










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.3 no.3 Mérida jul./sep. 2012
Artículos
Heterosis y aptitud combinatoria para producción y calidad de forraje en seis poblaciones de maíz
Heterosis and combining ability for forage yield and quality in six maize populations
Alfonso Peña Ramosa, Fernando González Castañedaa, Gregorio Núñez Hernández*b, Ma. Del Rosario Tovar Gómezc, Víctor A. Vidal Martínezd, José Luis Ramírez Díaze
a Campo Experimental Pabellón, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). Km 32.5 Carr. Ags-Zac, A.P. 20660, Pabellón de A., Aguascalientes, México. pena.alfonso@inifap.gob.mx. Correspondencia al primer autor.
b Campo Experimental La Laguna, Centro de Investigación Regional Norte Centro del INIFAP. Torreón, Coah., México.
c Campo Experimental Santa Lucía. Centro de Investigación Regional del Centro del INIFAP. Texcoco, Edo., de México.
d Campo Experimental Santiago Ixcuintla, Nay. Centro de Investigación Regional Pacífico Centro del INIFAP.
e Campo Experimental Altos de Jalisco. Centro de Investigación Regional Pacífico Centro del INIFAP.
Recibido el 11 de junio de 2010.
Aceptado el 28 de marzo de 2011.
RESUMEN
Seis poblaciones de maíz derivadas de fuentes de germoplasma sobresalientes por sus cualidades forrajeras y sus 15 cruzas directas se evaluaron en cuatro localidades entre 1,117 y 1,932 m sobre el nivel del mar. El objetivo fue determinar la aptitud combinatoria general y específica, y la heterosis media para variables de producción: materia seca total (MST), de elote (MSEL) y de rastrojo (MSRAS) y variables de calidad del forraje: digestibilidad in vitro de la materia seca (DIVMS), contenido de fibra detergente neutro (FDN) y proteína cruda (PC). Las poblaciones difirieron significativamente en las tres variables de producción, pero no en calidad forrajera; en cambio las cruzas difirieron en producción de MSEL y en DIVMS. Los efectos de aptitud combinatoria general (ACG) fueron significativos sólo para MST y MSEL; mientras que los efectos de aptitud combinatoria específica (ACE), lo fueron adicionalmente para DIVMS. La población P3 tuvo la mayor ACG para MST (0.66 t ha-1), y la población P5F para MSEL (0.49 t ha-1). La cruza precoz P1xP2 presentó la heterosis media más alta para MST con 17.6 % y efectos positivos y significativos de ACE para MST y DIVMS con valores de 1.48 t ha-1 y 22.6 g kg-1. En cambio, las cruzas P2xP5F, P1xP4 y P1xP6F tuvieron los valores de heterosis más altos para MSEL con valores de 18.1 a 26.2 %. Ninguna cruza tuvo heterosis media importante para caracteres de calidad de forraje.
Palabras clave: Zea mays L., Patrones heteróticos, Producción de forraje, Calidad de forraje, Cruzas dialélicas.
ABSTRACT
Six corn populations drawn from outstanding forage quality germplasm, and 15 of their direct crosses were assessed in four sites located between 1,117 and 1,932 m asl. The purpose of the present study was to determine general and specific combining ability and mid parent heterosis for whole plant dry matter (TDM), ear dry matter (EDM), stover dry matter (SDM), in vitro dry matter digestibility (IVDMD), neutral detergent fiber content (NDF) and crude protein (CP). Populations showed significant differences for all dry matter yield characteristics, but not for forage quality, while crosses showed differences for EDM and IVDMD. General combining ability effects (GCA) were high and significant only for TDM, while specific combining ability effects (SCA) were significant for IVDMD. Population P3 had the highest GCA for TDM (0.66 t ha-1) and P5F for EDM (0.49 t ha-1). Early cross P1*P2 had the highest heterosis for TDM (17.6 %) and highly significant SCA effects for both TDM and IVDMD (1.48 t ha-1 and 22.6 g kg-1, respectively), while crosses P2*P5F, P1*P4 and P1*P6F had high mid parent heterosis for EDM, with values ranging from 18.6 % to 26.2 %. None of the crosses showed mid parent heterosis for forage traits of importance.
Key words: Zea mays L., Heterotic patterns, Forage yield, Forage quality, Diallel crosses.
INTRODUCCIÓN
En México la producción de maíz forrajero se incrementó en los últimos dos años en más de 2.3 millones de toneladas(1) debido a una mayor demanda de las explotaciones lecheras. El creciente aumento en el uso de maíz forrajero exige poner mayor atención en estrategias de manejo agronómico, calidad del forraje y selección, que permitan aprovechar al máximo el potencial existente por medio de programas de mejoramiento genético.
El estudio de la habilidad combinatoria y heterosis se ha utilizado para la determinación de patrones heteróticos y ha sido esencial para lograr avances en la expresión genética de caracteres importantes en maíz(2). Los patrones heteróticos pueden ser arbitrarios y no se limitan a combinaciones dentado x cristalino o tropical x templado(3). Algunos estudios(4,5) revelaron mayores niveles de heterosis en cruzamientos entre germoplasma con endospermo cristalino x dentado, aunque varias combinaciones y poblaciones dentro de un mismo tipo de endospermo también mostraron niveles altos de heterosis. En otro estudio(6) se encontraron varios patrones heteróticos alternativos a BSSS x Lancaster Sure Crop para la Faja Maicera de los Estados Unidos. En México también se han identificado patrones heteróticos para rendimiento de grano entre germoplasma dentado x cristalino(7) y excelentes combinaciones entre germoplasma exótico x adaptado(8). Otras combinaciones buenas para rendimiento de mazorca se han identificado al cruzar híbridos de diferentes compañías semilleras(9).
Los patrones heteróticos se han enfocado principalmente al rendimiento de grano y muy poco al rendimiento de forraje; pero se ha encontrado que la combinación de germoplasma dentado x cristalino genera niveles de heterosis mayores para rendimiento de materia seca total y de la parte vegetativa, que combinaciones dentado x dentado o cristalino x cristalino(10). En este tipo de cruzamientos se han encontrado niveles de heterosis de 20 % para materia seca total y de 25 % para materia seca de elote, pero en ningún caso para digestibilidad de la materia seca total y del rastrojo(11). En otro estudio(10) tampoco se encontró heterosis para calidad del forraje; pero se determinó que líneas mejoradas de grano cristalino transmiten un mayor número de características relacionadas con la digestibilidad a los cruzamientos híbridos que líneas dentadas(12) y que la selección de líneas por atributos de calidad del forraje genera híbridos con mayor calidad nutritiva(13,14). También se ha observado que líneas de maíz de la Faja Maicera de Estados Unidos relacionadas al grupo heterótico amarillo dentado, generaron mejores combinaciones para producción de forraje y digestibilidad con líneas no tradicionales de grano cristalino de Argentina, que con la línea Mo17 derivada de Lancaster Sure Crop(15).
En México se han realizado pocos trabajos para determinar patrones heteróticos para producción de forraje, y los que hay no incluyen aspectos de calidad, debido a que los análisis de laboratorio son caros y en el mercado se paga principalmente por el volumen y no consideran primordial el criterio de la calidad. Latourneire et al(16) encontraron que la combinación Tuxpeño Bajío x Sintético Forrajero fue una de las mejores cruzas para producción de materia seca con heterosis de 14.6 %; mientras que en otro estudio(17) se identificaron combinaciones entre híbridos comerciales con producción de forraje superior a las 80 t ha-1 que pueden ser útiles en programas de mejoramiento genético. Las opciones que ofrece el mercado actual en híbridos forrajeros son limitadas, por lo cual se considera que debería darse especial atención a la generación de nuevos híbridos con las características de producción y calidad forrajera que sean más eficientes para la producción de leche y carne. Para avanzar en esta dirección, es necesario seleccionar fuentes de germoplasma con variación genética amplia en producción de materia seca, digestibilidad y contenido de fibras, y determinar patrones heteróticos que permitan un proceso de hibridación exitoso. El objetivo del presente trabajo fue determinar la heterosis media y la aptitud combinatoria para características de producción y calidad forrajera en seis poblaciones de maíz.
MATERIALES Y MÉTODOS
El material experimental fueron seis poblaciones de maíz del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) formadas con diferente base germoplásmica y ciclo de madurez; cuatro poblaciones contienen de 8 a 10 líneas S3 seleccionadas por su mayor producción de grano, resistencia al acame y sanidad de planta y de mazorca; las otras dos restantes se formaron con híbridos comerciales seleccionados por su alta calidad forrajera (Cuadro 1).
En el año 2004 en el Campo Experimental de Pabellón, Ags., se multiplicó semilla de las seis poblaciones y se hicieron las cruzas directas posibles en forma dialélica entre los seis progenitores. Las cruzas se formaron polinizando en forma manual alrededor de 60 plantas de una población con polen de 60 plantas tomadas al azar de la otra población. Las seis poblaciones progenitoras y sus 15 cruzas, se evaluaron en el ciclo agrícola de primavera-verano (PV) 2005, en cuatro localidades ubicadas en el norte centro y occidente de México: Pabellón, Ags.; Torreón, Coah.; Sta. María del Oro, Nay. y Tepatitlán, Jal. (Cuadro 2). En las primeras dos localidades se evaluó en condiciones de riego y en las otras dos, en temporal. En Pabellón Ags., el suelo es de textura franco arenosa, pH= 7.5, profundidad de 80 cm y 1.0 % de materia orgánica, en Santa María del Oro, Nay., el suelo es ultisol (café-rojizo) de textura franca arenosa, profundo (>1.0 m), pH muy ácido (3.9), y contenido de materia orgánica de 1.5 %; en Tepatitlán, Jal. el suelo es luvisol férrico (suelos rojos), de textura arcillosa, profundo (>1.0 m), con pH acido (5.6) y materia orgánica de 2.1 %; y Torreón, Coah. posee suelo de origen aluvial, de textura migajón arcilloso, pH de 7.5, con profundidad mayor a 2.0 m, con materia orgánica de 1.1 % y sin problemas de sales.
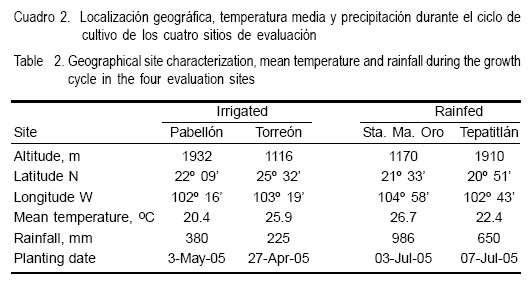
Cada experimento se sembró con un diseño experimental de bloques completos al azar con dos repeticiones; la unidad experimental fue de cuatro surcos de 5.0 m de largo y 0.76 m de separación. Se usó la densidad de población de 80,000 plantas ha-1; considerada como la más común usada por el productor. Al momento de la siembra se depositaron dos semillas por golpe cada 16.7 cm (seis golpes por metro), para aclarear a una planta después de la emergencia y dejar la densidad de población deseada. La dosis de fertilización fue 200N-90P-00K, la cual se aplicó en dos partes, la primera entre los 25 y 30 días después de la siembra con la mitad del nitrógeno y todo el fósforo, y la segunda entre los 50 y 60 días después de la siembra con el resto del nitrógeno. En todos los experimentos se aplicó herbicida preemergente al momento de la siembra para el control de la maleza. En Pabellón se dieron cuatro riegos de auxilio y en Torreón cinco; estos riegos fueron suficientes para un buen desarrollo del cultivo. En Tepatitlán, Jal., la precipitación fue de 650 mm, pero hubo sequía severa en la etapa de llenado de grano, de tal manera que la producción de mazorca fue baja; en Santa María del Oro, llovieron 986 mm; pero se tuvo algo de daño por acame debido a una tormenta severa.
La cosecha del forraje se hizo cuando la línea de leche en el grano alcanzó de un tercio a un medio de avance. El rendimiento de forraje se estimó en los dos surcos centrales de cada unidad experimental; para ello se cortó el total de plantas y se les midió peso fresco total y peso fresco de elotes. Además, se tomó una muestra al azar de cinco plantas completas y otra de cinco elotes, éstas se picaron y secaron en una estufa de aire forzado a una temperatura de 60 °C hasta alcanzar peso constante. Con estos datos se determinó la producción de materia seca total (MST) y materia seca del elote (MSEL) y por diferencia, se calculó la producción de materia seca de rastrojo (MSRAS). Las muestras secas de las cinco plantas se molieron en un molino Willey con una criba de 1 mm; una vez molidas, se les determinó el contenido de fibra detergente neutro (FDN)(18), digestibilidad in vitro (DIV)(19), y proteína cruda (PC) mediante espectroscopía de reflectancia en el infrarrojo cercano (NIRS). Además se estimó la correlación entre materia seca del elote y digestibilidad in vitro.
Se realizó un análisis de varianza conjunto para cada variable, en el cual la variación debida a genotipos se subdividió en progenitores, cruzas y progenitores vs cruzas, mediante el siguiente modelo lineal:
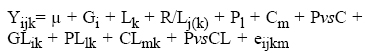
Donde Yijk= valor fenotípico observado del genotipo i, en la repetición j, anidada en la localidad k; μ= efecto común a todas las observaciones; Gi= efecto del i-ésimo genotipo; Lk= efecto de la k-ésima localidad; R/Lj(k)= efecto de la j-ésima repetición anidada en la k-ésima localidad; + P1= efecto del l-ésimo progenitor; Cm= efecto de la m-ésima cruza; PvsC= contraste progenitores vs cruzas; GLik= efecto de interacción genotipo x localidad; PLlk= efecto de interacción progenitor x localidad; CLmk= efecto de interacción cruza x localidad; PvsCL= contraste para la interacción progenitores vs cruzas x localidad y eijkm= efecto ambiental aleatorio correspondiente a la observación i,j,k,m. La comparación de medias se hizo mediante la prueba de diferencia mínima significativa (DMS) al 0.05 de probabilidad.
También se realizó un análisis de varianza combinado de las seis poblaciones y las 15 cruzas mediante el método 2 con efectos fijos de Griffing(20), el cual incluye en el análisis las cruzas directas y los padres. El modelo genético utilizado fue:
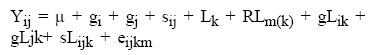
Donde i,j= 1, 2,., p padres; k= 1, 2,... , L localidades; m= 1, 2,..., r repeticiones; y los componentes del modelo: Yij= valor fenotípico observado de la cruza con los progenitores i y j, en la repetición m anidada en la localidad k; μ= efecto común a todas las observaciones; gi, gj= efecto de la aptitud combinatoria general de los progenitores i y j; sij= efectos de aptitud combinatoria específica de la cruza ij; RLm(k) =efecto de repeticiones anidadas en localidades; gLik= interacción del efecto de aptitud combinatoria general con la localidad; sLik= interacción del efecto de aptitud combinatoria específica con la localidad y eijkm= efecto ambiental aleatorio correspondiente a la observación i,j,k,m. Este análisis descompone las sumas de cuadrados de los genotipos en los efectos de ACG de los progenitores y los de ACE de las cruzas.
El análisis genético se hizo mediante el programa desarrollado por Castañón et al(21) mediante el procedimiento de SAS-IML(22). Tanto el modelo que incluye la partición de genotipos en progenitores y cruzas, como el modelo genético para ACG y ACE, se integraron en un análisis de varianza conjunto.
El error estándar (EE) de ACG y ACE se determinó mediante las ecuaciones siguientes:
EEGCA= √(((P - 1)/((P *(P + 2))*L* R)) σ2);
EEECA= √(((PΛ2 + P + 2)/(((P + Í)*(P + 2))*L*R)) σ2).
Donde P= progenitores; L= localidades; R= repeticiones y σ2= varianza del error conjunto. La significancia estadística de los efectos de ACG y ACE se probó mediante una prueba de t, dividiendo el valor del efecto entre su error estándar y comparándolo con el valor de t de tablas (P<0.05) con los grados de libertad del error conjunto. Usando las medias de los genotipos se calculó, para las variables de interés, el porcentaje de heterosis promedio de cada cruza; la cual se estimó con la ecuación siguiente: H= (F1-MP)/MP x 100. Donde: H= heterosis promedio; F1= valor de la cruza y MP= promedio de los progenitores.
RESULTADOS Y DISCUSIÓN
Análisis de varianza
El análisis de varianza combinado (Cuadro 3), muestra que las variables de producción: materia seca total, de elote y de rastrojo presentan significativamente mayor variación que las variables de calidad forrajera: digestibilidad in vitro, contenido de fibra detergente neutro y proteína cruda. Hubo diferencias significativas o altamente significativas entre localidades, genotipos, progenitores y efectos significativos para la interacción localidad x genotipos y localidad x cruza en los tres caracteres de producción. La fuente de variación progenitores vs cruzas, la cual en parte determina el grado de heterosis promedio de las cruzas (23) fue significativa sólo para materia seca total y de elote; aunque también se detectó interacción localidad x progenitores vs cruzas para estos mismos caracteres, sugiriendo que la heterosis promedio es afectada por el ambiente. En los caracteres de calidad, solamente la variación asociada con genotipos y cruzas fue significativa para digestibilidad in vitro y en ningún caso se detectó interacción significativa con localidades, sugiriendo que la expresión de este carácter fue estable a través de ambientes. En proteína cruda, ninguna fuente de variación fue significativa; lo cual contrasta con lo obtenido en otro estudio con poblaciones de maíz diferentes(16).
Con base en los valores de los cuadrados medios del análisis de varianza se determinó que en los caracteres de producción: materia seca total, materia seca de elote y materia seca de rastrojo la variación genética aditiva (ACG) fue mayor que la no aditiva (ACE); mientras que en los caracteres de calidad, la variación genética no aditiva fue ligeramente mayor que la aditiva; excepto en la variable digestibilidad in vitro (Cuadro 3).
En las variables de producción, los efectos de ACG y ACE fueron significativos sólo para materia seca total y de elote; mientras que en las variables de calidad, solamente los efectos de ACE fueron significativos para digestibilidad in vitro de la materia seca (Cuadro 3). Esto sugiere que en la producción de materia seca total y de elote los efectos genéticos aditivos y de dominancia son importantes; mientras que en la digestibilidad in vitro, lo son sólo los efectos de dominancia. Por lo anterior, se deduce que estos tres caracteres pueden ser mejorados exitosamente mediante programas de selección recíproca recurrente, debido a que en este método se capitalizan tanto los efectos aditivos como los no aditivos; a diferencia la producción de materia seca de rastrojo, contenido de fibras y proteína cruda; en los cuales se requeriría primero mejorar las poblaciones per se. De la Cruz et al(24) determinaron mayor contribución de los efectos de dominancia que de los aditivos para producción de materia seca, y a diferencia del presente trabajo no encontraron significación estadística en la fuente de variación ACE para digestibilidad y sí para el contenido de fibra detergente neutro; diferencias que pudieran ser explicadas primordialmente porque no se utilizó el mismo material genético en la investigación, y los atributos estructurales de la planta cambian de un genotipo a otro.
Las interacciones localidad x ACG y localidad x ACE fueron significativas para materia seca total y de elote, y en ningún caso para variables de calidad; esto sugiere que la respuesta de los progenitores en cruza para producción de forraje cambian de una localidad a otra; de ahí que sea importante identificar a aquellos progenitores y cruzas que expresen mayor consistencia en su expresión genética en una localidad dada; y en el caso de las variables de calidad, habrá que evaluar mayor número de ambientes para confirmar si la ausencia de interacción localidad x ACG y localidad x ACE se debe a que estas características son más estables o se trata de un sesgo debido al muestreo.
Producción de materia seca
Las poblaciones P3 y P4 tuvieron en promedio de localidades la mayor producción de materia seca total y de rastrojo con 18.9 y 13.0 t ha-1, respectivamente; mientras que la misma población P3 y la población P5F tuvieron mayor producción de elote con 6.5 y 6.1 t ha-1, respectivamente (Cuadro 4). La población P3 tuvo en Santa María del Oro y Tepatitlán, severos problemas de acame de raíz; lo que la hace menos deseable como fuente de mejoramiento. Las poblaciones precoces P1 y P2, fueron en cambio las más tolerantes al acame de raíz, pero de las menos productivas.
Catorce de las 15 cruzas tuvieron producciones de materia seca total similares estadísticamente entre sí (Cuadro 4). Seis de estas cruzas sobresalieron también en producción de materia seca de elote con rendimientos entre 6.4 y 7.0 t ha-1 y de ellas, cuatro tuvieron en común la población P5F y dos, la población P6F. Con excepción de la cruza P3 x P4, el resto tuvieron igual o mayor producción de materia seca total y de elote que el promedio de las poblaciones per se involucradas en las cruzas; indicando que manifiestan heterosis y que pueden ser fuentes apropiadas de mejoramiento genético para ambos caracteres. En promedio, las cruzas tuvieron mayor producción de materia seca total y de elote que las poblaciones. Existen resultados similares para producción de materia seca total en poblaciones de la Faja Maicera de Estados Unidos y de Europa donde encontraron que las mejores combinaciones se dieron entre poblaciones con endospermo dentado x cristalino (1 0). En el presente estudio no hubo un patrón heterótico definido en cuanto a la textura del grano, pero sí se observó una tendencia de mayor producción en aquellas cruzas que involucraron las poblaciones de alta calidad forrajera y de granos dentados.
Los efectos de ACG más altos y significativos para producción de materia seca total y de rastrojo, correspondieron a la población P3 con 0.66 y 0.48 t ha-1, respectivamente; en cambio para producción de materia seca de elote correspondió a la población P5F con 0.49 t ha-1 (Cuadro 5). Este resultado permite establecer que la población P3, fue la que más contribuyó a incrementar la producción de materia seca total y de rastrojo en sus cruzas; mientras que la población P5F, contribuyó a mejorar el peso seco de elote manteniendo sin cambios significativos la producción de la materia seca total en sus cruzas. Un mayor incremento en la producción de rastrojo con respecto a la producción de elote podría afectar negativamente la calidad del forraje, dado que el rastrojo es la parte de la planta con mayor contenido de fibras, y éstas contribuyen a que el forraje tenga menor digestibilidad(25). Por tanto, en el mejoramiento genético de maíz para la producción de forraje deben buscarse poblaciones que en cruza incrementen su producción de materia seca total mediante una mayor producción de elote, como se ha observado a través del mejoramiento de maíz en los Estados Unidos(26); este comportamiento se observó en la Población P5F, que en ciertas cruzas tuvo mayor producción de materia seca del elote sin cambios en la materia seca total.
Los efectos de aptitud combinatoria específica para los caracteres de producción de materia seca variaron significativamente entre cruzas, pero ninguna presentó efectos significativos favorables en todos (Cuadro 6); la cruza P1xP2, que individualmente en sus poblaciones tuvieron efectos negativos de ACG en producción de materia seca, presentó efectos significativos positivos de ACE para materia seca total y materia seca de rastrojo; mientras que la cruza P2xP5F para materia seca total y materia seca de elote. Las diferencias entre el comportamiento entre la ACG y ACE de las poblaciones P1, P2, P5F y sus cruzas P1xP2 y P2xP5F se explican porque en la ACE se expresa la interacción intra e interalélica y no la variación aditiva como en la ACG. La expresión de ACE es mayor cuando los alelos de las poblaciones tienen diferente origen genético y se manifiesta en niveles altos de heterosis; como los observados en las cruzas P1xP2 y P2xP5F en las variables señaladas (Cuadro 7). Al respecto, algunos estudios(10,11) revelan que los cruzamientos entre germoplasma dentado x cristalino tienen por lo general mayores respuestas heteróticas que dentado x dentado o cristalino x cristalino; mientras que en otros trabajos(8,15) se encontró que los niveles de heterosis más altos se detectaron con progenitores que provienen de zonas diferentes debido a mayor divergencia genética. En el presente estudio, las dos cruzas sobresalientes además de tener un progenitor de grano dentado y otro cristalino se formaron con germoplasma de diferente origen genético (Cuadro 1). Con base en estos resultados, se considera que ambas cruzas pueden ser buenos patrones heteróticos para lograr avances importantes en el mejoramiento genético para la obtención de híbridos exitosos. La cruza P1xP2, no fue una de las mejores en rendimiento (Cuadro 4), pero por ser más precoz (datos no presentados), es una buena alternativa para la región norte centro de México, donde la limitante de agua de riego es un fenómeno común. Por otra parte, la cruza P2xP5F adicionalmente a su alta aptitud combinatoria específica y heterosis media, tuvo altos rendimientos de materia seca total y de elote.
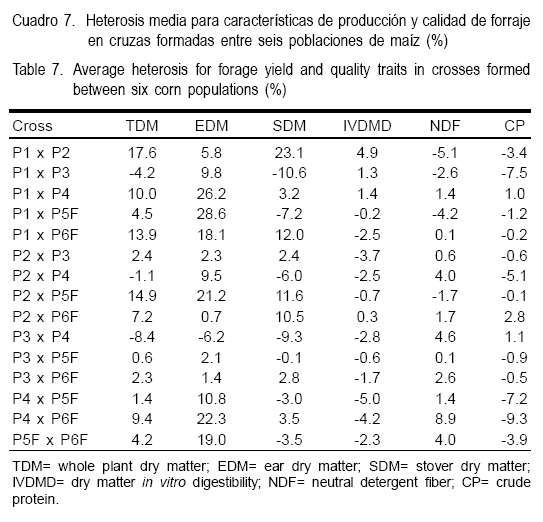
Las cruzas P1xP4, P1xP5F, P1xP6F y P4xP6F, mostraron una aptitud combinatoria específica significativa alta para rendimiento de materia seca de elote (Cuadro 6) y niveles de heterosis que variaron de 18.1 a 28.6 % (Cuadro 7). De estas cruzas sólo P1xP4 y P1xP6F tuvieron heterosis media para producción de materia seca total superior al 10 %, valor que es muy similar al obtenido en otros trabajos en la mejor cruza(16). Estas cruzas adicionalmente destacaron por su tolerancia al acame y sanidad a través de ambientes, de ahí que se les considere también como posibles fuentes de características que inciden en la producción de forraje.
Calidad forrajera
En calidad forrajera no hubo diferencia significativa entre las poblaciones, no obstante, que dos de ellas se formaron con germoplasma con alta digestibilidad de la materia seca y bajo contenido de fibras. La digestibilidad entre poblaciones varió de 666 a 695 g kg-1 y el contenido de fibra detergente neutro de 557 a 595 g kg-1 (Cuadro 4), con tendencias de las poblaciones P5F y P6F a tener mayor digestibilidad. En otro estudio se determinó mayor calidad forrajera en un grupo de poblaciones de maíz con variaciones en digestibilidad de 691 a 724 g kg-1 y en contenido de fibras de 497 a 524 g kg-1(25).
Hubo varias cruzas sobresalientes en digestibilidad de la materia seca (Cuadro 4), pero en general, todas fueron bajas e inferiores a 700 g kg-1, comparado con otros estudios donde se han obtenido valores de digestibilidad superiores a 750 g kg-1(27,28). La baja digestibilidad de las cruzas, se debió en parte a una reducida producción de materia seca de elote; lo anterior se corroboró con la correlación entre la materia seca de elote y la digestibilidad in vitro; ya que ésta fue positiva, alta y significativa (r= 0.84). Las cruzas sobresalientes fueron P1xP2 y varias cruzas en las cuales intervinieron las poblaciones P5F y P6F. En contenido de fibras y en proteína no hubo diferencias entre las cruzas.
Ninguna población tuvo efectos significativos favorables de aptitud combinatoria general, para las comerciales. Este resultado además de ser un indicador de falta de precisión en las estimaciones de ACG, reduce la probabilidad de detectar significación estadística en ellas. Probablemente incrementando el número de repeticiones por localidad, en la determinación de estas variables, mejoraría la confiabilidad de los efectos, pero el costo del experimento incrementaría sustancialmente.
Entre cruzas solamente P1xP2, presentó efectos significativos de aptitud combinatoria específica para digestibilidad con valores de heterosis de apenas 4.9 % (Cuadros 6 y 7). Pequeñas diferencias en la digestibilidad del forraje de maíz son importantes en la conversión a leche, de tal manera que a una misma producción de materia seca, diferencias en digestibilidad de 20 a 30 g kg-1 incrementan la producción de leche de una a dos toneladas por hectárea(27); por esta razón, la heterosis de 4.9 % observada en esta variable en la cruza P1xP2 puede considerarse importante y de utilidad para favorecer avances en un programa de hibridación. Moreno-González et al(10) no observaron heterosis importante en digestibilidad de la materia orgánica en un grupo de ocho poblaciones de maíz, pero sugieren que la selección recurrente para calidad forrajera tanto en poblaciones parentales como en las cruzas puede ser un medio apropiado para explotar posibles genes recesivos favorables. Se ha señalado(15), que la selección de características de calidad debería enfocarse principalmente al rastrojo; mientras que otros(30) destacan las ventajas de la selección recurrente para producción y calidad forrajera.
CONCLUSIONES E IMPLICACIONES
En los caracteres de producción la variación genética aditiva (ACG) fue mayor que la no aditiva (ACE); mientras que en los caracteres de calidad la variación no aditiva fue mayor que la aditiva; excepto en la variable digestibilidad in vitro; por lo que la selección por caracteres de producción será más exitosa. La población P3, tuvo efectos positivos y significativos de ACG para producción de materia seca total y de rastrojo, por lo que puede usarse como fuente de germoplasma para mejorar estos caracteres; la población P5F sólo para la producción de materia seca de elote; y ninguna de las poblaciones resultó adecuada como fuente de germoplasma para mejorar la calidad de forraje. La cruza formada con germoplasma dentado x cristalino, fue la única que tuvo efectos de ACE significativos y heterosis positiva, por lo que se considera un buen patrón heterótico para iniciar un programa de hibridación. Las cruzas P2xP5F, P1xP4 y P1xP6F podrían ser buenos patrones heteróticos para incrementar la producción de materia seca total o de elote, pero no la calidad del forraje.
LITERATURA CITADA
1. Servicio de Información Agroalimentaria y Pesquera (SIAP/ SAGARPA). Anuario Estadístico 2009. [en línea]. Disponible en http://www.siap.gob.mx. Consultada 14 Mar, 2010. [ Links ]
2. Hallauer AR, Miranda FJB. Quantitative genetics in maize breeding. 2nd ed. Iowa, USA: Iowa State University Press; 1988. [ Links ]
3. Soengas P, Ordás B, Malvar RA, Revilla P, Ordás A. Heterotic patterns among flint maize populations. Crop Sci. 2003;(43):844-849. [ Links ]
4. Beck DL, Vasal SK, Crossa J. Heterosis and combining ability of CIMMYT's tropical early and intermediate maturity maize (Zea mays L.) germplasm. Maydica 1990;(35):279-285. [ Links ]
5. Crossa J, Vasal SK, Beck DL. Combining ability estimates of CIMMYT's tropical late yellow maize germplasm. Maydica 1990;(35):273-278. [ Links ]
6. Melani MD, Carena MJ. Alternative maize heterotic patterns for the Northern Corn Belt. Crop Sci 2005;(45):2186-2194. [ Links ]
7. Ramírez DJL, Chuela BM, Soltero DL, Franco MJ, Morfín VA, Vidal MVA, Vallejo DHL, et al. Patrón heterótico de maíz amarillo para la región centro-occidente de México. Rev Fitotec Mex 2004;(27):1-17. [ Links ]
8. De la Cruz L, Ron PL, Ramírez DJL, Sánchez GJJ, Morales RMM, Chuela BM, Hurtado de la PSA, et al. Heterosis y aptitud combinatoria entre híbridos comerciales y germoplasma exótico de maíz en Jalisco, México. Rev Fitotec Mex 2003;(26):1-10. [ Links ]
9. De la Rosa LA, de León CH, Rincón SF, Martínez ZG. Efectos genéticos, heterosis y diversidad genética entre híbridos comerciales de maíz adaptados a El Bajío Mexicano. Rev Fitotec Mex 2006;(29):247-254. [ Links ]
10. Moreno-Gonzalez J, Martínez I, Brichette I, López A, Castro P. Breeding potential of European Flint and U.S. Corn Belt dent maize populations for forage use. Crop Sci 2000;(40):1588-1595. [ Links ]
11. Bertoia L, López C, Burak R. Biplot analysis of forage combining ability in maize landraces. Crop Sci 2006;(46):1346-1353. [ Links ]
12. Dhillon BS, Chr Paul, Zimmer E, Gurrath PA, Klein D, Pollmer WG. Variation and covariation in stover digestibility traits in diallel crosses of maize. Crop Sci 1990;(30):931-936. [ Links ]
13. Gurrath PA, Dhillon BS, Pollmer WG, Klein D, Zimmer E. Utility of inbred line evaluation in hybrid breeding for yield and stover digestibility in forage maize. Maydica 1991(36):65-68. [ Links ]
14. Argillier O, Méchin V, Barriere Y. Inbred line evaluation and breeding for digestibility-related traits in forage maize. Crop Sci 2000;(40):1596-1600. [ Links ]
15. Bertoia LM, Burak R, Torrecillas M. Identifying inbred lines capable of improving ear and stover yield and quality of superior silage hybrids. Crop Sci 2002;(42):365-372. [ Links ]
16. Latourneire L, Rodríguez S, de León H, Padrón E. Heterosis y aptitud combinatoria para rendimiento y calidad forrajera en poblaciones de maíz. Agronomía Mesoamericana 1996;(7):93-97. [ Links ]
17. Gutiérrez del RE, Espinoza BA, Palomo GA, Lozano GJJ, Antuna GO. Aptitud combinatoria de híbridos de maíz para la Comarca Lagunera. Rev Fitotec Mex 2004;27(1):7-11. [ Links ]
18. Van Soest PJ, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci 1991;(74):3583-3597. [ Links ]
19. Tilley JMA, Terry RA . A two-stage technique for the in vitro digestion of forages. J Brit. Grassland Soc 1963;(18):104-111. [ Links ]
20. Griffing B. Concept of general and specific combining ability in relation to diallel crossing systems. Aust J Biol Sci 1956;(9):463-493. [ Links ]
21. Castañón-Nájera G, Latournerie-Moreno L, Mendoza-Elos M. Macro de SAS-IML para analizar los diseños II y IV de Griffing. www.ujat.mx/publicaciones/uciencia2005. (21):27-35. [ Links ]
22. SAS Institute. SAS/IML Software: Usage and reference. Version 6. Ist. Ed. SAS Institute Inc., Cary, NC. 1990. [ Links ]
23. Gardner CO, Eberhart SA. Analysis and interpretation of the variety cross diallel and related populations. Biometrics 1966;(22):439-452. [ Links ]
24. De la Cruz LE, Rodríguez HSA, Palomo GA, López BA, Robledo TV, Gómez VA, Osorio OR. Aptitud combinatoria de líneas de maíz de alta calidad de proteína para características forrajeras. www.ujat.mx/publicaciones/uciencia. 2007;(23):57-68. [ Links ]
25. Peña RA, Núñez HG, González CF. Potencial forrajero de poblaciones de maíz y relación entre atributos agronómicos con la calidad. Téc Pecu Méx 2002;(40):215-228. [ Links ]
26. Lauer JG, Coors JG, Flannery PJ. Forage yield and quality of corn cultivars developed in different eras. Crop Sci 2001;(41):1449-1455. [ Links ]
27. Peña RA, González CF, Núñez HG, Jiménez GCA. Aptitud combinatoria de líneas de maíz de alta calidad forrajera. Rev Fitotec Mex 2004;27(1):1-6. [ Links ]
28. Coors JG, Albrecht KA, Bures EJ. Ear-fill effects on yield and quality of silage corn. Crop Sci 1997;(37):243-247. [ Links ]
29. Goncalves Ch L, Vieira MG, Vagno de SL, Pereira GO, Silva OJ. Parental commercial maize selection for silage production. Rev. Brasileira de Milho e Sorgo, 2008;7(2):183-194. [ Links ]
30. Frey TJ, Coors JG, Shaver RD, Lauer JG, Eilert DT, Flannery PJ. Selection for silage quality in the Wisconsin quality synthetic and related maize population. Crop Sci 2004;(44):1200-1208. [ Links ]

