











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Research on medium and large-sized mammals in Bolivia has so far focused on lowland habitats, especially the Amazonian, Chiquitano and Chaco forests, with a lack of information from Andean montane habitats, which harbor a great diversity of poorly known species (Wallace et al. 2010a). Currently, Andean montane habitats are undergoing an intensification of land use, as well as the effects of climate change, which are also very noticeable and accelerated in these regions (Herzog et al. 2011). Andean montane forests are unique and fragile ecosystems that are the main source of water on a regional and continental scale, but they are currently experiencing accelerated deforestation rates, which are the greatest threat to their significant biological diversity (Tejedor-Garavito 2012).
Abundance and density estimates are important for wildlife management and conservation studies because they are population parameters that vary over time and space, which allow monitoring temporal variations of the population and indirectly assess the quality of habitats (Walker et al. 2000). Estimating the density of Neotropical mammals can be costly and difficult to obtain due to nocturnal habits, low numbers of individuals and evasiveness, and therefore the use of relative abundance indices is recommended for species without individual tags. These indices are easy to estimate and are expressed as photographic events per unit of effort (Rovero and Marshall 2009; Monroy-Vilchis et al. 2011).
Knowledge of the activity patterns of a species is important for understanding the biology and ecology of the species and inform management and conservation plans (Foster et al. 2013; Ayala et al. 2020). Activity patterns can be influenced by abiotic factors such as light intensity and temperature (Albanesi et al. 2016), as well as biotic factors such as physiology, inter- and intraspecific interactions (Porfirio et al. 2016), and anthropogenic disturbances (Van Dyke et al. 1986). In particular, the activity patterns of carnivores tend to be synchronized with the time of greatest vulnerability of their main prey (Theuerkauf et al. 2003).
The use of camera traps in the field of biology and particularly in wildlife research has increased exponentially in the last 25 years contributing to the knowledge of many species that are difficult to detect and study due to their low densities and elusive behavior (Karanth et al. 2004; Nichols et al. 2011). Camera traps are used as a tool for research on topics as diverse as the presence and distribution of species (Bowkett et al. 2007), species richness (Tobler et al. 2008; Ahumada et al. 2011), abundances and population densities (Wallace et al. 2003; Tobler et al. 2013), predator-prey relationships (Ayala et al. 2020), survival (Karanth et al. 2002), occupancy (Nichols and Karanth 2002), habitat preference (Alempijevic et al. 2021), activity patterns (Ayala et al. 2020), and behavior (Viscarra et al. 2019). The use of camera traps is one of the most important and versatile methods for conservation biology research (Ayala et al. 2020; Mena et al. 2020).
Our study focused on three questions: 1) What are the species of medium and large mammals present in the High Andes and montane cloud forests of the Andean portion of the Greater Madidi-Tambopata Landscape? 2) What species are the most abundant? 3) What is the pattern of activity? The information obtained will respond to information gaps in a little studied mammal community and this baseline will allow us to monitor wildlife population behavioral changes in the years to come.
Materials and methods
Study area. The research was carried out at 12 Andean sites in the Greater Madidi-Tambopata Landscape, a transboundary landscape, between northwestern Bolivia and southeastern Perú (Figure 1). This area covers approximately 142,530 km2 (Bolivia 106,810 km2; Perú 35,720 km2), composed of six national protected areas, as well several indigenous communities and municipal protected areas. This landscape is characterized by an impressive altitudinal range (180 to 6,044 masl), and a varied topography and climate that have resulted in diverse plant and animal communities and a high number of endemic species. The Puina, Pampas de Apolo, Mamacona, Cargadero, Isañuj, Sarayoj, Keara and Suchez study sites are within the Madidi National Park and Natural Area of Integrated Management (PN ANMI Madidi), Pusupunku and Piwara are within the Apolobamba National Natural Area of Integrated Management (PN ANMI Apolobamba), and the Acero Marka and Camino Yungas sites are immediately adjacent to the Cototapata National Park and Natural Area of Integrated Management (PN ANMI Cotapata). Sampling sites covered Andean and montane habitats (Table 1). The High Andes region includes mountains that exceed 6,000 masl, but our study was concentrated in an altitudinal range of 4,100 to 4,902 masl characterized by undulating plateaus crossed by mountain ranges. The climate is generally cold and dry with intense solar radiation and large daily thermal amplitudes. Average annual temperatures range from below 0 °C to 9 °C (Navarro 2002). The montane cloud forests are found on the steep slopes and peaks and can be divided into the following levels: tree line forest (3,000 to 3,600 masl), upper montane forest (2,700 to 3,000 masl), mid-montane forest (1,700 to 2,600 m asl) and lower montane forest (1,100 to 1,700 masl). The climate and topography result in a relatively low and evergreen forest with leathery leaves (Ribera 1995; Identidad Madidi and SERNAP 2017, 2019). Epiphytes abound, especially mosses and lichens, which cover tree trunks, branches, and treetops, as well as a thickness soft layer that covers the ground. The average temperature is 18 °C, and the humidity is 80 % (Paniagua-Zambrana et al. 2003).
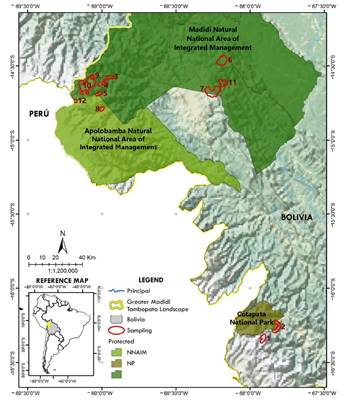
Figure 1 Map of 12 sampling sites (numbered red polygons: 1 = Acero Marka, 2 = Camino Yungas, 3 = Cargadero, 4 = Isañuj, 5 = Keara, 6 = Mamacona, 7 = Pampas de Apolo, 8 = Pasto Grande, 9 = Puina, 10 = Puina 2, 11 = Sarayoj, 12 = Suchez) in the Andean portion of the Greater Madidi-Tambopata Landscape.
Camera Trap Methodology. Between 2012 to 2017, a total of 12 camera trap campaigns were implemented. A variable number of camera trap stations were placed 1 to 2 km apart from each other, each station consisted of one or two cameras placed face to face, each set at 50 to 70 cm from the ground (Noss et al. 2013). Three digital camera trap models were used: Reconyx® HC500, HC550® (USA) and Bushnell® HD 119477 (USA). All cameras were programmed to operate 24 hours a day, to take 1 to 10 shots, depending on the camera model (Ayala et al. 2020).
The camera trap stations were placed in the predominant habitats of each study area: a) open areas along the main and secondary rivers; b) forest, often located along animal trails and small streams within the forest; c) pampa, open grassland locations, and d) rocky outcrops, this terrain is arid, sparsely vegetated, and steep. Between 7 and 71 stations were placed at each study site, sampling effective areas of between 2.1 to 92.4 km2 (Table 1). Each camera trap campaign was conducted for a period of between 5 to 80 of effective sampling days. The location of all the camera trap stations were georeferenced.
Relative Abundance Index. To quantify photographic records, we calculated independent events for each species, where photographs of the same species at a given station separated by 30 minutes are considered as separate events (O'Brien et al. 2003). Relative abundance index (RAI) was calculated through the capture rate expressed as the number of independent events per total number of traps night during the study and multiplied by 100 (O'Brien et al. 2003). The traps night are the days that the camera traps were active during the study.
Table 1 Sampling sites, habitats, number of camera trap stations and camera trap nights sampled in the Andean portion of the Greater Madidi-Tambopata Landscape.
| Year | Survey Site | Habitat | Altitude Range (masl) | Camera Trap Stations | Camera Trap Nights | Sampled Polygon (km2) |
|---|---|---|---|---|---|---|
| 2012 | Acero Marka (AMK) | High Andes - Tree Line Forest | 3,117 - 3,759 | 19 | 1,254.2 | 5.5 |
| Puina 1 (PNA1) | Tree Line Forest | 3,157 - 3,404 | 7 | 241.1 | 2.2 | |
| Pusupunku-Piwara (PP) | Tree Line Forest | 2,837 - 3,514 | 12 | 373.3 | 2.1 | |
| 2015 | Pampas Apolo (PDA) | Mid-montane Cloud Forest | 1,691 - 2,336 | 59 | 482.3 | 38.4 |
| Puina 2 (PNA 2) | High Andes - Tree Line Forest | 3,152 - 4,761 | 71 | 974.5 | 92.4 | |
| 2016 | Mamacona (MAN) | Low-montane Cloud Forest | 1,521 - 2,123 | 28 | 312.3 | 21.9 |
| Cargadero (CGR) | Mid-montane Cloud Forest | 1,317 - 3,223 | 28 | 350.3 | 3.5 | |
| Isañuj (ISÑ) | High Andes - Tree line Forest | 3,298 - 3,611 | 15 | 298.3 | 2.5 | |
| Camino Yungas (CYUN) | Mid-montane Cloud Forest | 1,670 - 3,049 | 28 | 515.4 | 17.9 | |
| 2017 | Sarayoj (SRJ) | Upper Foothill Forest | 1,157 - 1,448 | 20 | 135.9 | 8.8 |
| Keara (KR) | Upper Montane Cloud Forest | 2,750 - 3,000 | 16 | 176.4 | 2.1 | |
| Suchez (SCH) | High Andes | 4,805 - 4,902 | 8 | 30.2 | 2.4 | |
| Total | 12 | 1,157 - 4,902 | 311 | 5,144.3 | 193.5 |
Activity Pattern Analysis. All photographs were classified according to the time automatically registered on each photograph. The following categories were defined for the analysis: a) Sunrise, 30 minutes before the first ray of sunlight rises above the horizon, b) Day, begins at the moment when the first rays of the sun appear on the horizon, c) Sunset, begins at the moment when the sun is lost on the horizon, d) Night, begins 30 minutes after the sunset on the horizon (Ayala et al. 2020). This classification was made based on the sunrise and sunset time of each sampling day for the location of each sampling site (www.tutiempo.net).
Statistical analyses were performed in R software version 3.3.2 (R Core Team 2011). Using the Circular package (Agostinelli and Lund 2013), a Rayleigh test was conducted to assess the distribution uniformity on the daily activity data to evaluate whether or not species exhibited a random activity pattern over the circadian cycle. The synchronization of non-random data could indicate that the animals are nocturnal, diurnal, or crepuscular, whereas if the data are evenly distributed throughout the day, the species are classified as cathemeral (Pratas-Santiago et al. 2016).
We used Kernel density estimation to generate the activity patterns for each species, Kernel density estimations are a non-parametric way to estimate the probability density function of a distribution of records (Linkie and Ridout 2011). The analysis of activity patterns of species with more than 10 independent events was carried out (Gerber et al. 2012; Monterroso et al. 2014).
Results
The sites sampled covered an altitudinal gradient from 1,157 to 4,902 masl. A total of 311 digital camera trap stations were installed, obtaining a sampling effort of 5,144 trap nights (TN; Table 1). A total of 21,346 photographs and 1,152 independent events were obtained, identifying 28 species of mammals. The species with the highest number of photographs were Didelphis pernigra (n = 10,305), Lagidium viscacia (n = 2,030) and Lycalopex culpaeus (n = 1,929; Table 2).
Table 2 Number of photographs, independent events (IE) and Relative Abundance Index (RAI) registered at 12 sample sites (AMK = Acero Marka, PNA 1 = Puina 1, PNA 2 = Puina 2, PP = Pusupunku-Piwara, PDA = Pampas de Apolo, MAM = Mamacona, CGR = Cargadero, ISÑ = Isañuj, CYUN = Camino Yungas, SRJ = Sarayoj, KR = Keara, SCH = Suchez) in the Andean portion of the Greater Madidi-Tambopata Landscape.
| Species | Photos | IE | AMK | PNA1 | PP | PDA | PNA2 | MAM | CGR | ISÑ | CYUN | SRJ | KR | SCH |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Primates | ||||||||||||||
| Sapajus apella | 25 | 5 | 0.86±0.40 | 0.35±0.33 | ||||||||||
| Lycalopex culpaeus | 1,929 | 235 | 16.38±7.86 | 1±0.43 | 1.04±0.11 | 6.81±3.16 | 19.89±12.16 | |||||||
| Leopardus colocola | 27 | 3 | 0.3±0.17 | |||||||||||
| Leopardus jacobita | 17 | 3 | 0.31±0.18 | |||||||||||
| Leopardus pardalis | 44 | 4 | 0.36±0.34 | 2.14±1.55 | ||||||||||
| Leopardus tigrinus | 177 | 22 | 0.41±0.12 | 0.54±0.11 | 0.31±0.17 | 1.72±0.66 | 0.3±0.29 | 0.63±0.44 | 1.51±0.59 | |||||
| Puma concolor | 63 | 11 | 0.08±0.08 | 0.35±0.33 | 1.96±1.11 | 0.39±0.36 | 0.57±0.57 | |||||||
| Puma yagouaroundi | 19 | 2 | 0.39±0.39 | |||||||||||
| Conepatus chinga | 608 | 53 | 3.15±2.35 | 1.11±1.06 | 1.73±0.87 | 3.5±3.23 | ||||||||
| Eira barbara | 143 | 27 | 0.83±0.15 | 1.07±0.53 | 0.69±0.45 | 0.57±0.38 | 0.2±0.21 | 10.48±7.39 | 0.59±0.56 | |||||
| Neogale frenata | 76 | 15 | 0.15±0.15 | 0.21±0.22 | 0.11±0.11 | 0.19±0.19 | 0.57±0.57 | |||||||
| Nasua sp. | 1,086 | 33 | 0.17±0.16 | 2.9±0.42 | 3.21±1.22 | 1.01±0.53 | 0.96±0.47 | |||||||
| Tremarctos ornatus | 459 | 17 | 0.24±0.12 | 1.8±0.31 | 0.29±0.28 | 1±0.49 | ||||||||
| Artiodactyla | ||||||||||||||
| Hippocamelus antisensis | 172 | 12 | 0.48±0.28 | 0.62±0.28 | ||||||||||
| Odocoileus virginianus | 8 | 1 | 0.1±0.11 | |||||||||||
| Mazama chunyi | 1,063 | 84 | 2.47±1.02 | 3.32±0.27 | 1.33±0.21 | 1.7±0.68 | 0.31±0.23 | 1.05±0.56 | 1.44±0.72 | 3.14±1.22 | 1.77±1.72 | |||
| Mazama americana | 81 | 10 | 1.09±0.55 | 1.04±0.54 | 1.42±0.95 | |||||||||
| Xenarthra | ||||||||||||||
| Dasypus novemcinctus | 92 | 12 | 0.03±0.91 | 1.19±1.15 | ||||||||||
| Rodentia | ||||||||||||||
| Cuniculus paca | 442 | 47 | 7.67±2.26 | 4.11±3.14 | 0.6±0.58 | 5.05±2.14 | ||||||||
| Cuniculus taczanowskii | 1,961 | 76 | 1.98±0.96 | 3.32±1.22 | 1.86±0.21 | 0.43±0.45 | 2.25±0.91 | 3.08±1.26 | 2±0.98 | 0.6±0.57 | ||||
| Dasyprocta variegata | 299 | 29 | 0.21±0.21 | 4.47±1.37 | 0.54±0.56 | 1.13±0.95 | 4.97±2.86 | |||||||
| Coendou bicolor | 10 | 1 | 0.22±0.21 | |||||||||||
| Lagidium viscacia | 2,030 | 170 | 1.66±1.12 | 14.43±3.67 | 6.74±4.13 | |||||||||
| Notosciurus pucheranii | 36 | 8 | 0.63±0.47 | 1.02±0.53 | 0.19±0.19 | 0.76±0.69 | ||||||||
| Hadrosciurus spadiceus | 3 | 1 | 0.73±0.71 | |||||||||||
| Didelphimorphia | ||||||||||||||
| Didelphis marsupialis | 152 | 17 | 1.36±0.58 | 6.74±3.15 | ||||||||||
| Didelphis pernigra | 10,305 | 249 | 6.16±3.27 | 10.71±2.32 | 0.1±0.12 | 0.78±0.38 | 2.77±1.44 | |||||||
| Lagomorpha | ||||||||||||||
| Lepus europaeus | 19 | 5 | 0.52±0.36 |
The species with the highest relative abundance were L. culpaeus with a range of 1 - 19.89/100 TN, D. pernigra with 0.1 - 10.71/100 TN, L. viscacia with 1.6 - 14.43/100 TN, Eira barbara with 0.2 - 10.48/100 TN, Mazama chunyi with 0.31 - 3.14/100 TN and Nasua sp. 0.17 - 2.9/100 TN (Table 2).
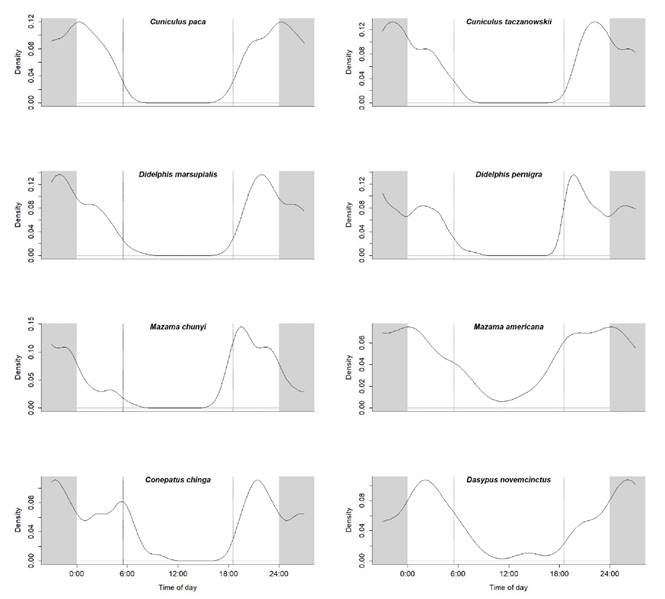
Figure 2 Activity patterns of nocturnal species in the Andean portion of the Greater Madidi-Tambopata Landscape. The two parallel dotted lines represent approximate sunrise and sunset (between 6:00 to 6:30 and 18:00 to 18:30 h, respectively, throughout the year). Shaded areas represent nighttime.
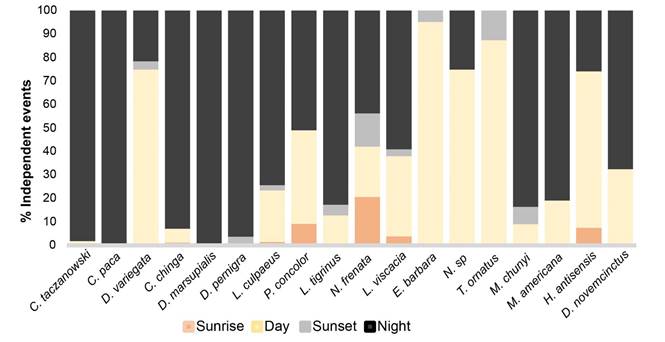
The Rayleigh test for data uniformity revealed that eleven of the 18 species with more than 10 independent events showed clear nocturnal activity patterns (Table 3). Conepatus chinga (92 % nocturnal) registered activity peaks between 20:30 to 21:30 hrs and 4:00 to 5:30 hrs. Cuniculus paca (100 %) had activity peaks between 22:30 to 24:00 hrs, whilst C. taczanowskii (97 %) had activity peaks between 20:30 to 22:00 hrs. Activity peaks were between 18:30 to 19:30 hrs for Didelphis pernigra (95 %), and between 20:30 to 22:30 hrs for D. marsupialis (100 %). The dwarf brocket deer (Mazama chunyi) is predominantly nocturnal (83 %), with very little diurnal (10 %) and crepuscular activity (7 %), and a clear activity peak between 17:30 to 19:30 hrs. The red brocket deer (Mazama americana) is predominantly nocturnal (80 %), with very little diurnal (20 %), activity peaks were between 18:30 to 24:00 hrs. The nine-banded armadillo (Dasypus novemcinctus) is primarily nocturnal (70 %), with little diurnal (30 %), and a clear activity peak between 18:30 to 1:30 hrs (Figures 2, 4). The Andean fox (Lycalopex culpaeus) is mainly nocturnal (74 %), with some diurnal (22 %) and crepuscular (4 %) activity, and peaks between 18:30 to 19:30 hrs and 22:30 to 23:30 hrs. The oncilla (Leopardus tigrinus) is primarily nocturnal (82 %), with some diurnal (14 %) and crepuscular (4 %) activity, and activity peaks between 17:30 to 21:30 hrs and 5:30 to 6:30 hrs. The northern vizcacha (L. viscacia) is nocturnal (58 %), with less diurnal (34 %) and crepuscular activity (8 %), and activity peaks between 20:30 - 01:30 hrs (Figures 3, 4).
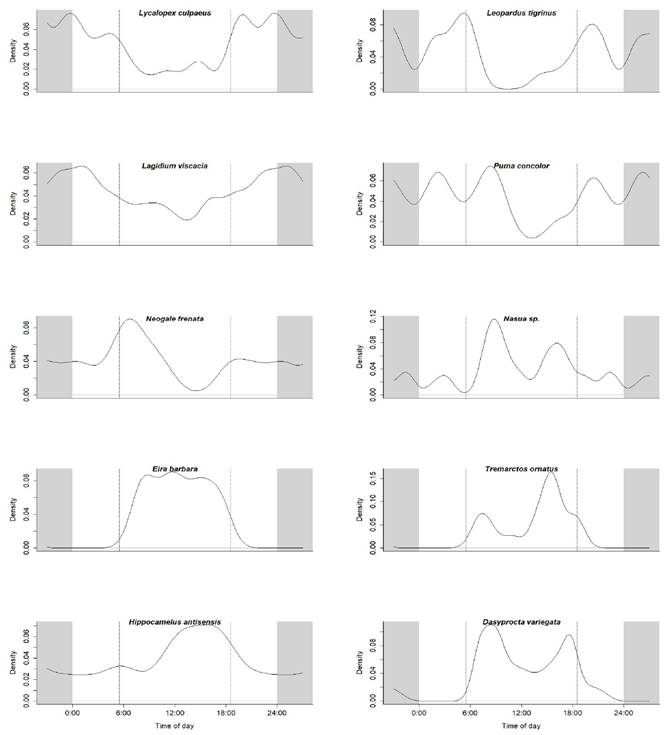
Figure 3 Activity patterns of species with nocturnal, cathemeral and diurnal habits in the Andean portion of the Greater Madidi-Tambopata Landscape. The two parallel dotted lines represent approximate sunrise and sunset (between 6:00 to 6:30 and 18:00 to 18:30 h, respectively, throughout the year). Shaded areas represent nighttime.
The Rayleigh test for data uniformity revealed that the long-tailed weasel (Neogale frenata) is cathemeral predominantly nocturnal (43 %) and crepuscular (35 %), with less diurnal activity (21 %). The puma (Puma concolor) is cathemeral predominantly nocturnal (50 %), with less diurnal (40 %) and crepuscular activity (10 %) and activity peaks between 8:30 to 11:30 hrs and 18:00 to 21:30 hrs (Figures 3, 4, Table 3). Five species presented predominantly diurnal habits (Table 3), the tayra (Eira barbara 96 %) without showing activity peaks during the day. The Andean bear (Tremarctos ornatus 88 %) showed an activity peak between 13:30 to 15:30 hrs. The agouti (Dasyprocta variegata) is mainly diurnal (76 %) with some nocturnal and crepuscular activity (24 %) and a peak between 16:30 to 17:30 hrs. The coati (Nasua sp.) was predominantly diurnal (76 %), with less nocturnal activity (24 %) and an activity peak between 06:30 to 10:00 hrs. The north Andean deer (Hippocamelus antisensis) is predominantly diurnal (70 %) with some nocturnal and crepuscular activity (30 %) and a peak between 8:30 to 17:30 hrs (Figures 3, 4).
Discussion
Camera traps can record the presence of rare species, document geographic range, and reveal behaviors, as well as estimate populations, and identify habitat requirements (Karanth and Nichols 2002; O’Connel et al. 2011). In our study we were able to register cryptic species such as N. frenata and L. tigrinus, which previously had few records in these habitats. The camera traps also confirmed the presence of a phenotypically distinct coati (Nasua sp.).
Leopardus tigrinus is one of the species that has very little abundance data in montane forest and we obtained abundance data for this species from six sampled sites, where the sites with the highest abundance were Mamacona (MAM; 1.72 ± 0.66/100 TN) and Camino Yungas (CYUN; 1.51 ± 0.59/100 TN), both sites belong to the lower and mid montane forest between 1,300 to 2,700 masl where a variety of marsupials and small arboreal rodents, which are frequent preys for this feline, are present (Wang 2002). The nocturnal activity patterns found on L. tigrinus are similar to those reported in other studies in similar habitats (Bonilla-Sánchez et al. 2020).
Table 3 Rayleigh uniformity test (Z) for the temporal activity of medium and large-sized mammals in Andean montane habitats of the Greater Madidi-Tambopata Landscape. * = for a non-random distribution (P < 0.05).
| Species | Classification | Z | p |
|---|---|---|---|
| Leopardus tigrinus | Nocturnal | 0.3709 | 0.046 |
| Lycalopex culpaeus | Nocturnal | 0.3533 | 0.01 |
| Conepatus chinga | Nocturnal | 0.5655 | 0.01 |
| Mazama chunyi | Nocturnal | 0.7300 | 0.01 |
| Mazama americana | Nocturnal | 0.4692 | 0.01 |
| Dasypus novemcinctus | Nocturnal | 0.6066 | 0.01 |
| Cuniculus paca | Nocturnal | 0.7565 | 0.01 |
| Cuniculus taczanowskii | Nocturnal | 0.7625 | 0.01 |
| Didelphis marsupialis | Nocturnal | 0.7571 | 0.01 |
| Didelphis pernigra | Nocturnal | 0.6527 | 0.01 |
| Lagidium viscacia | Nocturnal | 0.2292 | 0.001 |
| Nasua sp. | Diurnal | 0.2948 | 0.055 |
| Hippocamelus antisensis | Diurnal | 0.3262 | 0.028 |
| Eira barbara | Diurnal | 0.6608 | 0.01 |
| Dasyprocta variegata | Diurnal | 0.4517 | 0.01 |
| Tremarctos ornatus | Diurnal | 0.5809 | 0.002 |
| Puma concolor | Cathemeral | 0.2614 | *0.391 |
| Neogale frenata | Cathemeral | 0.3087 | *0.243 |
The mountain paca (C. taczanowskii) is a rodent that inhabits Andean cloud forests from Venezuela to northern Bolivia and is most abundant between 2,000 to 3,350 masl (Wallace et al. 2010b). In this study, we registered the species between 1,943 to 3,552 masl at eight of our study sites (Table 2), with lower relative abundances than those reported in the montane forests of Peru (Jiménez et al. 2010: 17.0 and 1.6/100 TN). Activity patterns were predominantly nocturnal and are consistent with those reported in other studies (Jiménez et al. 2010). C. taczanowskii is considered an upland species while Cuniculus paca a lowland species, however, they may be sympatric in certain areas of their distribution ranges (Wallace et al. 2010b). In our study, both species were recorded at the Camino Yungas (CYUN) and Cargadero (CGR; Table 2), and these sites belong to the middle montane forest, which are transitional areas between upland and lowland habitats. Due to the scarce records of C. paca, it was not possible to carry out the analyses of the overlapping of activity patterns to observe whether there was a temporal segregation between the two species as they have similar morphology and trophic requirements. It is therefore important to continue to generate information in transitional habitats to better understand the ecology of both species.
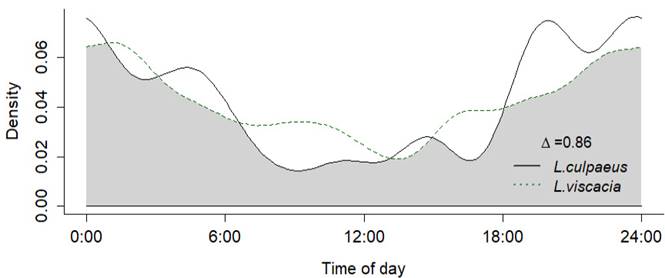
Figure 5 Overlap coefficient of daily activity patterns between Lycalopex culpaeus - Lagidium viscacia. The overlap is represented by the shaded area.
The northern vizcacha (L. viscacia) has been described as a diurnal-crepuscular species (Walker et al. 2007), but at high altitude sites nocturnal activity patterns with peaks at dawn have been reported (Lucherini et al. 2009; Huaranca et al. 2019). Our results coincide with a primarily nocturnal activity pattern, with an activity peak between 20:30 to 1:30 hrs. The sites of AceroMarka (AMK), Suchez (SCH) and Puina (PNA) where L. viscacia was recorded are above 3,700 masl where one of the main livelihoods of the local people is the raising of camelid cattle, Vicugna pacos and Lama glama (INE 2013). In these sites there is a trophic overlap regarding grass species (Festuca sp., Stipa sp.) between camelid livestock and vizcachas (Cortes et al. 2002). On the other hand, Tarifa et al. (2004) reported that vizcachas compete for food with cattle, therefore we consider that vizcachas are more active at night to avoid competing with camelids for food resources during the day. Dietary studies on the Andean cat (Leopardus jacobita) registered L. viscacia as the principal prey (Walker et al. 2007; Napolitano et al. 2008), and Andean cats exhibited an activity pattern similar to that of vizcachas (Lucherini et al. 2009; Huaranca et al. 2019). Our study yielded insufficient records of Andean cat to evaluate activity pattern overlap with vizcachas.
In our study, we obtained abundance estimates of L. culpaeus for five of the sites sampled, with the highest abundances found at the Aceromarka (AMK), Keara (KR) and Suchez (SCH) sites (Table 2). These three sites have camelid livestock, and it is possible that the high abundances of the Andean fox are related to the presence of domestic camelids, as dietary studies on L. culpaeus report their consumption of domestic camelids (Walker et al. 2007). The activity patterns of L. culpaeus are better studied in the region (Johnson and Franklin 1994; Salvatori et al. 1999; Lucherini et al. 2009), and in general agree with the findings in our study that L. culpaeus is primarily nocturnal. Dietary studies consider L. culpeus as a generalist and opportunistic species with L. viscacia as the third most important prey (Walker et al. 2007). Using data from our study we performed an overlap analysis of activity patterns between L culpaeus and L. viscacia according to standardized methodology (Ridout and Linkie 2009; Ayala et al. 2020), and we registered a significant overlap between both species (Δ1 = 0.86, IC = 0.7 - 0.99, U 2 = 0.14, P > 0.1, Watson test of homogeneity) confirming that L. viscacia is one of the main preys for L. culpaeus (Figure 5).
The puma (P. concolor) despite being the largest felid in montane and high Andean habitats, has little information on abundances in these habitats. In our study it was recorded in five sites, with low abundances ranging from 0.08 to 1.96/00 TN. It is likely that this low abundance is due to conflict between local people and predators that consume domestic camelids. Studies on puma diet in the Bolivian highlands indicate that V. pacos and L. glama are the second and third most consumed food items, representing 32 % of their diet, causing pumas to be hunted or driven away (Pacheco et al. 2004). The cathemeral activity patterns found on P. concolor are similar to those reported in other studies in similar habitats (Caceres-Martinez et al. 2016).
The tayra (E. barbara), was recorded at seven sampling sites and the site with the highest abundance was Sarayoj (SRJ; Table 2). It is likely that the abundance was higher there because it was the lowest altitude site sampled and is the transition with piedmont habitats where similar abundances of 9.49/100 TN were reported with camera traps (Wallace et al. 2010a). Compared to previous studies in montane forests (Jiménez et al. 2010), relative abundance at our study sites were higher (Table 2) than in Peru (0.8 to 2/100 TN). This is a diurnal and crepuscular species with activity peaks between 13:00 to 15:00 hrs. The activity patterns we found are consistent with those reported in other studies (Delgado et al. 2011; Albanesi et al. 2016).
The Andean bear (T. ornatus) is considered a landscape species, principally due to its ecological role as a seed disperser (Velez-Liendo and Paisley 2010) and is also primarily diurnal (Paisley and Garshelis 2006; Jiménez et al. 2010; Bonilla-Sánchez et al. 2020). Andean bears were registered at four sites in our study, at higher relative abundances than in Peru (Jiménez et al. 2010).
The unidentified coati (Nasua sp.) was registered at five sites between 1,557 to 3,398 m a.s.l., and was primarily diurnal with a relative abundance of 0.17 to 3.2/100 TN. Conversely, the mountain coati (Nasuella olivacea) in the montane forests of Colombia was primarily nocturnal with activity peaks between 19:00 to 20:00 hrs, (Ramírez-Mejía and Sánchez 2016). Previous studies indicate that coatis are mainly diurnal and crepuscular (Cáceres-Martínez et al. 2016).
Our results clearly demonstrate that D. pernigra is primarily a nocturnal species and these results coincide with previous studies (Cáceres-Martínez et al. 2016; Ramírez-Mejía and Sánchez 2016; Albanesi et al. 2016). However, Pérez-Hernández et al. (2016) reported activity patterns in low proportions during daylight hours, indicating that this behavior is possibly due to the behavioral plasticity and generalist character of this species, which is considered tolerant to anthropic intervention and urban environments.
Molina's hog-nosed skunk (C. chinga) is a nocturnal species, with no data on abundance in Bolivia (Tarifa et al. 2010). Studies in Patagonia, Argentina, revealed a completely nocturnal activity for C. chinga, with activity peaks between 19:00 to 22:00 hrs and 02:00 to 05:00 hrs (Donadio et al. 2001), although, there are recent reports of cathemeral habits in the Velez Peninsula of Patagonia (D’Agostino and Udrizar 2021). In our study, C. chinga is strongly nocturnal, with very few records during the day (Figure 2).
We provide some of the first information on the relative abundance and activity patterns of mammals in Andean habitats, especially the mysterious and relatively inaccessible montane cloud forests of the Tropical Andes. Our findings reveal the presence of 28 species of mesomammals in the sampled habitats, of which 11 species are nocturnal, 5 diurnals and 2 cathemeral species, where the most abundant species were L. culpaeus in the Andean habitats and E. barbara in montane habitats. The increasing threats to both habitats underscore the importance of long-term conservation research efforts to provide conservation management relevant knowledge on population and ecological dynamics, in addition to generating more support for national, municipal, and private protected areas in the Tropical Andes.

