











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La mutagénesis inducida es una técnica por la cual se han generado 671 variedades mutantes de ornamentales, en su mayoría corresponden a Chysanthemum spp. (285 variedades), Rosa spp. (67 variedades), Dahlia spp. (35 variedades), Alstroemeria spp. (35 variedades) y Streptocarpus spp. (30 variedades) y fueron generadas principalmente en China, Holanda, Japón, India y Alemania (FAO/IAEA, 2020). Esta técnica ha sido muy utilizada debido a que la mayoría de las plantas de ornato se propagan vegetativamente, lo que permite la fácil reproducción de mutantes (Rangaiah, 2006; Yamaguchi, 2018) y es justificable cuando no se tiene variabilidad natural; por lo tanto, es necesario inducirla para tener un abanico amplio de individuos con características deseables.
Las ventajas de este proceso son: la posibilidad de generar variabilidad; la rapidez con la que se puede obtener un individuo con caracteres deseables, es decir, reduce el tiempo en comparación con el mejoramiento tradicional (De la Cruz, 2010) y al no ser considerados organismos genéticamente modificados (OGMs) este método está libre de restricciones regulatorias (Parry et al., 2009). Las desventajas son: las mutaciones ocurren de manera impredecible en todo el genoma, son puntuales y es difícil obtener un mutante con múltiples características deseadas, se requiere un gran número de semillas o partes vegetativas para que se logre identificar el efecto de un agente mutagénico; resulta difícil identificar un pequeño número de individuos con fenotipos novedosos dentro de una población grande y muchas mutaciones no tienen un efecto detectable en las plantas (De la Cruz, 2010).
Las mutaciones inducidas son las provocadas por agentes exógenos denominados mutágenos, los cuales pueden ser químicos o físicos; entre los químicos se encuentran el etilmetanosulfonato (EMS), sulfato de dimetilo y sulfato de dietilo, metilnitrosourea (MNU) y etilnitrosourea (ENU) (Oladosu et al., 2016) y entre los agentes físicos encontramos a los rayos x, rayos gamma (Jain, 2005, Yamaguchi et al., 2008), luz ultravioleta (Ahloowalia y Maluszynski, 2001) e iones de carbono (Wu et al., 2009).
En plantas ornamentales, los mutágenos más utilizados en el mejoramiento son los físicos (Jain, 2005, Wu et al., 2009; Yamaguchi, 2018; Yamaguchi et al., 2018; Hernández-Muñoz et al., 2019), ya que pueden estimular la germinación, acelerar el desarrollo de las plantas, favorecer la resistencia a diferentes tipos de estrés, principalmente enfermedades y provocar cambios en el color, forma y tamaño de flores (Maluszynski et al., 2001; Ahloowalia y Maluszynski, 2001, Chopra, 2005; Datta y Teixeira, 2006). En especies donde no se cuenta con información previa, lo primero que se debe hacer es un estudio de radiosensibilidad, que consiste en evaluar la sensibilidad que tienen las células, tejidos, semillas y estructuras vegetativas al agente mutagénico para determinar la dosis letal media. La radiosensibilidad varía de acuerdo con la especie, el nivel de ploidía, grado de diferenciación del tejido y contenido de humedad (De la Cruz, 2010).
En plantas propagadas por semillas las poblaciones mutagenizadas son generadas por exposición de semillas (M0) al mutágeno, la obtención de plantas (M1) autógamas o alógamas que darán lugar a semillas (M2). En virtud de que las mutaciones son usualmente un evento recesivo, se recomienda realizar la selección en la generación M2, en la cual se hacen evidentes las mutaciones en condición homocigótica recesiva (Chikelu, 2013). Las plantas M2 continúan su desarrollo y se obtienen semillas M3, en esta etapa, la población todavía está segregando y no todas las plantas M3 llevarán las mutaciones identificadas en la M2.
La población mutagenizada puede fijar una mutación hasta la generación M6, generando material casi homocigoto, aunque en el proceso se pierde hasta la mitad de las mutaciones presentes en la M1 (Parry et al., 2009; Ukai y Nakagawa, 2011).
En México se han realizado experimentos donde se ha determinado que la radiación gamma 60Co estimula la germinación en semillas de Laelia autumnalis (Lex.) Lindl. (Hernández-Muñoz et al., 2017). En nardo (Polianthes tuberosa L.) se ha evaluado su efecto en tubérculos y plantas in vitro (Estrada-Basaldua et al., 2011), en crisantemo (Dendranthema grandiflorum (Ramat.) Kitam.) se produjeron mutantes con genotipos de lento desarrollo, enanos y con moteado en hojas (Castillo-Martínez et al., 2015), mientras que al irradiar bulbos de Tigridia pavonia (L. f.) DC. se logró modificar el color de la flor (Díaz-López et al., 2003).
En girasol (Helianthus annuus L.) y jamaica (Hibiscus sabdariffa L.) la radiación gamma tuvo efecto sobre la germinación de semillas (Díaz-López et al., 2017), similar comportamiento en uchuva (Physalis peruviana L.) para germinación de semilla y altura de planta, así como componentes de rendimiento (Antúnez et al., 2020a, b) y por último, en nochebuena (Euphorbia pulcherrima Willd. ex Klotzsch) tuvo efecto sobre la emergencia, características de semillas y parámetros morfológicos en plantas adultas (Canul-Ku et al., 2012), habiéndose registrado en 2016, la variedad Juanita de nochebuena, como flor de maceta obtenida por mutagénesis (Canul-Ku et al., 2019).
Euphorbia fulgens es una especie que se comercializa como ornamental en Europa y otros países, principalmente como flor de corte, aunque también se oferta planta en maceta. Su nombre común es scarlet plume (pluma escarlata) y es considerada una planta de día corto. Se comercializan diversos cultivares con distintos colores de inflorescencias: naranja, rosa, rojo, blanco, salmón, durazno y amarillo; mientras que el color de las hojas puede ser verde, rojo oscuro o morado.
En nuestro país, de donde es originaria, en las poblaciones silvestres sólo se encuentran inflorescencias con tonalidades que van del naranja al rojo (Pérez-Nicolás et al., 2021). Por lo que es posible, que dichas variedades se originaron a través de mutagénesis inducida. Así, la variedad ‘Albora’ se desarrolló en Holanda por irradiación de esquejes con rayos X (40Gy), fue aprobada oficialmente en 1976 y su principal atributo mejorado es el color de las inflorescencias (FAO/IAEA, 2020). Considerando lo antes expuesto, el objetivo de la presente investigación fue evaluar el efecto de radiación gamma de 60Co en semillas y detectar cambios en caracteres morfológicos de plantas adultas de E. fulgens en las generaciones M1 y M2 para obtener mutantes con diferente color de apéndices petalíferos.
Materiales y métodos
El experimento se realizó en un invernadero de cristal ubicado en la Universidad Autónoma Chapingo, la cual se localiza a 19° 20’ latitud norte y 98° 53’ longitud oeste, a 2 240 m de altitud. La temperatura y humedad relativa se registró con un data logger HOBO® U12-012 (Onset Computer Corporation, Massachusetts, EUA), la temperatura media varió entre16 y 24 °C y la humedad relativa entre 38 y 68%. En abril del año 2018 se colectaron frutos de Euphorbia fulgens en dos sitios del estado de Oaxaca, el sitio 1 se localiza en San Jerónimo Coatlán y el sitio 2 en Santiago Jamiltepec.
Los frutos se colocaron en bolsas de papel de estraza hasta que se secaron y liberaron sus semillas. Las semillas se depositaron en bolsas de glassine y se mantuvieron a temperatura ambiente, se eligieron 200 semillas de cada sitio las cuales se distribuyeron en 10 bolsas conteniendo 20 semillas cada una (M0). El 04 de mayo del mismo año, las semillas se irradiaron en un irradiador Transelektro (LGI-01, fabricado en Hungría) con fuente de 60Co, localizado en el Instituto Nacional de Investigaciones Nucleares (ININ). Se establecieron 9 dosis de radiación con gammas de 60Co (50, 100, 150, 200, 250, 300, 450, 600, 750 Gy) más el testigo sin irradiar, teniéndose 40 semillas por tratamiento.
Ese mismo día se sembraron las semillas irradiadas (M1) bajo un diseño completamente al azar en charolas de unicel de 200 cavidades utilizando como sustrato turba/perlita (50:50 v/v). Las plantas M1 en etapa de primer par de hojas verdaderas opuestas (finales de mayo, principios de junio) se trasplantaron a macetas de 11 x 11 x 15 cm, utilizando como sustrato turba/agrolita/vermiculita (40:30:30 v/v/v). En esta etapa, se agregó al agua de riego fertilizante Ultrasol® inicial N15-P30-K15+M. E. AQTEX 0.5 g L-1 cada ocho días. A finales de julio, agosto y septiembre se aplicó Ultrasol® Multipropósito N18-P18-K18 + M. E. AQTEX 1 g L-1 cada ocho días.
A inicios de octubre del mismo año, las plantas se cambiaron a bolsas negras de polietileno de 25 x 25 cm que contenían sustrato de tierra de hoja de encino/fibra de coco/vermicomposta/tierra negra/corteza de pino (30:30:20:10:10). Se realizaron aplicaciones preventivas con fungicidas para evitar la aparición de hongos con Benomilo, 1.5 g L-1; para evitar la aparición de mosca negra se usó Bifendrina 1.5 ml L-1 alternando con Cipermetrina, adicionalmente se colocaron trampas cromáticas de color azul y amarillo. No obstante, la plaga que se presentó fue la araña roja (Tetranychus urticae), por lo que se controló con Abamectina 1.5 ml L-1 y Bacillus thuringiensis 0.5 g L-1. El control de malezas se realizó de manera manual.
En la generación M1, se registró el porcentaje de emergencia y el número de plantas que sobrevivieron hasta la etapa adulta con dos a tres frutos desarrollados. En esta etapa fenológica se tuvo una población M1 de 137 plantas y se registraron 27 variables en todas ellas; se utilizó un flexómetro para medir la altura de planta (cm) y mediante una regla se midieron las siguientes variables: longitud de peciolo (cm), longitud de lámina (cm) y ancho de la lámina (cm).
Estos datos fueron usados para calcular la relación entre el ancho de la lámina y la longitud de la lámina (cm) y la relación entre la longitud de peciolo y la longitud de la lámina (cm). También se contabilizó el número de ramas con ciatios, el número de cimas y número de ciatios hermafroditas, se midió la longitud de entrenudos (cm), la longitud de la inflorescencia (cm), longitud de la cima más larga (cm), longitud de entrenudos en la inflorescencia (cm), longitud de pedúnculo (cm), largo de involucro (cm), largo del apéndice petalífero (cm), ancho del apéndice petalífero (cm), longitud del pedicelo (cm), longitud del ovario (cm), longitud de los estilos (cm), longitud de los estambres (cm), longitud del fruto (cm) y el diámetro del fruto (cm), el cual se midió con un vernier Pretul®.
Por último, se registraron las siguientes variables con base en la guía para variedades de E. fulgens de la Unión Internacional para la protección de nuevas variedades de plantas (UPOV, 1988): el color de los apéndices petalíferos, el color de las láminas del tercio superior de la parte con cimas (rojo o verde), la intensidad del color rojo en las mismas láminas, así como el color de las láminas en el tercio inferior de la parte con cimas (rojo o verde) para la cual se utilizaron las cartas de color de The Royal Horticultural Society (2001).
Posteriormente, a finales de diciembre 2018 y a principios de enero 2019 y dado que esta especie es alógama (Pérez-Nicolás, 2020), se realizaron cruzamientos aleatorios para garantizar la producción de semillas, entre individuos obtenidos de la generación M1; después en los meses de marzo y abril de 2019 se cosecharon los frutos, se sembraron las semillas el día 12 de mayo de 2019 y se siguió su desarrollo hasta la etapa adulta (generación M2). Cabe señalar, que tanto la siembra como el desarrollo de las plantas se llevó a cabo bajo las mismas condiciones que el ciclo anterior (generación M1).
En la generación M2, se registró el porcentaje de emergencia y el número de plantas que sobrevivieron hasta la etapa adulta con dos a tres frutos desarrollados. En esta etapa fenológica se realizó la caracterización de 48 plantas adultas durante la última semana del mes de noviembre, momento en que la mayoría de las plantas se encontraban floreciendo.
Con base en los resultados de la generación M1, se determinó medir las 17 variables, en todos los individuos de la generación M2: altura de planta con un flexómetro; diámetro del tallo con un Vernier Pretul® y con una regla la longitud de peciolo, longitud de lámina, ancho de lámina, longitud de entrenudos, longitud de inflorescencia, longitud de la cima más larga, longitud de entrenudos en la inflorescencia, longitud de pedúnculo (cm), largo de involucro (cm), largo del apéndice petalífero (cm), ancho del apéndice petalífero (cm). Se contabilizaron las hojas antes de la inflorescencia, número de cimas, número de ciatios. Por último, se tomó el color de apéndices petalíferos con las cartas de color de The Royal Horticultural Society (2001).
En ambas generaciones, las variables vegetativas se tomaron en el segundo tercio de la planta, de tres hojas y tres entrenudos y las estructuras reproductivas del segundo tercio de la inflorescencia, de tres cimas y de tres ciatios.
Para evaluar la generación M1, se realizó un análisis de varianza (Anova) entre los diferentes tratamientos (0, 50, 100, 200 y 300 Gy) y cuando se detectaron diferencias estadísticas entre tratamientos se aplicó la prueba de comparación de medias de Tukey (p≤ 0.05) utilizando el paquete estadístico Infostat versión 2020 (Di Rienzo et al. 2008). Para evaluar las características fenotípicas en las generaciones M1 y M2 se consideraron 13 variables y se hizo un análisis estadístico descriptivo a través de tablas de frecuencias y gráficos en Excel.
Resultados y discusión
En la generación M1, en ambos sitios de colecta, el tratamiento testigo (sin radiación) presentó el mayor porcentaje de emergencia, el cual disminuyó conforme se incrementó la dosis de radiación. Se calculó un modelo de regresión lineal simple por sitio de colecta entre la dosis de radiación y el porcentaje de emergencia, el cual explicó 86% del comportamiento observado. En el sitio 1 por cada Gray de radiación 60Co la emergencia disminuyó en 0.08%, mientras que en el sitio 2 disminuyó en 0.03% (Figura 1).
El porcentaje de emergencia para el testigo en el sitio 1 fue de 80%, mientras que en el sitio 2 fue del 55%. En ambos sitios, a partir de 250 Gy murieron entre 55% y 56% de las plantas, antes de llegar a la etapa adulta. En las dosis más altas (450, 600 y 750 Gy) emergieron de una a tres plántulas, la mayoría presentaron hipocótilos y cotiledones deformes y sin clorofila. En las dosis de 750 Gy sobrevivió un individuo de cada sitio, por lo que no fue posible calcular la dosis letal media, sin embargo, considerando las curvas de regresión para ambos sitios se puede estimar la LD50 en 275.41 Gy, para el sitio 1, y en 401 Gy, para el sitio 2.
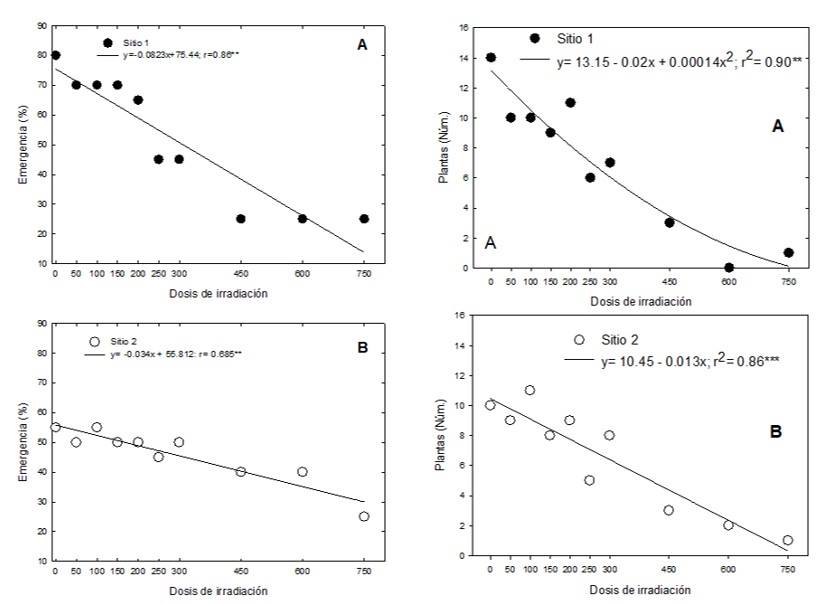
Figura 1 Porcentaje de emergencia de plántulas y sobrevivencia de plantas de E. fulgens, provenientes de semillas irradiadas con rayos gamma 60 Co, de dos sitios de colecta en México. A) sitio 1; y B) sitio 2.
Las plantas que sobrevivieron a las dosis más altas presentaron malformaciones en hojas e inflorescencias (Figura 1). Los resultados obtenidos en el presente estudio coinciden con Díaz-López et al. (2017) que en jamaica y girasol al incrementar la dosis de radiación obtuvieron porcentajes de germinación menores en comparación con el testigo, al igual Canul-Ku et al. (2012) en nochebuena obtuvieron porcentajes de emergencia menores al aumentar la dosis de radiación.
La sobrevivencia de plantas mostrada a altas dosis de radiación en este trabajo se puede explicar debido a que las semillas que se sometieron a irradiación provienen de plantas silvestres, las cuales tienen mayor plasticidad para adaptarse a cambios repentinos en su hábitat en comparación con las especies cultivadas.
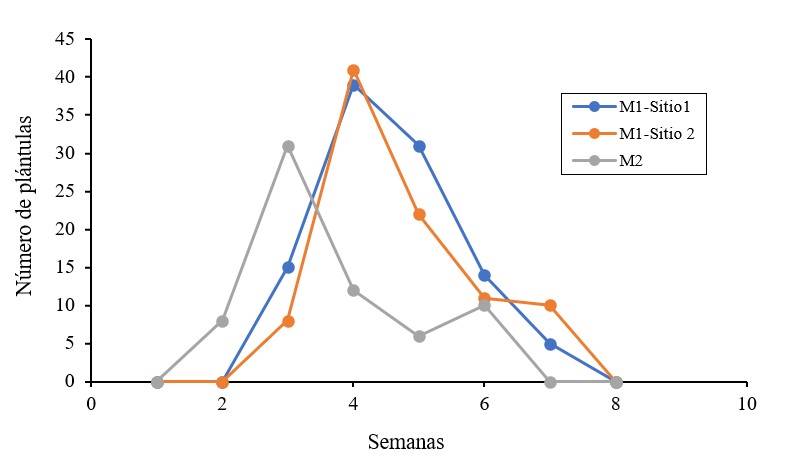
En la generación M1 la emergencia para ambos sitios de colecta se presentó 15 días después de la siembra, en un total de siete semanas, mientras que en la generación M2, la emergencia se presentó siete días después de la siembra, en un total de seis semanas (Figura 2), por lo que, hubo una disminución en el tiempo de emergencia bajo las mismas condiciones de cultivo.
El análisis de varianza no mostró diferencias significativas entre tratamientos para la mayoría de las variables consideradas tanto de estructuras vegetativas como reproductivas. En el número de ramas con flores, el tratamiento 300 Gy fue significativamente diferente al tratamiento 200 Gy, mientras que el resto de los tratamientos no produjeron diferencias significativas. Para la variable color de los apéndices petalíferos, el tratamiento 300 Gy fue significativamente diferente al testigo y a los tratamientos 50 Gy y 100 Gy (Cuadro 1). Sin embargo, en trabajos con semillas irradiadas de nochebuenas silvestres ha sido posible detectar efectos inmediatos, encontrando diferencias significativas en siete características, entre estas, la altura de la planta y diámetro del ciatio (Canul-Ku et al., 2012).
Cuadro 1 Variables morfométricas en plantas de Euphorbia fulgens (M1) a diferentes dosis de radiación gamma 60 Co.
| Variable | 0 Gy | 50 Gy | 100 Gy | 150 Gy | 200 Gy | 300 Gy |
| Altura | 101.88a | 102.97a | 92.05a | 98.34a | 100.31a | 98.54a |
| Longitud de peciolo | 3.1a | 2.73a | 2.87a | 3.1a | 3.21a | 3.05a |
| Longitud de la lámina | 7.86a | 7.92a | 7.49a | 7.86a | 8.12a | 7.65a |
| Ancho de la lámina | 2.36a | 2.3a | 2.29a | 2.35a | 2.41a | 2.27a |
| Ancho lámina/longitud lámina | 0.3a | 0.3a | 0.31a | 0.3a | 0.3a | 0.3a |
| Longitud peciolo/longitud lamina | 0.4a | 0.35a | 0.38a | 0.39a | 0.39a | 0.4a |
| Longitud de entrenudos | 1.57a | 1.56a | 1.76a | 1.63a | 1.57a | 1.54a |
| Número de ramas con inflorescencias | 1ab | 1ab | 1ab | 0.94ab | 0.89b | 1.21a |
| Longitud de la inflorescencia | 27.5a | 20.99a | 20.43a | 20.06a | 21.08a | 27.27a |
| Número de cimas | 16.2a | 12.6a | 12.6a | 12.06a | 12.67a | 16.56a |
| Longitud de la cima más larga | 5.43a | 5.26a | 4.79a | 4.78a | 4.43a | 5.2a |
| Número de ciatios | 72.85a | 58.67a | 67.25a | 53.06a | 62.83a | 7.64a |
| Longitud de entrenudos en la inflorescencia | 1.09a | 0.97a | 1.07a | 0.96a | 1.02a | 1.2a |
| Longitud de pedúnculo | 1.12a | 1.22a | 1.13a | 1.14a | 0.96a | 1.19a |
| Largo del involucro | 0.31a | 0.31a | 0.31a | 0.28a | 0.27a | 0.3a |
| Largo apéndice petalífero | 0.53a | 0.53a | 0.54a | 0.51a | 0.48a | 0.56a |
| Ancho apéndice petalífero | 0.49a | 0.49a | 0.52a | 0.46a | 0.44a | 0.51a |
| Longitud pedicelo | 0.85a | 0.83a | 0.81a | 0.79a | 0.72a | 0.79a |
| Longitud de estilos | 0.30a | 0.29a | 0.3a | 0.28a | 0.27a | 0.3a |
| Longitud del ovario | 0.11a | 0.17a | 0.12a | 0.1a | 0.09a | 0.1a |
| Longitud del estambre | 0.65a | 0.62a | 0.63a | 0.59a | 0.58a | 0.66a |
| Longitud de fruto | 0.47a | 0.23a | 0.46a | 0.36a | 0.37a | 0.39a |
| Diámetro de fruto | 0.44a | 0.21a | 0.42a | 0.34a | 0.34a | 0.36a |
| Número de frutos | 2.6a | 2a | 2.9a | 1.56a | 2.28a | 3.43a |
| Número de semillas | 4.75a | 2.44a | 4.40a | 2.13a | 4.28a | 4.64a |
| Color de apéndices petalíferos | 1.55b | 1.33b | 1.20b | 1.69ab | 1.78ab | 2.57a |
| Intensidad de color de la lámina de la hoja en el tercio superior de la inflorescencia | 1.3a | 1.11a | 1.5a | 1.25a | 1.23a | 1.21a |
abc= dosis con letra distinta en el sentido de la fila son estadísticamente diferentes (p< 0.05).
En la generación M1, de 400 semillas establecidas que corresponden al sitio uno y sitio dos sobrevivieron el 34.25%, de estas plantas se obtuvieron 296 semillas M1, de las cuales sobrevivió el 16.21% que corresponden a la generación M2, ya que la mayoría no fueron viables. En ambas generaciones se obtuvieron características semejantes en estructuras vegetativas (Figura 3). La variable altura en los progenitores (M1) osciló entre 35 y 165 cm, el intervalo con mayor frecuencia fue de 100 a 116 cm, mientras que en la descendencia (M2) osciló de los 50 a los 168 cm y la mayoría se ubicó en el intervalo de 101 a 134 cm. La longitud de peciolo en los progenitores M1 osciló de 1 a 6 cm, el intervalo con mayor número de plantas fue 2.2 a 2.7 cm, en la progenie, M2, de 1.7 a 4.6 cm, la mayoría de las plantas se ubicó en el intervalo de 2.5 a 2.9 cm (Figura 3).
La longitud de lámina en la generación M1 estuvo entre los 4.2 a 10.4 cm, el intervalo con mayor frecuencia fue de 7.3 a 8 cm y en la generación M2, de 6.1 a 9.7 cm, donde el intervalo con mayor número de plantas fue de 7.3 a 7.9 cm. El ancho de la lámina 0.7 a 3.9 cm, el intervalo con mayor frecuencia fue de 2.6 a 2.9 cm y en la descendencia osciló entre 1 y 4 cm, el intervalo con mayor número de plantas fue de 2 a 2.4 cm. La longitud de entrenudos de 0.6 a 3.1 cm, el intervalo con mayor número de plantas fue de 1.3 a 1.6 cm y en la descendencia de 1.1 a 3.7 cm y el intervalo con mayor número de plantas fue de 1.9 a 2.2 cm (Figura 3).
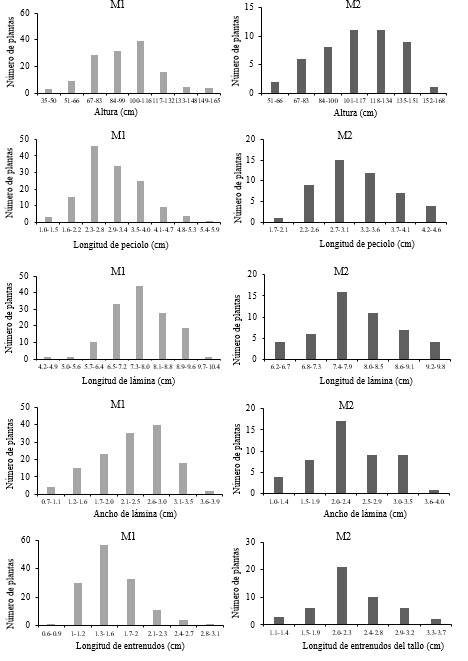
En variables de estructuras reproductivas encontramos variación entre los progenitores y descendencia, destacando menor número de cimas en progenitores, pero mayor número de ciatios respecto a la descendencia (Figura 4). La longitud de inflorescencia, en la generación M1, osciló de 3 a 67 cm encontrándose la mayoría en el intervalo de 19 a 26 cm y en la generación M2 de 9.5 a 32.5 cm el intervalo con mayor número de plantas es de 19.4 a 22.6 cm. La longitud de la cima más larga en parentales fue de 1.9 a 11.6 cm y la mayoría se encontró en el intervalo de 3.1 a 4.2 cm y en su descendencia fue de 2.9 a 7.2 cm y la mayoría se encontró en 3.6 a 4.2 cm. En la generación M1 se presentó de 3 hasta 36 cimas, pero la mayoría se ubicó en el intervalo de 8 a 12 cimas, en la generación M2, de 6 a 30 cimas y la mayoría de 14 a 17 cimas (Figura 4).
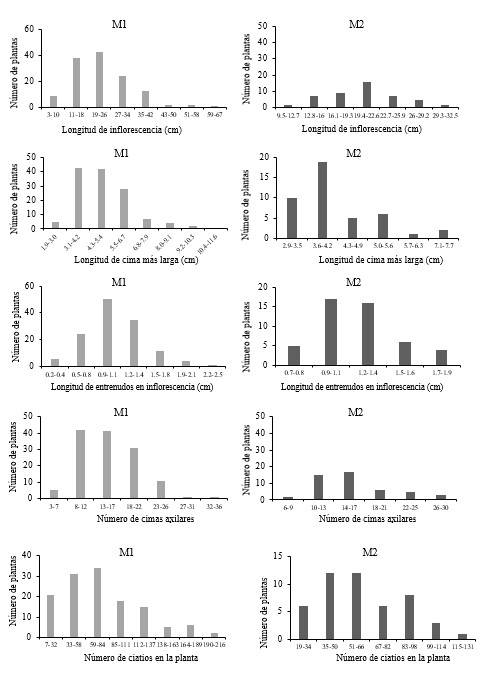
La longitud de entrenudos en la inflorescencia en los progenitores estuvo entre los 0.2 a 2.5 cm y la mayoría se ubicó en el intervalo de 9.9 a 1.1 cm y en la descendencia fue de 0.8 a 1.9 cm y la mayoría se encontró en 9.9 a 1.3 cm. El número de ciatios contabilizados en cada planta de la generación M1 fue de 7 a 216, la mayoría fue de 59 a 84 ciatios. En la generación M2 fue de 19 a 131 y la mayoría presentó de 35 a 66 ciatios. La longitud del apéndice petalífero en la generación M1 fue de 0.4 a 0.7 pero la mayoría fue de 0.5 y en su descendencia fue de 0.3 a 0.8 y la mayoría fue de 0.4 a 0.5 (Figura 4). El ancho del apéndice petalífero en progenitores fue de 0.3 a 0.7 y la mayoría fue de 0.5, mientras que en su descendencia fue de 0.3 a 0.7 y la mayoría en el intervalo de 0.4 a 0.5 cm.
Respecto a la variable color en los progenitores se registraron 8 colores diferentes siendo el más común Orange Red Group N30 A, mientras que en la descendencia se registraron 13 colores diferentes siendo el más común el mismo color que los progenitores (Figura 5 y 6).
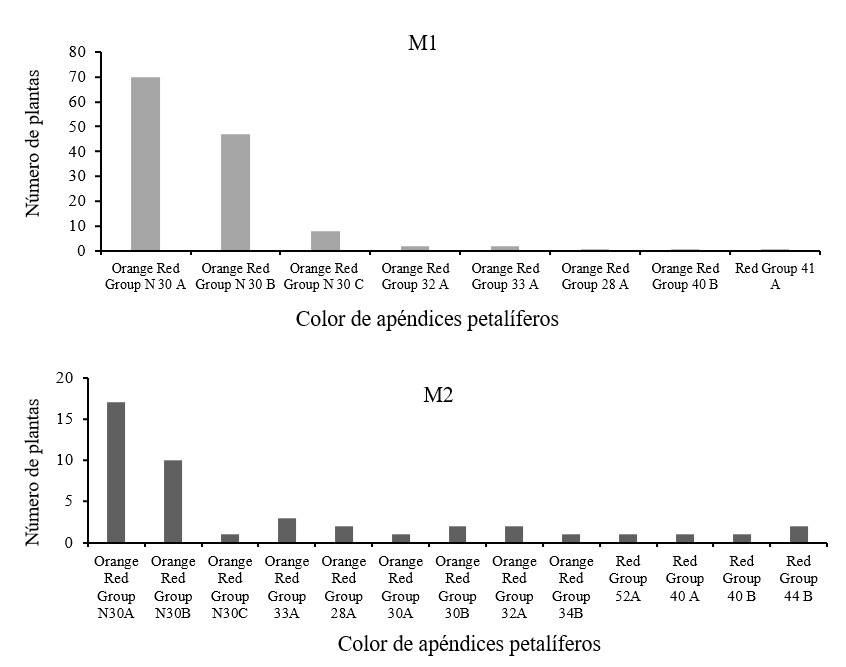

En un estudio previo donde se evaluaron características morfológicas de individuos de poblaciones silvestres y de un primer ciclo de cultivo de plantas obtenidas de semillas sin irradiar, la variable color no presentó diferencias significativas (Pérez-Nicolás et al., 2021). A pesar de que usando una dosis de radiación de 300 Gy fue posible obtener mutantes con diferente color de apéndices petalíferos, lo cual era uno de los principales objetivos del presente estudio, la sobrevivencia de las plantas fue baja y se presentan malformaciones debido a que el daño propiciado por la radiación al material genético es mayor, por lo que es conveniente realizar experimentos con 100 Gy o menos, ya que estas dosis que no afectaron la supervivencia de las plantas.
Además, se recomienda usar mayor cantidad de semillas en dosis que propiciaron los cambios de coloración, considerando que la tasa de mutación inducida es de alrededor de 1x10-4 (De la Cruz, 2010). Sin embargo, por ser una especie microendémica de distribución restringida lo más recomendable sería irradiar esquejes a dosis bajas, los cuales podrían obtenerse de los materiales generados en este trabajo y con ello no tener que extraer material de poblaciones silvestres y provocar alteración.
Conclusiones
En las dosis de radiación utilizadas en este trabajo, 50-750Gy, no se alcanzó la letalidad total. Con base en las curvas de radiosensibilidad obtenidas, se estima la LD50 en 275 Gy, para el sitio 1 y 401 Gy para el sitio 2. Las dosis aplicadas tuvieron efecto en el porcentaje de emergencia y sobrevivencia de las plantas, pero no hubo efecto en la mayoría de las variables morfométricas evaluadas, sólo en el número de ramas con flores y el color de los apéndices petalíferos. La radiación tuvo un efecto sobre la variación del color de inflorescencias desde la generación M1 y éste fue mayor en la generación M2. Con relación a las generaciones de mutagénesis, las características fenotípicas en los progenitores (M1) fueron similares a las que presentó su descendencia (M2) bajo las mismas condiciones de cultivo, destacando que en la generación M2 se registró mayor variación de color.

