











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El estudio de las causas y efectos que originan los disturbios forestales naturales y los provocados por el hombre son importantes para desarrollar estrategias de manejo sustentables. Las características actuales de un hábitat específico son producto de eventos naturales e inducidos que han ocurrido en el pasado (Kneitel, 2010), los cuales han modificado la diversidad, composición y estructura de los bosques bajo manejo (Kuuluvainen, 2002; Hernández et al., 2013).
Cuando se adoptan sistemas de manejo para la formación de bosques regulares, se plantea como objetivo el incremento de la producción y productividad de la madera, y se simplifica tanto la estructura como la diversidad de especies arbóreas (Castellanos et al., 2008). Esto es uno de los factores principales que aceleran los cambios y provocan la reducción de las funciones del ecosistema, así como la limitación del nivel de productividad del sitio a escala espacial y temporal (Monárrezet al., 2018).
Independientemente de la estructura deseada (regular o irregular), el manejo forestal debe considerar la producción de bienes y servicios de manera sostenible. El reto principal es la gestión y utilización de los bosques y terrenos forestales conservando la diversidad biológica, la capacidad productiva y la de regeneración a través de la aplicación de los tratamientos silvícolas (Aguirre, 2015).
De manera particular, el método de regeneración de matarrasa con plantaciones inmediatas promueve la renovación del rodal con taxones de rápido crecimiento, lo que mejora la riqueza y la diversidad arbórea (Návar y González, 2009). Sin embargo, además de la presencia de las especies plantadas, puede surgir el establecimiento espontáneo de todos los taxa de árboles que ya estaban ahí antes de la aplicación de dicha corta; ya sea a partir de la semilla almacenada en el suelo o de los individuos adyacentes al área de corta, así como por la capacidad de reproducción vegetativa de algunos de ellos (Smith et al., 1997).
Esta práctica silvícola incluida en los programas de manejo forestal se ha estado realizando en los bosques de Pinus y Quercus del estado de Durango y ha impactado en la biodiversidad de los bosques a nivel local y regional (Corral et al., 2005; Solís et al., 2006;). Por lo tanto, el objetivo de la presente investigación fue evaluar el efecto de las cortas de matarrasa en la diversidad arbórea de las plantaciones forestales comerciales en el ejido La Ciudad, Pueblo Nuevo, Durango.
Materiales y Métodos
Área de estudio
El estudio se realizó en el ejido La Ciudad, en el municipio Pueblo Nuevo, estado de Durango, que pertenece a la Unidad de Manejo Forestal 1008 “El Salto”, ubicada en el macizo montañoso de la Sierra Madre Occidental (Figura 1). El clima en el lugar es semifrío subhúmedo con lluvias en verano, temperatura media anual entre 5 °C y 12 °C, la temperatura del mes más frío varía de -3 °C a 18 °C, con un porcentaje de precipitación invernal de 5 a 10.2 % (García, 1973). La Carta Edafológica Serie II indica que el suelo se clasifica como Regosol (INEGI, 2010) y la vegetación arbórea está compuesta por especies de los géneros Pinus y Quercus (Carta Uso de Suelo y Vegetación Serie VI).
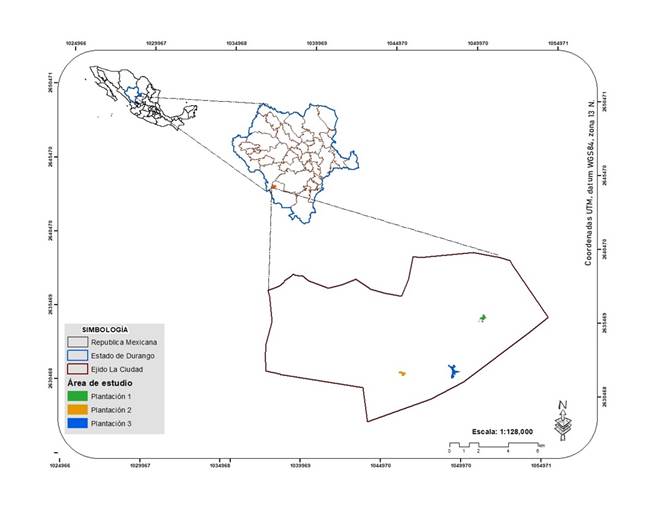
Selección de la muestra
El muestreo incluyó la selección de tres áreas, en las que se aplicaron cortas de matarrasa con plantación inmediata en el verano del año 2010, y los rodales adyacentes (RA) con vegetación arbórea establecida de manera natural, que representan las condiciones que existían dentro de las áreas de plantaciones antes de la aplicación de las cortas de matarrasa. Las especies plantadas a espaciamientos de 3 × 3 m fueron Pinus cooperi Blanco y Pinus durangensis Martínez (50 % para ambas especies), provenientes de viveros de la región de estudio, con un año de edad, además son las de mayor crecimiento y valor económico en la región. Las áreas de plantación se denominaron: plantación 1 (P1), con una superficie de 10.29 ha; plantación 2 (P2), con 12.45 ha; y plantación 3 (P3), con 9.43 ha. A partir del centro de cada plantación y en dirección a los puntos cardinales y subcardinales, se ubicaron 20 sitios circulares de 150 m2 a una distancia de 25 m entre ellos (60 sitios).
Con la finalidad de evaluar los cambios que ocurrieron en la diversidad arbórea de las plantaciones, se hizo una comparación de esta con la vegetación de los RA, los cuales se denominaron RA1, RA2 y RA3. En función de los mismos rumbos de las plantaciones hacia los RA, se localizaron dentro de cada uno 10 sitios de 0.1 ha (30 sitios).
Tanto en los sitios de las plantaciones, como de los RA, se identificaron todos los árboles por género y especie. Para el caso de las plantaciones se midió el diámetro a la base con un calibrador Truper ® Caldi-6MP, y para los RA el diámetro normal a la altura de 1.30 m con una forcípula Haglöf Mantax 59722, esto debido a que los RA presentaban estructuras diamétricas regulares, con edades cercanas al turno y, por lo tanto, existe escasa presencia de renuevos.
Análisis de datos
A partir de la información del número de taxones nuevos por sitio, se obtuvieron las curvas de riqueza acumulada, para justificar que el esfuerzo de muestreo fuera suficiente e incluyera la mayoría de las especies presentes tanto en las plantaciones, como en los RA (Dzib et al., 2014).
La descripción y comparación de la diversidad alfa de las plantaciones y de los
RA, se realizó con los índices de Riqueza de especies (S) y el
de Diversidad de Shannon-Wiener (
Para evaluar la ocurrencia de cambios significativos en S y en
Índices de similitud
El nivel de reemplazamiento de especies, originado por la aplicación de la corta
de matarrasa con plantaciones frente a los RA, se evaluó con los índices de
similitud cualitativos de Jaccard (
Donde:
El intervalo de valores de ambos índices varía de cero, cuando no hay especies
compartidas hasta uno, cuando los dos sitios comparten las mismas especies. El
Índice de Valor de Importancia
El Índice del Valor de Importancia de especies (IVI) se estimó
para evaluar la existencia de cambios estructurales en la composición de los
taxones a consecuencia de la aplicación de la corta de matarrasa con plantación.
El IVI, desarrollado por Curtis
y McIntosh (1951) consiste en sumar los valores relativos de densidad (
Donde:
La
La
Resultados y Discusión
Riqueza y diversidad
El número de especies en una comunidad determinada es una expresión mediante la cual se obtiene una estimación rápida y sencilla de la diversidad (Jiménez, 2000). En la Figura 2 se observa el punto de inicio de la asíntota superior de las curvas de acumulación de especies, por lo que se deduce que el tamaño de muestra fue suficiente para registrar la riqueza de especies presentes tanto en las plantaciones, como en los rodales adyacentes (Magurran, 2004).
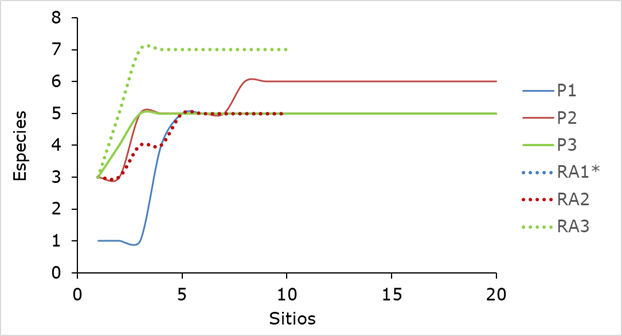
*La curva RA1 no se aprecia porque es idéntica a la curva RA2.
Figura 2 Curvas de acumulación de especies del muestreo de las tres plantaciones y rodales adyacentes.
Dentro del área de estudio (plantaciones y rodales adyacentes) se registró un total de ocho taxones distribuidos en los géneros Pinus, Quercus, Juniperus y Arbutus. Entre la P3 y el RA3 se identificó la mayor diferencia de especies (dos); P2 y RA2 discreparon por una especie, y no se obtuvieron diferencias entre P1 y RA1 (Cuadro 1). En términos generales, la riqueza de especies total registrada en esta investigación es mayor a las siete citadas por Hernández et al. (2013) para los bosques de Chihuahua. De igual manera, Návar y González (2009), en un estudio realizado en bosques templados de Durango, calcularon de 7 a 7.8 especies en promedio en áreas sin tratamiento silvícola (0 % de remoción). Asimismo, determinaron una disminución gradual en el número de taxa a medida que aumentaba el porcentaje de remoción del área basal, hasta 5.7 especies en promedio con el tratamiento de matarrasa (100 % de remoción).
Cuadro 1 Especies presentes en las plantaciones y rodales adyacentes en la región de Pueblo Nuevo, Durango.
| No. | Especie | P1 | P2 | P3 | RA1 | RA2 | RA3 |
|---|---|---|---|---|---|---|---|
| 1 | Arbutus xalapensis Kunth | 1 | 1 | 1 | 0 | 0 | 1 |
| 2 | Juniperus deppeana Steud. | 1 | 1 | 1 | 1 | 1 | 1 |
| 3 | Pinus cooperi Blanco | 1 | 1 | 1 | 1 | 1 | 1 |
| 4 | Pinus durangensis Martínez | 1 | 1 | 1 | 1 | 1 | 1 |
| 5 | Pinus engelmannii Carr. | 0 | 0 | 0 | 1 | 1 | 0 |
| 6 | Pinus leiophylla Schiede ex Schltdl. & Cham. | 0 | 0 | 0 | 0 | 0 | 1 |
| 7 | Quercus rugosa Née | 0 | 1 | 0 | 0 | 0 | 1 |
| 8 | Quercus sideroxyla Humb.& Bonpl. | 1 | 1 | 1 | 1 | 1 | 1 |
| Total de especies | 5 | 6 | 5 | 5 | 5 | 7 |
P1 = Plantación 1; RA1 = Rodal adyacente 1; P2 = Plantación 2; RA2 = Rodal adyacente 2; P3 = Plantación 3; RA3 = Rodal adyacente 3; 1 = Presente; 0 = Ausente.
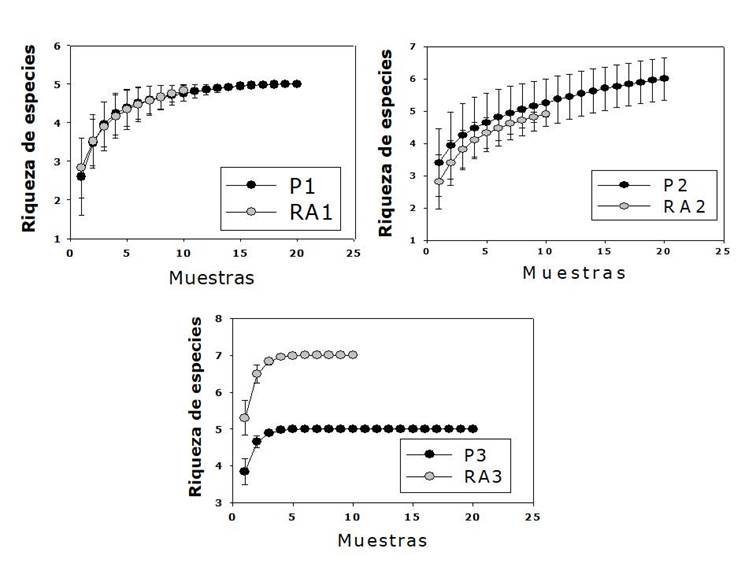
Las curvas de rarefacción indicaron que la riqueza de especies de la P3 fue significativamente menor a la del RA3. Esto coincide con los estudios de Corral et al. (2005) y Vásquez-Cortez et al. (2018), quienes documentan que el número de especies que se comparte puede disminuir en un área, al realizar tratamientos silvícolas. Las curvas de P1 y P2 con sus rodales adyacentes evidenciaron que la riqueza de especies tiende a mantener significativamente el mismo número de taxa, en relación con los rodales adyacentes (Figura 3).Al respecto, Leyva-López et al. (2010) y Hernández et al.(2013) señalan que la aplicación de cortas intensivas para propiciar la regeneración (árboles Padre), no garantiza solo el establecimiento del renuevo de los árboles del género seleccionado como árbol Padre, debido a que dentro de las áreas intervenidas existen otras especies con mecanismos diferentes de regeneración. En este estudio, tanto en la P1 como en la P2 además de los taxones del género Pinus plantadas, se confirmó el surgimiento de otras especies de pino.
En la Figura 4 se observa que no existe traslape en los intervalos de confianza en el Índice de Diversidad Proporcional de Shannon entre las plantaciones P2 y P3 con sus respectivos rodales adyacentes, lo que muestra que existen diferencias significativas entre la diversidad forestal de esas plantaciones y sus rodales adyacentes. Entre P1 y RA1 no se observaron cambios significativos, lo que indica que tanto la riqueza de especies como en su proporción fueron similares.
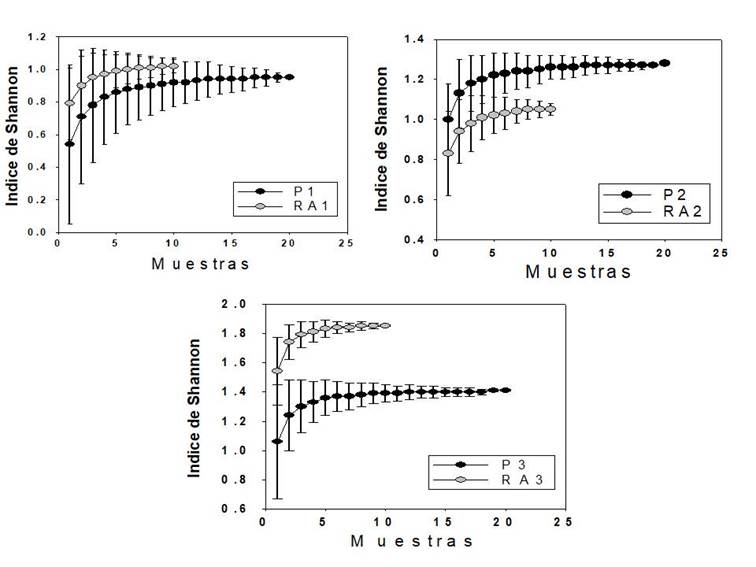
Figura 4 Graficas de rarefacción construidas para la diversidad proporcional de Shannon-Wiener (95 % de confianza).
Para P2 y su RA, las curvas de rarefacción basadas en la riqueza de especies no
confirmaron diferencias significativas (Figura
3). Sin embargo, si existieron para
Índices de similitud
Al evaluar la similitud de las especies presentes en las plantaciones con los rodales adyacentes, los índices de Jaccard y Sörensen permitieron deducir que la composición de especies entre la P3 y el RA3 tuvo mayor similitud (cinco especies en común), en comparación con las otras plantaciones y sus rodales adyacentes, las cuales presentaron menor similitud (Cuadro 2).
Cuadro 2 Índices de similitud de las plantaciones y sus rodales adyacentes.
| P vs RA | I J | I S |
|---|---|---|
| 1 | 0.66 | 0.80 |
| 2 | 0.57 | 0.72 |
| 3 | 0.71 | 0.83 |
P = Plantación; RA = Rodal adyacente; I J = índice de similitud de Jaccard; I S = Índice de similitud de Sorensen.
Del total de las especies registradas en el área de estudio: Pinus durangensis, Pinus cooperi, Quercus sideroxyla Humb.& Bonpl. y Juniperus deppeana Steud. estuvieron presentes tanto en las plantaciones, como en los RA (Cuadro 1). Con relación a las especies registradas en las áreas tratadas con cortas de matarrasa, la mayoría de los individuos de P. durangensis y P. cooperi provienen principalmente de la plantación, mientras que Quercus sideroxyla, Q. rugosa Née, J. deppeana y Artus xalapensis Kunth de las semillas contenidas en el suelo o por reproducción vegetativa. En este sentido, la similitud de especies y la diferencia en el número de ellas puede explicarse a partir de las estrategias de establecimiento y desarrollo que cada taxón presenta ante el tipo y grado de disturbio a los que fue expuesto (Leyva-López et al., 2010).
Por otra parte, Hernández et al.
(2013) determinaron que conforme prospera la masa forestal después de
ser intervenida, se comparte un número menor de especies, pero su riqueza en el
bosque se mantiene. Lo anterior coincide con este estudio, aunque la similitud
entre la P2 y el RA2 fue de 0.57 para
Índice de Valor de Importancia
Si se considera a los RA como la condición original y que fue modificada por las plantaciones, P. cooperi se mantuvo como el taxón de mayor IVI en la P2 y RA2; de la misma manera, P. durangensis no presentó cambios en el IVI entre la P3 y RA3, pero en la P1 desplazó con el mayor IVI a P. cooperi del RA1. En contraste, destaca la ausencia de P. engelmannii Carr. y P. leiophylla Schiede ex Schltdl. & Cham. en las plantaciones, respecto a sus rodales adyacentes donde tenían presencia (Cuadro 3).
J. deppeana no registró modificaciones considerables en el IVI en ninguna de las plantaciones. Q. sideroxyla aumentó su posición del IVI en las tres plantaciones y A. xalapensis se observó en la P1 y P2, pero no se le identificó en sus RA (Cuadro 3).
De manera general, se esperaría que en las plantaciones solo existieran P. durangensis y P. cooperi en la misma proporción que fueron plantadas (50 % para cada especie). Sin embargo, es importante señalar que tanto estas, como otras nativas pueden emerger de manera espontánea por la semilla depositada en el suelo antes y durante la corta, la semilla de rodales adyacentes o de forma vegetativa, a partir de las nuevas condiciones que se presentan en cada área. Con base en lo anterior, Oliver y Larson (1996) y Leyva-López et al. (2010) citan que la presencia de un mayor o menor número de taxones dentro de las áreas estudiadas se debe a la capacidad de cada una de ellas de reproducirse o regenerarse de manera vegetativa; así como a su vulnerabilidad al nivel de disturbio, estrés y competencia. Además, Vásquez-Cortez et al. (2018) reconocen que las áreas aledañas a las matarrasas limitan la dominancia de las plantas y la pérdida de diversidad arbórea y arbustiva.
Cuadro 3 Índice de valor de importancia ecológica de las especies.
| Especies | P1 | RA1 | P2 | RA2 | P3 | RA3 |
|---|---|---|---|---|---|---|
| Arbutus xalapensis Kunth | 51.98 | 0.00 | 31.34 | 0.00 | 37.46 | 48.72 |
| Juniperus deppeana Steud. | 50.98 | 75.97 | 75.99 | 84.37 | 60.07 | 50.63 |
| Pinus cooperi Blanco | 17.14 | 116.84 | 92.39 | 119.51 | 56.55 | 43.57 |
| Pinus durangensis Martínez | 130.18 | 47.80 | 39.65 | 51.56 | 91.17 | 52.28 |
| Pinus engelmannii Carr. | 0.00 | 21.64 | 0.00 | 17.63 | 0.00 | 0.00 |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 29.10 |
| Quercus rugosa Née | 0.00 | 0.00 | 1.95 | 0.00 | 0.00 | 30.94 |
| Quercus sideroxyla Humb.& Bonpl. | 49.70 | 37.74 | 58.64 | 26.90 | 54.72 | 44.72 |
| Sumatoria | 300 | 300 | 300 | 300 | 300 | 300 |
P1 = Plantación 1; RA1 = Rodal adyacente 1; P2 = Plantación 2; RA2 = Rodal adyacente 2; P3 = Plantación 3; RA3 = Rodal adyacente 3.
Con respecto a Juniperus,Rzedowski (1981); Bakker et al. (1996) y Biondi y Ferrante (2004) precisaron que tiene la capacidad de colonizar lugares degradados y forma parte de la vegetación secundaria; es de aparición temprana y su presencia se asocia a disturbios originados por actividades humanas. Lo que sugiere una posibilidad de repoblación mayor y desarrollo del género cuando el nivel de competencia y sombra se reduce, significativamente, por las intervenciones silvícolas. De igual manera, Leverkus et al.(2014) documentaron que las especies de Quercus rebrotan de forma muy favorable en áreas incendiadas y degradadas, lo que les confiere resiliencia a los bosques que conforman.
Por otra parte, aunque existe escasa información sobre la capacidad de rebrote de Arbutus después de una corta, en el área de estudio se observó alta incidencia de nuevos ejemplares. Al respecto, Díaz-Hernández et al. (2014) confirmaron una elevada probabilidad de rebrote de Arbutus en una zona con disturbio ocasionado por un incendio forestal.
Conclusiones
Los resultados de este estudio permiten concluir que las cortas de matarrasa con plantaciones realizadas en bosques mezclados mantienen la riqueza de especies; sin embargo, la diversidad si es afectada significativamente en dos de las tres plantaciones estudiadas, con respecto a sus rodales adyacentes.
A pesar de que en las cortas de matarrasa se hace una remoción total de todas las especies arbóreas y solo se plantaron ejemplares de P. durangensis y P. cooperi, la similitud de especies entre las plantaciones y los rodales adyacentes es alta, debido al surgimiento espontáneo de otros taxones nativos presentes antes de la corta y a la influencia de los rodales adyacentes. Esto propicia que el Índice de Valor de Importancia de las especies mantuviera una estructura, jerarquía y dominancia similar entre los taxa de las plantaciones y los rodales adyacentes.

