.50** | 60.00** |
| EMdeF | 3.44ns | 226.71** | 55.79** | 32.13** | 111.21** | 85.97** | 48.70** | 17.16** | 10.90** |
| APdeF x EMdeF | 1.10ns | 64.71** | 12.29** | 12.03* | 29.54** | 33.78* | 6.21* | 4.07* | 4.98* |
| CV | 12.41 | 6.41 | 9.03 | 15.54 | 7.46 | 11.34 | 28.13 | 26.27 | 33.46 |
ns = not significant; * = significant differences (p≤0.05); ** = highly significant differences (p≤0.01); APdeF = fruits post-harvest storage; EM-deF = fruits ripening stages; CV = coefficient of variation.
Moisture content
The moisture content in seeds from all the treatments was between 6 and 10 %. According to Copeland and McDonald (2001), that is the moisture interval for live seeds, in addition it is the range established by SNICS (2014) in all the categories of seeds (basic, registered, certified and enabled).
Electrical conductivity
The electrical conductivity (EC) of the seeds is associated to the flow of solutes (amino acids, sugars) and electrolytes (mainly potassium) through cellular membranes (Vidigal et al., 2009). According to the comparison of means, the lowest EC was observed in the three ripeness stages with 14 days of storage (UR14, 0.6; HR14, 0.6; R14, 0.6 µS mL-1) and in the seeds from half-ripe and ripe fruits with 7 days of storage (HR7, 0.6; R7, 0.6 µS mL-1); the highest values of EC were in the seeds from not-stored unripe fruits (UR, 1.5 µS mL-1) (Figure 1). Low values of electrical conductivity in seeds is an indicator of stable cellular membranes, with lower flow of solutes and electrolytes (Powell, 1986), in other words, these were seeds with greatest vigor, germination and viability. I this context, seeds of not-stored unripe fruits had a higher flow of solutes and electrolites, it is probable that the seeds that were in a physiologically unripen stage. However, when storing green fruits for 14 days (UR14) seeds electrical conductivity decreased of 40 % (slow releasing of solutes and electrolytes), indicating that during fruits post-harvest storage, the seeds continued their ripeness and stabilized their cellular membranes (Dias et al., 2006).
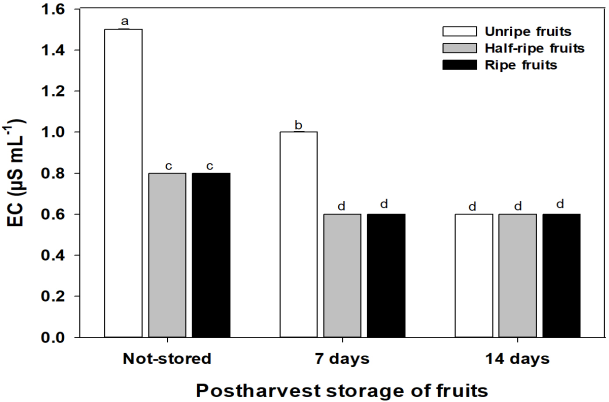
Figure 1 Electrical conductivity (EC) of habanero pepper seeds ( Capsicum chinense Jacq.) extracted from unripe, half-ripe and ripe fruits; not-stored and stored during 7 and 14 postharvest days. Data were presented as mean ± S.E.M. Different letters indicate significant differences among treatments (Tukey, α = 0.05, LSD = 0.00606). n = 200.
Germination and emergence
Fruits post-harvest storage for 14 days increased the percentage of germination of the seeds in the three fruits ripeness stages (R14, 98 %; HR14, 94 % and UR14, 91 %), seeds germination percentage R7 (91 %) was close to it; but, the germination of HR7 (73 %), HR (69 %) and UR7 (61 %) was at least 25 % inferior to the one of R14; likewise, the germination of seeds obtained from not-stored unripe fruits was the lowest (18 %) (Figure 2A). Fruit ripeness at the moment of the harvest obviously affected seeds germination (UR, 18 %; HR, 69 % and R, 83 %). In addition, fruits post-harvest storage before obtaining their seeds increased the final percentage of germination, regardless of the ripeness stage in which fruits were harvested, in other words, germination of the seeds from unripe, half-ripe and ripe fruits stored for 14 days increased 73, 25 and 15 %, respectively, in comparison with not-stored fruits. It is probable that the embryo from unripe seeds had the required structures (radicle, hypocotyl and cotyledons) for germinating, but without the physiological ripeness and the reserves required during germination (Ohto et al., 2007; Trigiano et al., 2008). Storing habanero pepper fruits (unripe, half-ripe and ripe) for 14 days increased germination with values higher than 90 %, this value is higher than the one required for habanero pepper seeds (85 %) in the categories (basic, registered, certified and enabled) of the SNICS (2014) and higher than the value that was reported by Dos-Santos et al. (2016) who obtained 85 % of seeds germination of ripe fruits of C. chinense stored for seven days. In this sense, Carrillo et al. (2009) mentioned that seeds from Capsicum genus completed their physiological ripeness by going through a period of rest that can be from one to six weeks after the fruits were harvested. Seeds viability associated with fruits post-harvest storage times was attributed to the fact that fruits from Capsicum genus were climacteric and continued their physiological ripeness in tandem with the embryo of the seed, this allowed the seed to finish its formation process and to decrease the content of abscisic acid (ABA), preparing for germination (Kermode, 1995).
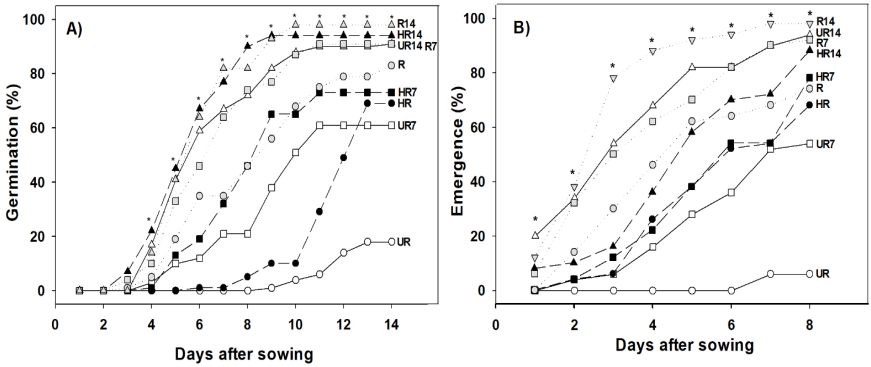
Figure 2 A) Germination and B) Emergence of habanero pepper seeds (Capsicum chinense Jacq.) extracted from unripe (UR), half-ripe (HR) and ripe (R) fruits; not-stored (control) and stored during 7 (UR7, HR7 y R7) and 14 (UR14, HR14 y R14) postharvest days. Data were presented as mean, * = significant statistical differences among treatments (ANOVA, p ≤ 0.05; LSD of %G = 0.177; LSD of %E = 0.294), n = 100.
In seedlings emergence, significant statistical differences were observed (p≤0.05) from the first day after sowing. Storing post-harvested fruits favored the emergence of seedlings, the treatments R14 (98 %), HR14 (88 %) and UR14 (94 %) exceeded the treatments without storage R (74 %), HR (68 %) and UR (6 %) for 24, 20 and 88 %, respectively; R7 (92 %) was the only treatment of those stored for seven days with seedlings emergence higher than 85 %, the rest of the treatments remained under 80 % (Figure 2B). In this sense, Doijode (2001) mentioned that during fruits post-harvest storage, the embryo continued its physiological ripeness process until completing an adequate formation in all its stages (histodifferentiation, accumulation of reserves and tolerance to desiccation), which triggered changes in endogenous concentrations of gibberellic acid and abscisic acid, this favored seedlings emergence and root length.
Germination and emergence rate
Germination and emergence rates are indicators of the speed at which seeds germinate and seedlings emerge, respectively. Just like percentages of germination (%G), germination rates (GR) of seeds from treatments R14 (19.22 germinated/day), HR14 (20.08 germinated/day), UR14 (17.59 germinate/day) and R7 (16.62 germinated/day) were statistically higher (Tukey, α = 0.05) than the rest of the treatments (Table 2), this tendency was observed since the seventh day after sowing, where only these treatments (R14, HR14, UR14 and HR7) had exceeded 50 % of germination (Figure 2A). However, in the emergence rate (ER), the number of seedlings emerged per day of R14 (13.93 seedlings/day) was statistically similar to the one of UR14 (12.67 seedlings/day), but superior to the rest of the treatments (Tukey, α = 0.05); with the exception of R7 (9.95 seedlings/day) and R (7.05 seedlings(day), treatments from fruits stored for seven days (HR7 and UR7) and not-stored fruits (HR and UR) had emergence rates under 5 seedlings/day (Table 2), which was reflected in the percentage of emergence (Figure 2B). According to Matthew & Khajeh-Hosseini (2006) a slow germination produced heterogeneous seedlings, but high emergence rates correlated with bigger and more homogeneous seedlings (Demir et al., 2008). Likewise, Bradford (2004) and Bewley et al. (2013) mentioned that unripe fruits (green) provided low quality seeds, but storing them for a period of time before the extraction of seeds can increase germination up to 73 % and seedlings emergence up to 88 %.
Table 2 Germination rate (GR) and emergence rate (ER) of habanero pepper seeds (Capsicum chinense Jacq.) extracted from unripe (UR), half-ripe (HR) and ripe (R) fruits; not-stored (control) and stored during 7 and 14 postharvest days.
| Treatments | GR (germinated/day) |
ER (seedlings/day) |
|---|---|---|
| Unripe | 0.98 ± 0.26 e | 0.16 ± 0.11 f |
| Unripe 7 days | 7.88 ± 0.75 c | 3.38 ± 0.33 e |
| Unripe 14 days | 17.59 ± 0.61 a | 12.67± 0.57 ab |
| Half-ripe | 4.00 ± 0.27 d | 4.30 ± 0.73 e |
| Half-ripe 7 days | 10.72 ± 0.67 bc | 4.56 ± 0.25 de |
| Half-ripe 14 days | 20.08 ± 0.88 a | 7.19 ± 0.88 cd |
| Ripe | 12.08 ± 1.13 b | 7.05 ± 0.84 cd |
| Ripe 7 days | 16.62 ± 0.36 a | 9.95 ± 0.43 bc |
| Ripe 14 ays | 19.22 ± 0.55 a | 13.93 ± 0.63 a |
Data were presented as mean ± S.E.M. Different letters in the same column indicated significant differences among treatments (Tukey, α = 0.05; LSD of GR = 0.051; LSD of ER = 0.057). n = 100.
Root length and relative growth rate
In root length of seedlings, significant statistical differences were observed (ANOVA, p≤0.05) among treatments at 7 das, as well as at 14 das. At 7 das, roots of seedlings from R14 (3.8 cm) were the longest ones; in contrast, roots of seedlings from UR (0 cm) did not grow (Figure 3A). At 14 das, seedlings from R14 had the longest roots (6.3 cm), but values for seedlings from treatments HR14 (5.2 cm), UR14 (5.5 cm) and R7 (5.3 cm) were very close; nevertheless, roots of seedlings from UR were the shortest ones (0.49 cm) (Figure 3A). In the relative growth rate based on root length (RGRRL), similar tendencies were observed. In the three treatments where fruits were stored for 14 days (R14, HR14 and UR14, 1.57, 1.56 and 1.63 mm cm-1 day-1, respectively) and in R7 (1.58 mm cm-1 day-1) and HR7 (1.53 mm cm-1 day-1), there were no significant statistical differences, but they were statistically superior to HR (1.24 mm cm-1 day-1), UR7 (1.33 mm cm-1 day-1) and UR (0 mm cm-1 day-1) treatments (Figure 3B).
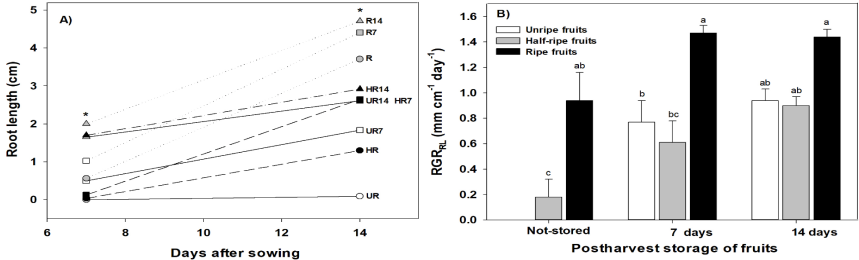
Figure 3 A) Root length (RL) and B) Relative growth rate of RL (RGRRL) of seedlings growing from habanero pepper seeds (Capsicum chinense Jacq.) extracted from unripe (UR), half-ripe (HR) and ripe (R) fruits; not-stored (control) and stored during 7 (UR7, HR7 and R7) and 14 (UR14, HR14 and R14) post-harvest days. Data were presented as mean ± S.E.M., * = significant statistical differences (ANOVA, p≤0.05). Different letters indicated significant differences among treatments (Tukey, α = 0.05; LSD of RL at 7 days = 0.418; LSD of RL at 14 days = 0.856; LSD of RGRRL = 0.366), n = 50.
In general, root length increased with the storage and with fruit ripeness, in this work the roots of seedlings growing from seeds collected from unripe and half-ripe fruits without storage were the ones presenting the lowest growth (Figure 4). In this regard, Cervantes et al. (1998) and Paponov et al. (2000) mentioned that the deficiency in the increase of root length left seedlings at a disadvantage to obtain resources to subsist.
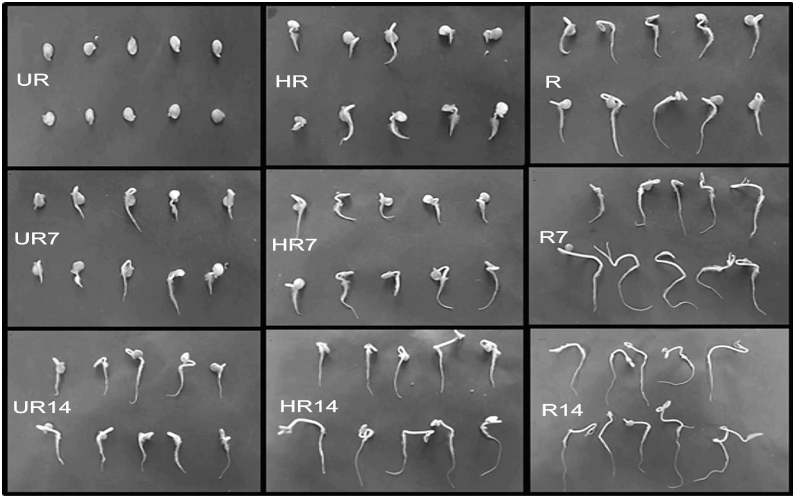
Figure 4 Habanero pepper seeds at seven days after sowing, extracted from unripe (UR), half-ripe (HR) and ripe (R) fruits; not-stored (control) and stored during 7 (UR7, HR7 and R7) and 14 (UR14, HR14 and R14) post-harvest days.
According to the ISTA (2004), seed vigor is the total sum of the properties that determine the potential level of activity and development during germination and seedlings emergence. Seeds from ripe fruits per se obviously produced more vigorous seedlings, but storing half-ripe and unripe fruits for 14 days before extracting their seeds propitiated the production of vigorous seedlings. In this sense, the embryo of habanero pepper seeds was assumed to continue its ripeness process while remaining attached to the placenta of the fruit, no matter whether the fruit is in the plant or in post-harvest storage. In the seed industry, harvesting unripe and half-ripe fruits for seeds extraction instead of waiting until the fruit reaches its total ripeness in the plant is important because it prevents fruits to be exposed to biotic and abiotic damages in the field. Likewise, advancing fruits harvest (unripe and half-ripe fruits) can increase the number of cuts in the cycle of a crop, because carbon assimilates (photosynthates) that fruits demand for their ripeness are canalized to the formation of new floral primordia (Marcelis et al., 2004; Garruña-Hernández et al., 2012).
Conclusion
Harvesting both unripe and half-ripe fruits and storing them or 14 days increased seeds physiological attributes and seedlings quality, which allowed to obtain similar values to those of the seeds obtained from ripe fruits. Fruits post-harvest storage before seeds extraction increased percentages and rates of germination and emergence, decreased electrical conductivity of the solution where seeds were soaked and increased roots growth of seedlings from the first week after sowing. Considering data generated in this research work, seeds producers were recommended to harvest unripe habanero pepper fruits (unripe and half-ripe) and to store them for 14 days, so that they finalize their ripeness before extracting the seeds; thus obtaining quality seeds and avoiding competition among fruits in the plant.

