











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Climate changes have occurred constantly throughout Earth’s history (Zalasiewicz and Williams 2021). However, it has been recognized that anthropic climate has been the main driver of species extinctions over the past 100 years (Cahill et al. 2013) because ecosystems are being severely transformed and threatened, with impacts reaching the global scale. Deforestation, changes in land use, and extraction of certain species are among the factors causing the greatest impact on biodiversity loss (Ochoa-Ochoa and Ríos-Muñoz 2019). Anthropic climate change is likely to compound and exacerbate these factors.
The main effects of climate change are rising ambient temperature and changes in humidity and dryness (IPCC 2021). The global rise in temperature has unavoidable effects at global, regional, and local scales that occur at a faster rate than the capacity of species for physiological adaptation, thus affecting variations in population dynamics, behavior, and ecological interactions, leading to changes in their geographic distributions (Elith and Leathwick 20091; Ackerly et al. 2010; Aguado and Escalante 2015). Protecting the remaining primary forests, especially those with high carbon capture, has been suggested as a natural approach to controlling climate change and preventing biodiversity loss (Griscom et al. 2017).
Local stressors, mainly derived from the ever-growing human population, have led to changes in forest cover and land use. As a result, primary or secondary vegetation has been converted into extensive crop fields, pastures dedicated to intensive livestock raising, and human settlements (Mas and Flamenco 2011). Urban areas are also undergoing expansion, most often with no planning or structure. This phenomenon is causing damage to biodiversity, hydrological systems, and ecosystems due to the growing demand for services, construction of roads and airports, urban development, and water extraction for human consumption, among others (Grimm et al. 2008).
The term “protect” (to cover or shield from injury or destruction; to defend, guard; Merriam-Webster 1961) is frequently mentioned and has been adopted in the conservation field. In 2015, a group of world leaders agreed on an agenda for sustainable development, with the protection of the Earth included as one of the goals (CDB 2018). Aiming to meet the objective of “protection” against the different anthropic pressures, México has created a system of Protected Natural Areas (PNAs) covering 11.14 % of the country (Conanp 2019). This system harbors different ecosystems with their respective assemblages of flora and fauna. In many cases, PNAs were created to protect species endemic to the region or the country that are in some risk category (Bezaury-Creel and Gutiérrez-Carbonell 2009).
Worldwide, the vast majority of PNAs are affected by anthropic pressures at three levels (global, regional, and local), so that assessing the effects of climate change on particular species is a complex task. General and specific pressures act in synergy where the individual effect of each of the different stressors cannot be clearly identified. In Baja California Sur, Sierra La Laguna has an unique condition worldwide for being part of a sky island - an isolated area hosting different types of ecosystems along an elevational gradient (Osborne et al. 2019). It faces little anthropic disturbance: it has no access roads to the middle and upper parts, it is located in an area with very low human population density, there are no productive activities associated with the mountain range, and any human activities carried out in the area are minimal and low-impact. The oak-pine forest located at the highest elevations can only be reached on foot and with controlled access. The mountain range is home to several endemic taxa, such as the reptiles Elgaria paucicarinata and Xantusia vigilis; the birds Hylocharis xantusii, Melanerpes formicivorus, Pipilo maculatus magnirostris, Sitta carolinensis, and Vireo victoriae; and the mammals Peromyscus truei lagunae and Sorex ornatus lagunae (Álvarez-Cárdenas et al. 1988; Rodríguez-Estrella 1988; Cortés-Calva 2012; Segura-Trujillo et al. 2012). Despite these favorable features, global climate change may affect some taxa that are endemic to this PNA. The objective of this study is to assess the potential impact of climate change on taxa endemic to this sky island exposed to virtually no local anthropic pressures.
Materials and Methods
Study area. The Sierra La Laguna Biosphere Reserve is located in the southern region of the Baja California peninsula, in the state of Baja California Sur, between the municipalities of La Paz and Los Cabos. This reserve is unique in its type for being part of a sky island due to its natural isolation, with a maximum elevation of 2,080 meters (Figure 1). The low human population density in the region presents virtually no anthropic pressure, making this region an ideal model of ecosystem protection. This sky island harbors various vegetation types, including arid ecosystems such as dry deciduous forest and crassicaule shrubland at lower areas of the mountain range and oak-pine forests at higher elevations.
To assess the effects of global temperature rise at the local level, we applied ecological niche models to make projections for climate change scenarios considering different temperature rise levels. Historically, P. t. lagunae and S. o. lagunae are found in the oak-pine forest. Given their inaccessible and restricted habitat, there are very few records with unique localities (4 and 5, respectively, separated by less than one km between them); therefore, these two species were not candidates for use in ecological niche models because the models would not be statistically robust, they have to be considered a cluster of localities (Pawar et al. 2007). Instead, we used the tree species Pinus cembroides lagunae, Quercus brandegeei, and Q. devia, which are typical of the oak-pine forest in Sierra La Laguna. Also included were Peromyscus eva, a rodent species that inhabits lower elevations associated with the dry deciduous forest, and Chaetodipus rudinoris and C. spinatus, representative of arid ecosystems. These mammal species were used as references to examine their potential elevational displacement due to global climate change.
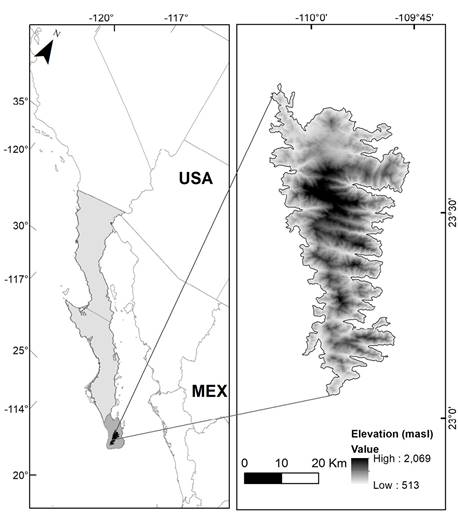
Figure 1 Geographic location of the sky island Sierra La Laguna, Baja California Sur, México. Light gray represents the Baja California peninsula. Medium gray represents the Cabo region. Dark gray represents Sierra La Laguna sky island.
The ecological niche models were developed using the Maximum Entropy (MaxEnt) algorithm in R (R Development Core Team 2020) for the present time using climatic variables from the Digital Climate Atlas of Mexico on a 250-meter scale (http://uniatmos.atmosfera.unam.mx/ACDM/servmapas). We used r < 0.80 to avoid collinearity and correlation (Dormann et al. 2013). The best models were selected based on good performance (< 10 % omission rate; Peterson et al. 2011), lowest value of the Akaike’s Information Criterion corrected for small sample size (AICc), and a ROC < 0.05 (Peterson et al. 2011). These were projected using three general circulation models: HadGEM2-ES (Jones et al. 2011), CCSM4 (Vertenstein et al. 2016), and MPI-ESM-LR (Giorgetta et al. 2013) considering a minimum global temperature rise of 1.5 °C and a maximum of 4.4 °C (for details of the modeling methodology, refer to Monroy-Gamboa et al. 2021).
Results
To assess the status of mammal species, two different analyses were conducted. The first included the associated plant species that define the oak-pine forest, i. e., Pinus cembroides, Quercus brandegeei, and Q. devia. The second was based on the fact that the temperature rise will lead to a forest desertification process with a shift toward xeric species.
The result of the modeling of P. c. lagunae, Q. brandegeei, and Q. devia showed a decrease in the area with climatic suitability under the three projected scenarios, except for Q. devia with the general circulation model HadGEM2-ES; with an increase of 4 °C the model showed a minimum increment (2.28 km2) in the surface (Figure 2; Table 1). Both the optimistic and pessimistic climate-change scenarios predicted the total disappearance of climatic suitability for the endemic population of P. c. lagunae, a species that is characteristic of the forest at higher elevations. The models for Peromyscus eva, associated with the dry deciduous forest, showed a loss of between 18 % and 21 % of the climatically suitable area (Figure 2; Table 1). The models for Chaetodipus rudinoris and C. spinatus, strongly associated with the xeric habitats in the lower part of the mountain range, showed a broadening of their elevational range between 108 to 682 and 1,295 m, respectively, and an expansion of the area with climatic suitability (Figure 2; Table 1). Both results indirectly show the reduction of the forest and the mammal species associated with it, which can be considered a collapse of the forest due to desertification.
Table 1 Area, in km2, presenting suitable climatic conditions for the species inhabiting the Sierra La Laguna sky island, currently and under three general circulation models as climatic change scenarios assuming minimum and maximum temperature rises of 1.5 ºC and 4.4 ºC, respectively.
| Current | Climate change | ||||||
|---|---|---|---|---|---|---|---|
| 1.5° C | 4.4° C | ||||||
| CCSM4 | HadGEM2-ES | MPI-ESM-LR | CCSM4 | HadGEM2-ES | MPI-ESM-LR | ||
| Pinus cembroides lagunae | 693.25 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Quercus brandegeei | 1,390.78 | 1,306.57 | 1,389.51 | 1,187.66 | 1,268.23 | 874.52 | 977.51 |
| Quercus devia | 1,378.36 | 614.23 | 1,130.76 | 743.40 | 303.86 | 1,380.64 | 1,052.88 |
| Chaetodipus rudinoris | 0.00 | 236.76 | 428.70 | 50.62 | 829.87 | 820.41 | 78.85 |
| Chaetodipus spinatus | 86.61 | 1,302.89 | 1,392.54 | 651.03 | 1,392.54 | 1,392.54 | 903.14 |
| Peromyscus eva | 1,345.34 | 1,050.28 | 1,098.43 | 1,097.39 | 1,095.74 | 1,096.99 | 1,099.00 |
| Sky island | 1,392.54 | ||||||
| Protected area | 1,124.50 |
General circulation models: CCSM4 (Vertenstein et al. 2016), HadGEM2-ES (Jones et al. 2011), and MPI-ESM-LR (Giorgetta et al. 2013).
Discussion
In the Sierra La Laguna Biosphere Reserve, there are no vehicle roads to reach the oak-pine forest; a private road reaches up to an elevation of 1,220 meters but allows no access to the public. Public vehicular accesses are located at the foothills of the mountain range up to 620 masl. The forest can only be accessed by walking through five specific points: La Burrera, San Dionisio, La Zorra, La Victoria, and San Pedro del Frijolar, through paths running across approximately 900 meters of elevation with very steep slopes. There are three points controlled by park rangers: two in the accesses most frequently used by visitors (La Burrera and San Dionisio) and a third at the destination site, the park ranger’s hut (Conanp 2015).
The only regular visitors to the reserve are park rangers and forest fire brigades; property and land owners visit the reserve occasionally to conduct monitoring and surveillance activities in the areas granted to them. In some seasons of the year, small groups of visitors register with park rangers; the activities and camping sites are restricted by PNA regulations that visitors meet. Sierra La Laguna is an area where the extraction of forest materials is prohibited, including dead wood, so there is neither tree cutting nor firewood collection. Also, there are no human settlements above ~600 masl, except for the forest station in La Laguna (for forest guards) and a hut owned by the Centro de Investigaciones Biológicas del Noroeste at Palo Extraño. Both facilities are for temporary use only and are reached by walking an average time of six and eight hours, respectively. Most of the human settlements within the reserve (around 65) are located between 400 and 600 masl and are single-family houses, with a very low percentage of permanent households. Most of these rural households are registered as inhabited (INEGI 2020).

Figure 2 Area and elevations with suitable climatic conditions for characteristic species of the sky island of Sierra La Laguna under climatic change scenarios. Rows show the increments of temperature and columns the general circulation models used in each case. The area representing the sky island is shown with a black outline, where the three characteristic species of trees (Pinus cembroides lagunae, Quercus brandegeei, and Q. devia) are currently distributed. Gray represents the suitable climatic area with climatic change scenarios for Q. brandegeei. Black represents the suitable climatic area with climatic change scenarios for two species: Q. brandegeei and Q. devia. Pinus cembroides lagunae is missing because this species lost its climatic suitability in all the climatic change scenarios).
There are no agricultural activities associated with the forest. Also, there is no goat or sheep farming, and bovine cattle farming is minimal; some feral farm animals have been recorded, which have escaped from extensive cattle raising carried out in the ranches located at lower elevations.
Results of our niche models show that the plant species typical of the oak-pine forest will face a decrease in their areas of optimal climatic conditions under temperature rises of 1.5 °C and 4.4 °C, projected from the three general circulation models. In all scenarios, Pinus cembroides lagunae loses the climatic suitability conditions that allow it to survive. The results of the model for Pinus show a reduction of its areas with optimal climatic conditions. This means that the expected recruitment of Pinus will be lower than its mortality rate, leading to a negative demographic trend; thus, the total number of Pinus individuals would decline gradually through time, leading to the increasing isolation of mature individuals until their ultimate disappearance. Although the models for Q. devia and Q. brandegeei suggest areas with climatic suitability for the different temperature-rise scenarios, the increase in global temperature will affect these species by reducing the area climatically suitable for their survival and reproduction. The results generally project that, in the short term, the vegetation will evolve from an oak-pine forest to an oak forest and then will entirely disappear. The historical records of Peromyscus truei lagunae and Sorex ornatus lagunae associate these species with sites where P. c. lagunae and Q. devia predominate (Woloszyn 1985). However, there is a high probability that these two tree species will lack the conditions of climatic suitability allowing them to survive if temperature rises as much as 1.5 °C and 4.4 °C; therefore, the species associated with them will certainly be highly affected.
It has been shown that suitable climatic conditions for xeric species will expand in area, reaching elevations where more temperate conditions currently exist (Monroy-Gamboa et al. 2021), so P. eva may populate the area where P. t. lagunae currently thrives.
The analysis of species associated with desert ecosystems shows an expansion of the area of distribution and elevational range, so the overall modeling suggests the replacement of forest species by species with xeric characteristics. Chaetodipus rudinoris and C. spinatus, species that are characteristic and closely associated with the desert, broaden their potential elevational range because niche models project the expansion of areas with xeric climates; this implies that these species will gradually colonize higher elevations and replace the species that inhabit current forests.
Worldwide, the five years with the highest temperatures since 1990 have been recorded from 2015, with 2020 being the year with the second-highest temperature. Indeed, temperature has increased by 1.25 °C, i. e., we have nearly reached the 1.5 °C rise that was formerly projected to occur not before 2050 (Ripple et al. 2021); it has now been envisaged that limiting the temperature rise to even 2 °C will be an unattainable goal (IPCC 2021). These facts, combined with the virtually absent local anthropic influence on the environment and the results of boreal plant species showing a collapse and the expansion of xeric mammals, allow hypothesizing that forest mammals such as S. o. lagunae and P. t. lagunae are undergoing an apparently irreversible local extinction process directly resulting from a climate change effect that causes variations in environmental parameters. In the present case, these will not be favorable for the oak-pine forest and will likely cause its extinction in the short term.
The results obtained indicate that even for an area characterized by restricted access and with low or no anthropogenic impact, the 1.5 °C and 4.4 °C temperature-rise scenarios will directly impact the oak-pine forest. Changes in the forest as a vegetation type will have an immediate impact on the associated fauna. The results of our analyses point to the possibility of local extinctions of P. t. lagunae and S. o. lagunae, mammal populations that are endemic to this sky island associated with different historical and climatic processes in the Baja California peninsula. The changes in climate that previously occurred historically have been of lower intensity and over longer periods of time, allowing these species to survive. By contrast, the current climate change is occurring faster than the capacity of these species to adapt; seemingly, the current rise in temperature driven by anthropic causes will inevitably lead to the local extinction of these species.

