










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.3 no.1 Mérida ene./mar. 2012
Notas de investigación
Parámetros genéticos de pesos al nacer y al destete en ovinos de raza criolla de Chiapas
Genetic parameters for birth and weaning weights in the local Chiapas sheep breed from Mexico
Reyes López-Ordazª, Iván Olivera-Vegaª, José M. Berruecos Villalobos3, Marisela Peralta-Lailsonb, Raúl Ulloa-Arvizuª, Carlos G. Vásquez Peláezª
ª Departamento de Genética y Bioestadística, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México. Circuito Exterior, Ciudad Universitaria. Coyoacán. C.F. 04510. México. Tel.: +56 22 58 94; Fax: +56 22 59 56 carlosgv@servidor.unam.mx. Correspondencia al último autor.
b Instituto de Estudios Indígenas, Universidad Autónoma de Chiapas. Centro Universitario, Campus III.
Resumen
Heredabilidades para pesos al nacer (PN) y destete (PD) en ovinos de la raza Chiapas se estimaron a partir de los registros de corderos nacidos entre 1991 y 2006. El pedigrí incluyó 53 carneros, 523 ovejas y 1,326 corderos. Se compararon seis modelos utilizando el Criterio de Información Akaike (Akaike Information Criterion, AIC). El modelo 1 incluyó los efectos genéticos directos aditivos. El modelo 2 igual al modelo 1 más los efectos permanentes del medio ambiente. Los modelos 3 y 4 fueron como el modelo 1 más los efectos maternos con cov(directa, maternal) (σam) igual a 0 para el modelo 3, mientras que para el modelo 4, σam ≠ 0. Los modelos 5 y 6 corresponden a los modelos 3 y 4 adicionados con efectos ambientales permanentes. Los efectos de año, tipo y número de parto y sexo fueron incluidos en todos los modelos. La edad al destete fue agregada como covariable en la estimación de peso al destete. Los análisis mostraron al modelo 6 como el más adecuado para ambos caracteres, siendo el más completo y el criterio de comparación fue el menor. Los valores obtenidos de heredabilidad directos y maternos, así como los correspondientes a efectos maternos ambientales permanentes fueron 0.15 ± 0.09, 0.24 ± 0.09, 0.11 ± 0.08 para PN y 0.14 ± 0.09, 0.09 ± 0.08, 0.08 ± 0.06 para PD, respectivamente, siendo las correlaciones fenotípica 0.34±0.03 y genética 0.81±0.18. Estos son los primeros valores obtenidos de parámetros genéticos para PN y PD en la raza Chiapas.
Palabras clave: Borrego Chiapas, Efectos directos, Crecimiento, Parámetros genéticos, Efecto materno.
Abstract
Variance components and heritabilities for birth (BW) and weaning weights (WW) were estimated in the local Chiapas sheep breed using data from animals born between 1991 and 2006. The pedigree contained 53 rams, 523 ewes, and 1,326 lambs. Six models were compared to estimate heritabilities of BW and WW using the difference (Δi) between the ith model and the best model from the Akaike information criterion (AIC). Model 1 contained additive genetic effects. Model 2 was as Model 1 plus permanent maternal environmental effects. Models 3 and 4 were as Model 1 plus maternal effects with cov(direct, maternal) equal to zero 0 for Model 3 and equal to σam for Model 4. Models 5 and 6 were as Models 3 and 4 plus permanent environmental effects. Fixed effects of year, type and number of birth and sex were included in all models for both traits. Age at weaning was added as covariate for WW models. Model 6 was preferred for both traits because it accounted for direct and maternal effects and their correlation as well as maternal permanent environmental effects and the criteria for comparison were the smallest. Direct and maternal heritability as well as the permanent maternal environment estimates were 0.15±0.09, 0.24±0.09, 0.11±0.08 for BW and 0.14± 0.09, 0.09 ±0.08, 0.08±0.06 for WW, respectively, and the phenotypic (0.34±0.03) and genetic (0.81±0.18) correlation from model 6. These are the first estimates of genetic parameters for BW and WW in the Chiapas sheep breed.
Key words: Chiapas sheep, Direct effect, Growth, Genetic parameter, Maternal effect.
En los Altos de Chiapas, la etnia Tzotil explota una raza ovina característica de esa región. Esta raza local, llamada Chiapas, es descendiente de animales de las razas Churra, Lacha y Manchega introducidos en esa región en el siglo XVI Pese a su gran importancia socio-cultural y económica en dicha región(2,3), no se cuenta con información genética disponible sobre caracteres de crecimiento para programas de mejoramiento.
El crecimiento pre-destete es función del potencial genético del cordero, la influencia materna (genética en el caso de la oveja y ambiental en el cordero) y otros efectos ambientales. Estos efectos pueden ser cuantificados por medio de modelos adecuados. Algunos autores(4,5) han evaluado parámetros genéticos para efectos directos, maternos y ambientales permanentes desde el nacimiento al destete en varias razas ovinas, y comprobaron que la correlación entre los efectos directos y maternos es desfavorable. Por otra parte, la heredabilidad de efectos aditivos directos, depende del modelo utilizado para calcularla(6,7,8). El objetivo de este estudio fue estimar los parámetros genéticos de pesos al nacimiento (PN) y destete (PC) en la raza Chiapas por medio de diferentes modelos genéticos. Estos valores de heredabilidad contribuirán a elaborar un programa de mejoramiento genético para esta raza.
Datos y manejo
Este estudio se realizó utilizando información registrada en el Centro de Investigación y Transferencia de Tecnología de la Universidad Autónoma de Chiapas (CUITT, UNACH) ubicado en "los Altos" de Chiapas, a 1,780 msnm, a 16°32'24" N y 92°28'19" O. El clima es templado frío con veranos cortos y frescos. La temperatura media anual es de 13 °C y la precipitación promedio de 1,059.2 mm anuales(9). Los animales pastorean diariamente entre 6 y 8 h praderas de pasto kikuyu (Pennisetum clandestinum) y se encierran hasta el día siguiente. Se les suministra un suplemento basado en planta completa de maíz (elote y planta seca), pasta de soya, sales minerales y aditivos. El manejo sanitario incluye una desparasitación mensual para control de Fasciola hepática, Moniezia expansa, parásitos gastrointestinales, Oestrus ovis y coccidiosis. Si se presentan infecciones o neumonía se los trata con oxitetraciclina. Curante la época de empadre, se formaron grupos de 10 a 15 ovejas y estuvieron con un solo semental por corral durante 60 días a partir de principios de junio. La parición tiene lugar desde noviembre a fines de diciembre y los corderos se destetan a los 3 meses.
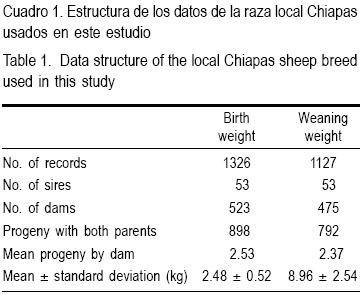
Se pesaron al nacer (PN) todos los corderos incluidos en este estudio (1,327) y un total de 1,127 al destete (PC). Se registró la información del pedigrí y la estructura de los datos se muestra en el Cuadro 1.
Análisis genéticos y estadísticos
Los pesos al nacimiento y al destete fueron analizados por medio de seis modelos lineales de una sola característica. Los efectos fijos fueron tipo del parto (simple, doble), número de parto de la madre (1,2,3,4,>5), año de nacimiento (19912006) y sexo de la cría (hembra, macho). El 36 % del total de madres fue primípara. La edad al destete en días (89 ± 22) fue incluida como covariable para peso al destete. Se empleó la prueba de Tukey para comparar las diferencias entre medias del número de parto de la madre. Las interacciones de dos vías no fueron significativas (P>0.05), y por lo tanto fueron eliminadas del modelo final(10).
Los componentes de varianza y covarianza fueron calculados por medio de la metodología REML, utilizando el programa ASReml(11). Los modelos fueron ajustados para estimar los parámetros genéticos para cada característica, incluyendo o eliminando distintas combinaciones de efectos genéticos maternos o materno–ambientales permanentes, tales como:
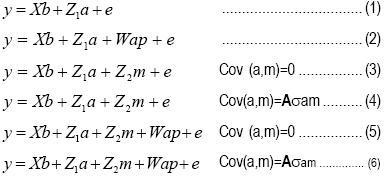
Donde:
y es el vector de los registros para características de PN y PC; b es el vector de efectos fijos del modelo final (tamaño del parto, año de nacimiento y sexo de las crías y edad al destete como covariable de PC); a es el vector de efectos genéticos aditivos del cordero; m es el vector de efectos genéticos maternos aditivos; ap es el vector de los efectos materno–ambiental es permanentes; X, Z, W son matrices de incidencia que relacionan los registros del vector y a los efectos en los vectores a, m y ap; siendo e el vector de efectos aleatorios de los residuos.
La matriz de varianza–covarianza de efectos aleatorios se define de la siguiente manera:
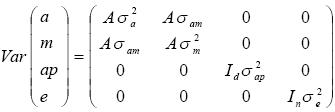
Donde:
A es la matriz de efectos genéticos directos aditivos; σa2 es la varianza de los efectos genéticos aditivos; σm2 es la varianza efectos maternos aditivos; σam es la covarianza entre efectos directos y maternos permanentes; σ2ap es la varianza de efectos materno ambientales permanentes; σe2 es la varianza residual que representa los efectos aleatorios tanto de ambiente temporal como desconocidos; siendo Id e In matrices identidad de orden adecuado, d= número de madres y n = número de animales con registro.
Para comparar los modelos se utilizó el criterio de información de Akaike (AICi = logL-K), logL es logaritmo de cociente de verosimilitud del modelo i y K es el número de parámetros del modelo; y las ponderaciones de Akaike (wi) como la probabilidad que el modelo i sea el mejor para la información disponible; wi= LMi/Σ LMi, donde LMi = exp(AICi - AICmin), y AICmin es el valor mínimo de los seis AICi. Por lo tanto, Δi = AICi-AICmin corresponde a la pérdida de información experimentada al usar el modelo i ajustado(12).
Finalmente, las correlaciones fenotípicas y genéticas se calcularon por medio de análisis bivariado, y los efectos fijos fueron aquéllos del análisis univariado.
En el Cuadro 2 se presentan las medias de cuadrados mínimos de los efectos fijos. El sexo de las crías mostró diferencias significativas tanto para PN como para PC (P<0.05). Los machos fueron 5 % más pesados que las hembras. Las crías de parto simple fueron 41.53 % más pesadas al nacimiento y 20.6 % más pesadas al destete que las de parto múltiple (P< 0.05). Se observaron corderos más pesados para ambos parámetros en los partos tercero y cuarto (P< 0.05). En las condiciones ambientales y de manejo de hatos imperantes en los Altos de Chiapas, la raza ovina Chiapas presentó un peso promedio al nacer de 2.48 ± 0.52 kg y de 8.96 ± 2.54 kg promedio al destete.
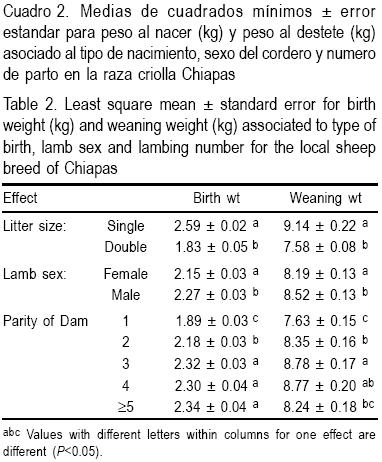
Los parámetros genéticos para PN y PC para los seis modelos se muestran en el Cuadro 3. La estimación más alta para heredabilidad directa de PN se obtuvo en el modelo 1 (0.37 ± 0.09) y las más bajas en los modelos 3 (0.09 ± 0.05) y 4 (0.08 ± 0.05). Las estimaciones de heredabilidad materna fueron similares en los modelos 3 (0.23 ± 0.04) y 6 (0.24 ± 0.09), y más altas que en los modelos 4 (0.20 ± 0.04) y 5 (0.15 ± 0.06). La correlación entre efectos genéticos directos aditivos y genéticos maternos (σam) fue media y negativa. La varianza de efectos maternos permanentes se redujo 0.07 ± 0.05 (modelo 5) cuando no se incluyó el efecto σam.
La estimación más alta de heredabilidad directa para PC se obtuvo en el modelo 1 (0.22 ± 0.07), que disminuyó cuando se introdujeron efectos genéticos maternos y ambientales. La estimación más baja se observó en el modelo 5 (0.07 ± 0.06). También se obtuvo una estimación baja de heredabilidad directa en los modelos 2 (0.08 ± 0.06) y 3 (0.08 ± 0.06). En los modelos 4 (0.15 ± 0.09) y 6 (0.14 ± 0.09) se encontró una estimación de heredabilidad directa que prácticamente duplica la de los modelos 2, 3 y 5. Las estimaciones de heredabilidad materna fueron mayores en los modelos 3 (0.11 ± 0.04) y 4 (0.17 ± 0.07) que las observadas en los modelos 5 (0.04 ± 0.05) y 6 (0.09 ± 0.08). Las estimaciones de varianzas materno ambiental es permanentes disminuyeron cuando se introdujeron efectos genéticos maternos en los modelos 5 y 6 (0.08 ± 0.06). La correlación entre efectos aditivos directos y efectos genéticos maternos para PC fue negativa y fluctuó entre -0.51 ± 0.29 y -0.61 ± 0.34.
Los valores de AIC de los modelos se ordenaron con el objeto de obtener los valores Δi. El AIC para PN muestra al modelo 3 como el mejor (AIC = -847.34), seguido por el modelo 6 (Δ6=0.96) y el modelo 4 (Δ4=1.48). Para PC, el mejor modelo resultó ser el 2 (AIC = 2384.84), seguido por los modelos 5 (Δ5 = 1.36) y 3 (Δ3 = 1.48), como era de esperar, porque esas características son influenciadas por el efecto materno.
Las correlaciones fenotípica y genética aditiva entre PN y PC fueron 0.34 ± 0.03 y 0.81 ± 0.18, respectivamente.
Efectos fijos
Las medias de cuadrados mínimos para PN y PC observadas en este estudio fueron similares a las comunicadas en 1987 cuando se caracterizó este tipo de ganado lanar explotado por los tzotziles en Chiapas(2). Muy probablemente esto es debido a que el ganado no ha sido objeto de selección genética para ninguna característica de crecimiento, y que el sistema de producción permanece prácticamente sin cambios. Esta raza es probablemente la de menor tamaño de las que se explotan en México, siendo sus pesos al destete un 40 % de los de la raza Suffolk (20.7 kg) a los 68 días(13), un 68 % de los de la raza Pelibuey (12.2 kg a los 75 días) y un 90 % de los de las razas Criollas del Centro de México (13.2 kg a los 110 días )(14). En concordancia con estudios realizados en otras razas, se encontraron efectos ambientales significativos para tipo de parto, número de parto de la madre, año de nacimiento y sexo de la cría(4,13,15).
Modelo genético
Las grandes diferencias en valores de pérdida de información del modelo 1 respecto del mejor modelo encontrado para PN (Δ1 = 39.08) y PC (A1 = 8.32) significa que será el menos probable de usar de los seis modelos propuestos(12). Se considera que el mejor modelo para PN es 6, ya que la pérdida de información fue de Δ3=0.96, además de que incluye todos los efectos descritos como de importancia en la literatura(6,7,8). Sin embargo, para PC los modelos 5, 3, 4 y 6 fueron similares al mejor modelo 2, (Δi varía entre 1.36 y 2.38) y muestran evidencia de que estos modelos son posibles de utilizar(12), y por lo tanto el modelo 6 fue tomado en consideración, al igual que para PN(6,7,8).
Peso al nacimiento
Los valores de heredabilidad aditiva directa (h2a) dependen del modelo que se emplee. En el modelo 1 se estimó 0.37 ± 0.06 y 0.15 ± 0.09 en el modelo 6. Estas estimaciones concuerdan con las mencionadas por otros autores en estudios realizados en las razas ovinas locales Menz(6), Cjallonké(7) y Sangsari(8) para peso corporal.
El valor de h2a disminuyó a 0.17, similar al informado por Al-Shorepy(16) y Mandal et al(17), cuando se introdujo el efecto de ambiente materno permanente. La introducción de efecto genético materno y de ambiente materno permanente al modelo afectó negativamente a h2a, h2m y c2 (sus valores cayeron a 0.10, 0.15 y 0.07 , respectivamente). El valor de h2a hallado es similar al mencionado en otros estudios(17,18). El valor de h2m calculado por medio del modelo 6 es similar al publicado por otros autores(17,19,20).
La estimación de c2 se encuentra dentro del rango que se menciona en la literatura (0.01 a 0.20)(17,21,22,23) e indica que la influencia de c2 sobre PN se debe al efecto de útero y de partos múltiples. Aún más, Cuguma et al(24) mencionan que la exclusión de c2 puede ser causa de una sobrestimación de h2m, especialmente en peso al nacer.
Los resultados observados en este estudio para ra,m en la raza Chiapas estuvieron dentro de un rango entre -0.38 y -0.58 para PN dependiendo del modelo empleado. Larsgard y Olsen(25) argumentan que la sensibilidad a la estructura de la información puede explicar la gran variación entre estimaciones, aunque también se sugiere la presencia de cambios debidos a circunstancias de manejo y ambientales(26). Meyer et al(27) señalan que estas estimaciones negativas altas pueden deberse a un problema derivado de una partición incorrecta de la variación materna de la directa.
La correlación entre efectos genéticos directos y genéticos maternos para características de crecimiento se mantuvo en general entre baja y mediana y fue consistentemente negativa, lo que sugiere un probable antagonismo entre los genes aditivos del cordero y los genes maternos que contribuyen a los efectos maternos(28). Otros autores(20, 23, 29, 30, 31) también mencionan correlaciones genéticas maternas negativas, con valores negativos extremos de -0.99 a -0.14.
Peso al destete
La heredabilidad aditiva directa (h2a) para peso al destete fluctuó entre 0.07 y 0.22. Estimaciones similares se publicaron para razas de doble propósito, con valores que varían de 0.02 a 0.19(32).
Cuando se incluyó el efecto materno, la h2a para PC disminuyó a 36 % del valor hallado en el modelo sin efecto materno (0.08). La estimación de h2m en el modelo 3, de 0.11, es similar a la mencionada en otros estudios(8,21).
En el modelo 6, las estimaciones de heredabilidad materna y directa fueron bajas; además, el valor de c2=0.08 se encuentra dentro del rango mencionado en otros trabajos(21,32).
Las estimaciones de la correlación entre efectos genéticos directos y maternos se encuentran dentro del rango de -0.51 a -0.61, parecido a la estimación de -0.51 mencionada por Maniatis y Pollot(33), pero menor a los valores hallados por otros autores(30,34).
Hagger(35) sugiere que la correlación negativa alta entre efectos aditivos directos y maternos pueden deberse a la ausencia de efectos de semental-hato o de semental-año en el modelo como se menciona en Lewis y Beatson(28) y a otras limitantes de la estructura de datos(33).
En general, las diferencias entre las estimaciones de los distintos parámetros dependen del modelo. Resultados parecidos son mencionados por distintos autores, llegando a la conclusión que la heredabilidad directa se sobrestima cuando los efectos maternos no se toman en cuenta(15,16,36). Las correlaciones genéticas y fenotípicas para PN y PC halladas en este trabajo son similares a las mencionadas por otros autores(6,8,32).
Las estimaciones de componentes de la varianza y heredabilidad para efectos genéticos aditivos directos y maternos para PN y PC en la raza ovina Chiapas sugieren la presencia de una variabilidad genética de importancia para estas dos características. Cebido a esto, los programas de selección para mejorarlas son factibles en esta raza. Un modelo que incluya tanto efectos genéticos aditivos directos y maternos como efectos de ambiente maternos permanentes fue considerado como el más adecuado para la evaluación genética de la población de esta raza ovina.
AGRADECIMIENTOS
Este estudio fue financiado por CONACYT (por medio de una beca otorgada a Iván Olivera Vega), UNAM-PAPI IT IN207707-3 y SAGARPA-CONACYT 2004-CO1-111/A1.
LITERATURA CITADA
1. FAO 2004, Cata base FAOSAT. http//www.fao.org [ Links ]
2. Perezgrovas GR, Castro GH. El borrego Chiapas y el sistema tradicional de manejo de ovinos entre las pastoras tzotziles. Arch Zoot 2000;(49):391-403. [ Links ]
3. Perezgrovas GR. Papel de las pastoras Tzotziles en la conservación de la diversidad del ganado lanar de Chiapas. Anim Genet Resour Inform 2009;45:65-69. [ Links ]
4. Rashidi A, Mokhtari MS, Safi Jahanshahi A, Mohammad Abadi MR. Genetic parameter estimates of pre-weaning growth traist in Kermani sheep. Small Rum Res 2008;74:165-171. [ Links ]
5. Ajoy M, Roy R, Rout PK. Cirect and maternal effects for body measurements at birth and weaning in Muzaffarnagari sheep of India. Small Rum Res 2008;75:123-127. [ Links ]
6. Gizaw S, Lemma S, Komen H, van Arendonk JAM. Estimates of genetic parameters and genetic trends for live weight and fleec traits in Menz sheep. Small Rum Res 2007;70:145-153. [ Links ]
7. Bosso NA, Cissé MF, Waaij EH, Fall A, Arendonk JA. Genetic and phenotypic parameters of body weight in West African Cwarf goat and Cjallonké sheep. Small Rum Res 2007;67:271-278. [ Links ]
8. Miraei-Ashtiani SR, Reza SSA, Moradi SM. Variance components and heritabilities for body weight traits in Sangsari sheep, using univariate and multivariate animal models. Small Rum Res 2007;73:109-114. [ Links ]
9. García E. Modificaciones al sistema de clasificación climática de Köppen. In., Series Modificaciones al sistema de clasificación climática de Köppen. México: Instituto de Geografía UNAM. México, CF. 1981. [ Links ]
10. JMP Institute Inc 2002.User's guide. Version 5.0. SAS Inst. Inc.Cary., Nc, USA. [ Links ]
11. Gilmour AR, Geogel BJ, Cullis BR, Thompson R. ASReml. In: Reference manual. Series ASREML. Harpenden, U.K: IACR-Rothamsted Exp Sta.2005. [ Links ]
12. Burnham KP and Anderson, C R. Multimodel inference understanding AIC and BIC in model selection. Sociol Meth Res 2004;2:261-304. [ Links ]
13. Sulaiman Y, Flores-Serrano C, Ortiz HA, Montaldo HH. Evaluación de métodos de corrección para efectos ambientales para peso al destete en corderos Suffolk. Vet Méx 2009;40:219-229. [ Links ]
14. Hernández CPC, Ruiz BRF. Curvas de crecimiento posdestete en ovinos criollos y Suffolk. [tesis licenciatura]. Edo. de México, Mexico: Universidad de Chapingo; 2002. [ Links ]
15. Matika, O, van Wyka JB, Erasmusa GJ, Bakerb RL. Genetic parameter estimates in Sabi sheep. Livest Prod Sci 2003;79:17-28. [ Links ]
16. Al-Shorepy SA. Estimates of genetic parameters for direct and maternal effects on birth weight of local sheep in United Arab Emirates. Small Rum Res 2001;39:219-224. [ Links ]
17. Mandal A, Neser FWC, Rout PK, Roy R, Notter CR. Estimation of direct and maternal (co)variance components for pre-weaning growth traits in Muzaffarnagari sheep. Livest Prod Sci 2006;99:79-89. [ Links ]
18. Ozcan M, Ekiz B, Yilmaz A, Ceyhan A. Genetic parameter estimates for lamb growth traits and greasy fleece weight at first shearing in Turkish Merino sheep. Small Rum Res 2005;56:215-222. [ Links ]
19. Cloete SWP, Scholtz JA, Guilmour AR, Olivier JJ. Genetic and environmental effects on lambing and neonatal behaviour of Cormer and SA Mutton Merino lambs. Livest Prod Sci 2002;78:183-193. [ Links ]
20. Cloete SWP, Greeff JC, Lewer R. Environmental and genetic aspects of survival liveweight in Western Australian Merino sheep. S Afr J Anim Sci 2001;31:123-130. [ Links ]
21. Hanford KJ, Van Vleck LC, Snowder GC. Estimates of genetic parameters and genetic trend for reproduction, weight, and wool characteristics of Polypay sheep. Livest Prod Sci 2006;102:72-82. [ Links ]
22. Maxa J, Norberg E, Berg P, Pedersen J. Genetic parameters for growth traits and litter size in Canish Texel, Shropshire, Oxford Cown and Suffolk. Small Rum Res 2007;68,:312-317. [ Links ]
23. Neser FWC, Erasmus GL, van Wyk JB. Genetic parameter estimates for pre-weaning weight traits in Corper sheep. Small Rum Res 2001;40:197-202. [ Links ]
24. Cuguma G, Schoeman SJ, Cloete SWP. Genetic parameter estimates of early growth traits in the Tygerhoek Merino flock. S Afr J Anim Sci 2002;32:66-75. [ Links ]
25. Larsgard AG, Olesen I. Genetic parameters for direct and maternal effects on weights and ultrasonic muscle and fat depth of lamb. Livest Prod Sci 1998;55:273-278. [ Links ]
26. Robinson OW. The role of maternal effects in animal breeding. V maternal effects in swinw J Anim Sci 1972;35:1303-1315. [ Links ]
27. Meyer K, Carrick MJ, Conelly BJP. Genetic parameters for growth traits of Australian Beef Cattle from multibreed selection. J Anim Sci 1993;71:2614-2622. [ Links ]
28. Lewis RM, Beatson PR. Choosing maternal effect models to estimate (co)variances for live and fleece weight in New Zealand Coopworth sheep. Livest Prod Sci 1999;58:137-150. [ Links ]
29. Ligda C, Gabriilidis G, Papadopoulos T, Georgoudis A. Investigation of direct and maternal genetic effects on birth and weaning weight of Chios lambs. Livest Prod Sci 2000;67:75-80. [ Links ]
30. María GA, Boldman KG, Van Vleck LC. Estimates of variances due to direct and maternal effects for growth traits of Romanov sheep. J Anim Sci 1993;71:845-849. [ Links ]
31. Maxa J,Norberg E, Berg P, Milerski M. Genetics parameters for body weight, longissimus muscle depth and fat depth for Suffolk sheep in the Czech Republic. Small Rum Res 2007;72:87-91. [ Links ]
32. Safari E, Fogarty NM, Gilmur AR. A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep. Livest Prod Sci 2005;92:271-289. [ Links ]
33. Maniatis N, Pollott GE. The impact of data structure on genetic (co)variance components of early growth in sheep, estimated using an animal model with maternal effects. J Anim Sci 2003;81:101-108. [ Links ]
34. El Fadili M, Michaux C, Cetilleuxb J, Leroy PL. Genetic parameters for growth traits of the Moroccan Timahdit breed of sheep. Small Rum Res 2000;37:203-208. [ Links ]
35. Hagger C. Litter. Permanent environmental, ram-flock, and genetic effects on early weight dain of lambs. J Anim Sci 1998;76:452-457. [ Links ]
36. Al-Shorepy SA, Alhadrami GA, Abdulwahab K. Genetic and phenotypic parameters for early growth traits in Emirati goat. Small Rum Res 2002;45:217-223. [ Links ]

