










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.12 no.2 Texcoco feb./mar. 2021 Epub 25-Abr-2022
https://doi.org/10.29312/remexca.v12i2.2847
Articles
Impact of cover, ecotype and endomycorrhizae on morphology and quality of piquin chili
1Universidad Autónoma Agraria Antonio Narro. Buenavista, Saltillo, Coahuila, México. CP. 25315. (rosalindamendoza@hotmail.com; robledo3031@gmail.com; miguel-cbg@hotmail.com; vimarc59@yahoo.com.mx).
2Tecnológico Nacional de México-Campus Instituto Tecnológico de Torreón. Antigua Carretera Torreón-San Pedro km 7.5, Torreón, Coahuila, México. CP. 27170. (reguillen@outlook.com).
3Doctorado en Ciencias en Agricultura Protegida-Universidad Autónoma Agraria Antonio Narro, Saltillo, Coahuila México. CP. 25315.
The research was carried out with piquin chili ecotypes from the states of Coahuila, Nuevo León and Zacatecas. Piquin chili plants were evaluated in its second year of production. Two environments were used: a) macro tunnels of white mesh, red, blue, black raschel type with 30% shade; and b) open field with 100% light transmission. 50 spores of a conglomerate of arbuscular mycorrhizal fungi (Glomus mosseae, Rhizophagus intraradices, Sclerocystis coremioides and Gigaspora albida) were inoculated directly to the radical system. A factorial arrangement 5 x 6 x 2 (cover, ecotypes and mycorrhizae) was used and its distribution corresponded to a random block design with four repetitions. The yield and some morphological characteristics of the plant were evaluated such as plant height (AP), stem diameter (DT), root length (LR), fresh plant weight (PFP), dry weight of the plant (PSP), fresh root weight (PFR), dry root weight (PSR), yield per plant (RPP). The results indicate that the white mesh favored the morphological characteristics of the plant with agronomics with 320% AP, 322.7% DT, 235.8% LR, 8 times PFP, 8.5 times PSP, 327.2% PFR, 5 times PSR, 6.8 times PTR, compared to plants developed in open field. In addition, the quality of piquin chili provides conditions for the development of endomycorrhizae (spores and percentage of colonization). Blue mesh with the lowest photosynthetically active radiation (RFA) positively influenced agronomic, quality variables and inoculation (number of spores and percentage of colonization). The ecotype that influenced SST, Vit C, phenols and NE was SNL and RTZ in agronomic variables and % colonization. The inoculation with the mycorrhizae consortium improved the agronomic characteristics and quality of the piquin chili fruit.
Keywords: Capsicum annuum; quality; shadow mesh; solar radiation
La investigación se realizó con ecotipos de chile piquín de Coahuila, Nuevo León y Zacatecas. Se evaluaron plantas de chile piquín del segundo año de producción. Se utilizaron dos ambientes: a) macro túneles de malla blanca, roja azul, negra tipo raschel con 30% de sombreo; y b) campo abierto con 100% de transmisión de luz. Se inocularon 50 esporas de un conglomerado de hongos micorrízicos arbusculares (Glomus mosseae, Rhizophagus intraradices, Sclerocystis coremioides y Gigaspora albida), directamente al sistema radical. Se utilizó un arreglo factorial 5 x 6 x 2 (cubiertas, ecotipos y micorrizas) y su distribución fue un diseño bloques al azar con cuatro repeticiones. Se evaluó el rendimiento y algunos caracteres morfológicos como altura de planta (AP), diámetro de tallo (DT), longitud de raíz (LR), peso fresco de planta (PFP), peso fresco de planta (PSP), peso fresco de raíz (PFR), peso seco de raíz (PSR), rendimiento por planta (RPP). Los resultados indicaron que la malla blanca favoreció los caracteres morfológicos de la planta con 320% en AP, 322.7% en DT, 235.8% la LR, 8 veces el PFP, 8.5 veces el PSP, 327.2% el PFR, 5 veces el PSR, 6.8 veces el RPP, en comparación a las plantas desarrolladas en campo abierto. Además, la calidad de chile piquín proporciona condiciones para el desarrollo de endomicorrizas (esporas y porcentaje de colonización). La malla azul con la menor radiación fotosintéticamente activa (RFA) influyó positivamente en las variables agronómicas, de calidad y la inoculación (número de esporas y porcentaje de colonización). El ecotipo que influyó en los SST, VitC, fenoles y NE fue el de SNL y el RTZ en variables agronómicas y % colonización. La inoculación con el consorcio de micorrizas mejoró los caracteres agronómicos y calidad del fruto de chile piquín.
Palabras clave: Capsicum annuum; calidad; malla sombra; radiación solar
Introduction
Piquin chili (Capsicum annuum L.) is a phytogenetic resource widely distributed in Mexico, given its nature the wild form of this species is the predominant one, its fruit is usually obtained by harvesting in wild populations, but it is threatened by its genetic diversity (Pagán et al., 2010). Piquin chili has not been completely domesticated since it is observed low germination and morphological and genetic variability (García-Federico et al., 2010; Hernández-Verdugo et al., 2015).
Piquin chili yields are affected by environmental conditions, soil moisture and fertilization (Rodríguez et al., 2005). In relationto environmental factors it has been shown that direct sunlight causes more compact plants, whereas with 80% shade the plants grow quickly and become larger. Capsicum plants’ response to different irradiance conditions may vary according to cultivar. The photoperiod, quality and quantity of sunlight coincide, directly, in the photosynthesis of plants and other phenotypic and functional characteristics (Peixoto et al., 2014).
The phenotypic responses of plants to varied light conditions have not been adequately used to modify the morpho-physiological characteristics of crops and obtain desired yield and quality (Kelly et al., 2015). The use of colored meshes is an alternative to avoid excessive radiation as with red mesh, which provides 42.6% total solar radiation (350 to 1 050 nm) and blue mesh 36% more blue light (400 to 500 nm) than black mesh (Ayala-Tafoya et al., 2015). These differences in radiation can cause differential response in photosynthesis and photomorphogenesis that produce effects on stem growth, foliar expansion, chloroplast development, chlorophyll synthesis and secondary metabolites. In cucumber the use of red and blue meshes increased yield 48.1 and 46.1% compared to the witness (Ayala-Tafoya et al., 2015).
On the other hand, the use of biofertilizers in the cultivation of piquin chili has shown positive effects on plant height, root length, increased dry biomass in saline and non-salty soils (Rueda et al., 2010). It has also been shown that mycorrhizal fungi have an affinity in different Capsicum species such as annuum, bacatum, chinense, frutescens and pubescens which confirms their positive effect (Cardona et al., 2008).
However, the application of commercial inoculums does not assure the positive effect on plant-mycorrhizal fungus synergism, due to the indiscriminate use of agrochemicals in traditional production systems (Koyama et al., 2017; Caruso et al., 2018). In addition, in other crops such as tomato, mycorrhizae (Glomus mossae and Glomus cubense) have been used in liquid form (10 and 20 spores) and solid form (20 and 40 spores) and have produced beneficial results for the plant (Mujica et al., 2012).
The objective of this research work was to evaluate the effect of inoculation with native endomycorrhizae of three locations in the northeast of the country under photoselective cover and open field in the morphology and quality of piquin chili.
Materials and methods
The study was carried out, in the Horticulture Department of the Antonio Narro Agrarian Autonomous University, Saltillo, Coahuila, Mexico, located at 25° 22’ north latitude and 101° 22’ west longitude, at a height of 1 580 meters above sea level. Six ecotypes of piquin chili were collected, called: MZC= Múzquiz, Coahuila; SAC= San Alberto, Coahuila; LNL= Linares, Nuevo León; SNL= Santiago, Nuevo León; PTZ= Tepetatilla Bridge, Zacatecas and RTZ= Tuxpan River, Zacatecas.
Seed planting was carried out in germinating boxes to which it was applied GA3 (500 ppm) to speed up the germination process, after one month the seedlings were transplanted into polyethylene bags with a capacity of 10 L and placed at a distance of 40 cm between plants and 1 m between grooves, as a substratum was used sphagnum peat moss (Pro Mix®) and perlite (Hortiperl®) in a proportion 2:1 (v/v). In the experiment these chili ecotypes were used in their second year of production, different luminosity environments (cover color) were used: a) macro tunnels of white mesh (MA), red (MR), blue (MA), black (MN) raschel type with 30% shade, with a hole size of 6 x 8 mm; each tunnel 4 m wide, 6 m long and 2.30 m high; and b) open field with 100% light transmission.
The crop was fertilized with nutrient solution, 25% in seedling, 50% in vegetative development, 75% in flowering and 100% in fructification. At the beginning of this evaluation, phosphorus input was reduced to 25%, with the intention that endomycorrhizae presented synergy with plant roots. The water supply was made from 0.5 to 2.5 L plant-1 day-1, with a fertigation system per stake.
The plants were inoculated after transplantation (50 spores) with a conglomerate of arbuscular mycorrhizal fungi (Glomus mosseae, Rhizophagus intraradices, Sclerocystis coremioides and Gigaspora albida), directly to the radical system, which were identified by comparative morphology (Sánchez-Sánchez et al., 2018).
These factors were evaluated by a factorial arrangement 5 x 6 x 2 (roof color:5, ecotypes:6 and mycorrhiza:2). The experimental design used corresponded to a random block with four repetitions. Microclimatic variables such as environmental temperature and relative humidity were recorded with a digital thermo hygrometer (Taylor® model 1452). Photosynthetically active radiation (RFA) was recorded with a Quantum portable sensor (Apogee® model SM-700). Measurements were made daily between 07:00 and 19:00 h, in the center of each deck, in clear sky conditions.
Morphological characteristics were evaluated in three plants by repetition and treatment, which included: plant height (AP), made with tape measure, stem diameter (DT), with a digital vernier (Digital Caliper®), 8 cuts were made, to obtain the average fruit yield per plant, were weighed with an electronic scale Rhino model Babol-100G with capacity of 100 g and resolution of 0.01 g. The dry weight of the plant (PSP) and the fresh weight of the plant (PFP) and of root (PFR) were determined on an OHAUS scale model CS-5000 with a capacity of 5 kg. To obtain the dry weight of the plant (PSP) and of root (PSR), the samples were placed on brown paper and subjected to 65°C for 48 hours on a Yamato drying stove model DX-602 and subsequently weighed on the aforementioned scale.
The number of leaves (NH), the number of fruits per plant (NFPP) was estimated by counting in each unit, evaluations were made in all experimental units. In quality variables three samples were evaluated per treatment and repetition for, total soluble solids (SST) with a digital refractometer HANNA 96-801, in which a drop of fruit pulp was placed in the cell of the apparatus, obtaining the content expressed in Brix. The content of ascorbic acid (Vitamin C), in fruits was determined by the AOAC methodology (2000).
The total phenol content (FT) was determined according to the methodology reported by Kim et al. (2006), with some modifications described below, 2 g of fresh piquin chili fruit was weighed and placed in 20 mL of 80% methanol, for 12 h at 4 °C, after the time was centrifuged at 12 000 rpm for 5 min, an aliquot of 200 μl was taken from the supernatant mixed with 150 μl of the agent Folin Ciocaltaeu 2 N, 2 ml of Na2CO3 to 2% were added, leaving it incubated for 25 min and finally the absorbance was read at 735 nm in spectrophotometer (Bio-145025 BIOMATE 5 Thermo elctron Corporation), the calibration curve was made with gallic acid.
The quantification of capsaicin (CAPs) was determined in fruits with physiological maturity, by the method described by Bennet and Kirby (1965), by a spectrophotometer (Bio-145025 Biomate-5 Thermo Electron Corporation) at a wavelength of 286 nm, in which capsaicin is in its organic phase. For the determination of the concentration a calibration curve was constructed of this antioxidant (Sigma, Co) in a range 0.5 to 1.5 mg ml-1. In quality variables the evaluations were tripled for each treatment.
The number of spores was quantified by triplicate for each treatment and repetition in 100 g of soil with the method of wet sieving and decanting (Genderman and Nicolson, 1963), root cleaning and staining was performed with the Phillips and Hayman method (1970) and colonization (McGonigle, 1990) which consists of washing the roots with running water, cut them and place them in 25 ml falcon tubes, cover them with 10% KOH by 24 h at room temperature, then rinsed with plenty of running water, covered with H2O2, for 5 min, then rinsed with running water, covered roots with 10% HCl for 10 min, then HCl was decanted and without rinsing the roots, the 0.05% trypan blue solution was added by 24 h at room temperature, after time the dye was removed with the help of a sieve and they were placed in lactoglycerol, finally segments of 1 cm root were cut and deposited in a slide, were observed in an optical microscope (Axio Scope A1, Carl Zeiss, Microscopy GmbH, Gotting, Germany). The results obtained were analyzed using a variance analysis and the comparison of means by the Tukey test (p≤ 0.05), using the SAS version 9.0 statistical program.
Results and discussion
In Figure 1 the RFA was measured noting that maximum radiation is between 13 and 15 h, and in CA there is 95% higher compared to the MR who captures radiation between 350 to 1 050 nm, MB and MN are similar and the lowest absorption was carried out with MA whose radiation is between 400 and 500 nm (Ayala-Tafoya et al., 2015). In flower crops the black mesh reduces RFA between 55 and 60% depending on the season and red mesh from 41 to 51% (Arthurs et al., 2013).
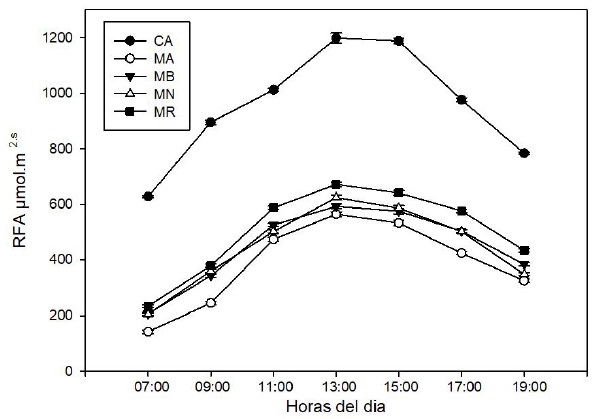
Figure 1 Photosynthetically active radiation in open field and colored meshes in piquin chili plants.
Table 1 shows the temperature values, the month with the highest record is may in open field decreasing between 1 and 2 degrees until September. It is also observed that MR and CA have similar temperature and MA decreases from 2 to 5 degrees in relation to CA. The Table 1 shows that HR increased each month until it reaches its maximum in September, also in CA the largest HR is observed only swing in September with MA and MB.
Table 1 Monthly average values of climate variables monitored in each environmental condition under open field and color meshes.
Cover |
Months evaluated |
Meses evaluados |
||||
May |
June |
July |
August |
September |
||
CA |
T (°C) |
26.98 |
25.56 |
23.34 |
22.86 |
20.74 |
HR (%) |
34.54 |
43.1 |
57.32 |
62.38 |
69.36 |
|
MA |
T (°C) |
21.45 |
22.34 |
21.55 |
19.33 |
18.44 |
HR (%) |
34.22 |
42.11 |
55.32 |
63.44 |
70.44 |
|
MB |
T (°C) |
22.13 |
23.11 |
22.11 |
20.55 |
18.98 |
HR (%) |
34.58 |
44.55 |
55.67 |
61.78 |
65.89 |
|
MN |
T (°C) |
22.33 |
22.41 |
22.15 |
20.77 |
20.11 |
HR (%) |
33.77 |
42.33 |
55.65 |
60.43 |
66.51 |
|
MR |
T (°C) |
23.78 |
24.51 |
23.11 |
21.52 |
20.17 |
HR (%) |
33.11 |
41.22 |
56.73 |
60.22 |
68.55 |
|
T°= temperature; HR= relative humidity.
Table 2 shows that MB positively favors agronomic variables evaluated with 320% in AP, 322.7% in DT, 235.8% in LR, 8 times the PFP, 8.5 times the PSP, 327.2% the PFR, 5 times the PSR, 6.8 times the RPP and 4.8 times the NFPP compared to CA which was the least favorable environment, perhaps under normal growing conditions of piquin chili plants these are under the shade of a tree or shrub which prevent direct radiation. The MB reflects 70% radiation allowing favorable conditions for the growth of the plant, the pearl-colored mesh increased its transmission from 700 nm which favors cucumber photosynthesis (Ayala-Tafoya et al., 2015) and consequently the growth of the plant unlike what was reported by Shahak et al. (2008) indicating that production was increased in three crops of pepper chili with 16% pearl mesh and 32% with red mesh compared to black mesh.
Table 2 Agronomic variables of the different factors evaluated in piquin chili plants.
Cover |
AP (cm) |
DT (mm) |
LR (cm) |
PFP (g) |
PSP (g) |
PFR (g) |
PSR (g) |
RPP (g) |
|
22.94e |
4.1f |
15.1e |
24.81e |
11.34e |
10.6e |
4.36e |
5.5f |
36.52f |
|
44.62d |
6.96e |
28.44d |
43.04d |
19.56e |
18.13d |
7.21d |
16.81d |
80.1e |
|
53.69b |
9.76b |
29.79c |
93.63b |
36.59b |
21.14c |
11.74c |
18.08c |
113.08b |
|
47.25cd |
9.39c |
36.12a |
64.13c |
29.31c |
24.31b |
14.54b |
22.8b |
105.17c |
|
73.42a |
13.23a |
35.6ab |
200.52a |
96.71a |
34.68a |
21.32a |
37.42a |
177.08a |
|
Ecotype | |||||||||
33.75e |
6.9e |
28.22e |
44.85d |
20.89e |
19.45f |
10.75d |
7.09f |
59.5f |
|
42.35d |
8.46c |
28.28d |
76.44c |
34.71cd |
24.37b |
14.53a |
26.08b |
133.75b |
|
44.6cd |
8.73c |
27.8f |
75.63c |
32.01d |
20.81c |
11.24c |
10.57d |
70.9e |
|
71.25a |
9.8a |
29.13c |
115.54a |
48.73a |
20.7d |
10.35f |
50.48a |
169.31a |
|
52.46b |
9.29b |
30.68a |
87.44b |
43.87b |
25.15a |
13.43b |
11.93c |
94.19c |
|
46.08c |
7.68d |
29.73b |
74.94c |
37.16c |
20.15e |
10.7e |
9.27e |
78.1d |
|
Consortium | |||||||||
38.28b |
7.31b |
25.25b |
58.57b |
27.15b |
18.03 |
9.05b |
14.13b |
88.24b |
|
58.56a |
9.65a |
32.69a |
99.71a |
45.3a |
25.52a |
14.62a |
24.33a |
113.68a |
|
CV (%) |
9.53 |
5.82 |
8.54 |
13.78 |
14.59 |
9.91 |
14.86 |
5.89 |
5.81 |
AP= plant height; DT= stem diameter; LR= root length; PFP= fresh weight of plant; PSP= dry weight of plant; PFR= fresh weight of root; PSR= dry weight root; RPP= yield per plant; NFP= number of fruits per plant; RTZ= Tuxpan River, Zacatecas; PTZ= Tepetatilla Bridge, Zacatecas; LNL= Linares, Nuevo León; MZC= Múzquiz, Coahuila; SNL= Santiago, Nuevo León; SAC= San Alberto, Coahuila; MB= white mesh; MA= blue mesh; MR= red mesh; MN= black mesh; CA= open field; SM= without mycorrhiza; CM= with mycorrhiza. Means with the same letter within each column in each factor do not differ statistically (Tukey, p≤ 0.05). CV=coefficient of variation. S*= significance; **= highly significant (p≤ 0.001).
The ecotype that favored the aforementioned variables was RTZ, only LNL increased the LR and MZC the PSR (Table 2). However, it has been reported that each morphotype presents different to the environmental conditions as is the case with morphotypes from Oaxaca whose phenotypic characteristics are very different (Castellón Martínez et al., 2014), to those from Nuevo León, Coahuila and Zacatecas used in this study. In Table 2, it is observed that inoculation with native mycorrhizae had a favorable response with the increase of 53% in AP, 32% in DT, 70.24% in PFP, 66.9% in PSP, 72.2% in R and 28.8% in NFPP. This matches Cardona et al. (2008) who found that arbuscular mycorrhizae colonize the roots of the genus Capsicum, for this the morphological characteristics of piquin chili were improved.
In Table 3, quality variables are presented, it is observed that for SST they are increased in fruits with 32% with MR and 37.6% in MB, vitamin C (42.9%) and total phenols also with (44.31%) MB and Capsaicin with MA 18.42%, in addition to the itching represented by Scoville units (SHU) in the MA, in the same table the ecotypes that present statistical differences for SST are the MZC and SNL ecotypes. For Vitamin C, SAC and SNL, total phenols SNL, for capsaicin and itching (US) PTZ.
Table 3 Quality variables of factors evaluated in piquin chili plants.
Cover |
SST (°Brix) |
VitC (mg 100 g-1) |
FT (µg EAG g-1) |
CAPs (mg g-1) |
U Scoville SHU |
6.41c |
60.85f |
34.77e |
2.66d |
42 560d |
|
6.69c |
66.3e |
37.09d |
3.07b |
49 120b |
|
8.46a |
84.29b |
42.42b |
2.95c |
47 200c |
|
7.52b |
71.24c |
43.05b |
3.15a |
50 400a |
|
8.82a |
86.96a |
50.18a |
3.09ab |
49 440ab |
|
Ecotype | |||||
6.71c |
82.98a |
34.91d |
2.98bc |
47 680bc |
|
8.57a |
68d |
37.47c |
2.93c |
46 880c |
|
8.31a |
79.95b |
37.54c |
3.09a |
49 440a |
|
7.4b |
78.38c |
46.28b |
3.02b |
48 320b |
|
7.41b |
82.97a |
35.82d |
2.93c |
46 880c |
|
8.39a |
45.21e |
55.37a |
3.02b |
48 320b |
|
Consortium | |||||
8.31a |
72.27b |
40.08b |
2.92b |
46 720b |
|
7.29b |
73.56a |
42.38a |
3.07a |
49 120a |
|
CV (%) |
5.09 |
0.85 |
5.79 |
3.98 |
3.98 |
SST= total soluble solids; Vit C= vitamin C; FT= total phenols; CAPs= capsaicin; RTZ= Tuxpan River, Zacatecas; PTZ= Tepetatilla Bridge, Zacatecas; LNL= Linares, Nuevo León; MZC= Múzquiz, Coahuila; SNL= Santiago, Nuevo León; SAC= San Alberto, Coahuila; MB=white mesh; MA= blue mesh; MR= red mesh; MN= black mesh; CA= open field; SM= without mycorrhiza; CM= with mycorrhiza. Means with the same letter within each column in each factor do not differ statistically (Tukey, p≤ 0.05). CV= coefficient of variation. S*= significance; **= highly significant (p≤ 0.001).
The CM application produced better results in vitamin C (1%), total phenols (3.87%) and capsaicin (5.13%) increasing the antioxidants of the fruit and itching, only in SST was obtained the 27% increase SM. In relation to quality a study was carried out with different chili morphtypes such as piquin and solterito which produced higher phenol and flavonoid content in addition to capsaicin (Wei et al., 2013), although mycorrhizae were not added, the tendency of morphotypes is to produce higher antioxidant content.
In Table 4 shows that when analyzing the cover separately the MB is the one that increases by 96% the number of spores and 101.4% of colonization. It is also shown that the SNL ecotype increased NE and RTZ the % colonization, in relation to inoculation with native mycorrhizae favored NE and % colonization in piquin chili plants. This is consistent with a study about papaya where it is inoculated with Glomus sp. finding an increase in the percentage of colonization (Quiñones-Aguilar et al., 2014).
Table 4 Microbiological variables of factors evaluated in piquin chili plants.
Cover |
Col (%) |
|
23.33e |
16.17c |
|
36.67b |
23.67b |
|
31.15cd |
25.67b |
|
35.83bc |
25.92b |
|
45.73a |
32.57a |
|
Ecotype | ||
29.27c |
25.58ab |
|
34.48ab |
25.42ab |
|
34.79ab |
23.42b |
|
35ab |
27.58a |
|
31.56bc |
23.5b |
|
37.19a |
25.08b |
|
Consortium | ||
5.38b |
2.31b |
|
62.05a |
47.89a |
|
CV (%) |
25.75 |
23.62 |
NE= number of spores; % Col= colonization percentage; RTZ= Tuxpan River, Zacatecas; PTZ= Tepetatilla Bridge, Zacatecas; LNL= Linares, Nuevo León; MZC= Múzquiz, Coahuila; SNL= Santiago, Nuevo León; SAC= San Alberto, Coahuila; MB= white mesh; MA= blue mesh; MR= red mesh; MN= black mesh; CA= open field; SM= without mycorrhiza; CM= with mycorrhiza. Means with the same letter within each column in each factor do not differ statistically (Tukey, p≤ 0.05).
Principal component analysis (ACP), performed for the cover type and variables evaluated Figure 3, showed that MB positively influenced in the RPP, PFR, AP, PSR, DT and NFPP variables, contrary to what was obtained with MN and CA, while the MA influenced the content of Caps and LR, in turn the MR promoted an increase in the variables Vit C, SST, PFP and PSP, so the use of different cover promote diverse results in morphology and quality in piquin chili plants; consistent with what was reported in cucumber (Ayala-Tafoya et al., 2015) when colored meshes were used.

Figure 3 Principal components of analyzed variables and cover type. AP= plant height; DT= stem diameter; LR= root length; PFP= fresh weight of plant; PSP= dry weight of plant; PFR= fresh weight of root; PSR= dry weight of root; RPP= yield per plant; NFP= number of fruits per plant; SST= total soluble solids; Vit C= vitamin C; FT= total phenols; CAPs= capsaicin; NE= number of spores; % Col= colonization percentage; MB= white mesh; MA= blue mesh; IN= greenhouse; MR= red mesh; MN= black mesh; CA= open field.
The ACP for ecotypes and variables evaluated Figura 4, revealed a dispersed behavior of the ecotypes evaluated; however, it emphasizes that the RTZ ecotype favors the variables AP, PSP, RPP, NFPP, DT; while SAC and PTZ ecotypes are not favored in these variables, while LNL ecotype favored the increase in PFR and PSR and MZC and SNL ecotypes influenced the quality variables, Vit C, Caps and SST, demonstrating that the ecotype plays a primary role in the variables that were evaluated. This is consistent with Wei et al. (2013) about different chili morphotypes where it finds differences in the content of capsaicin and phenols.
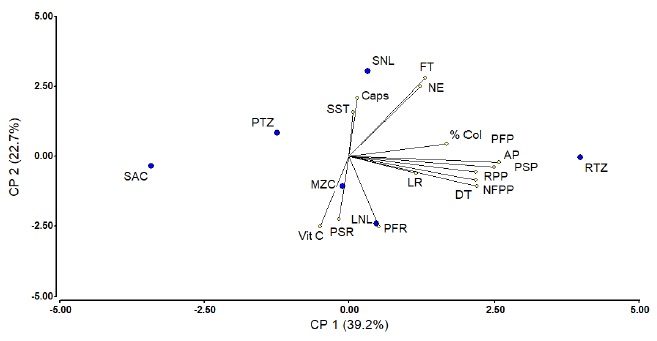
Figure 4 Principal components of the analyzed variables and the ecotype. AP= plant height; DT= stem diameter; LR= root length; PFP= fresh weight of plant; PSP= dry weight of plant; PFR= fresh weight of root; PSR= dry weight of root; RPP= yield per plant; NFPP= number of fruits per plant; SST= total soluble solids; Vit C= vitamin C; FT= total phenols; CAPs= capsaicin; NE= number of spores; % Col= colonization percentage; RTZ= Tuxpan River, Zacatecas; PTZ= Tepetatilla Bridge, Zacatecas; LNL= Linares, Nuevo León; MZC= Múzquiz, Coahuila; SNL= Santiago, Nuevo León; SAC= San Alberto, Coahuila.
The ACP with the application of mycorrhizae (CM) and the absence of these (SM) with the variables studied in Figure 5, demonstrates that there was a strong relationship in most variables evaluated when they were inoculated with mycorrhizae (Glomus mosseae, Rhizophagus intraradices, Sclerocystis coremioides and Gigaspora albid), with the exception of Vit C and the SST variable that was increased when the mycorrhizae were not applied. Similar results were obtained in tomato when was inoculated with Glomus mossae and G. cubense with 20 and 40 spores by increasing yield (Mújica, 2012).
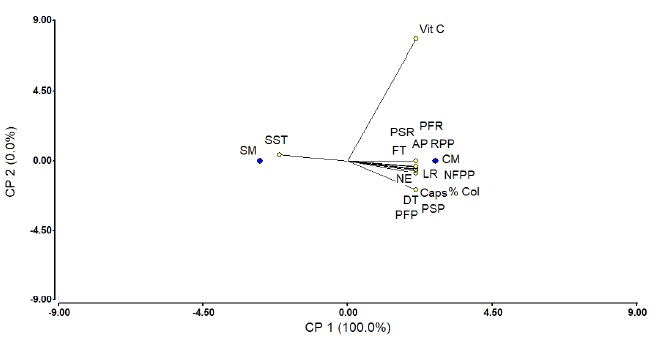
Figure 5 Principal components of the analyzed variables and Consortium. AP= plant height; D= stem diameter; LR= root length; PFP= fresh weight of plant; PSP= dry weight plant; PFR= fresh weight of root; PSR= dry weight of root; RPP= yield per plant; NFP= number of fruits per plant; SST= total soluble solids; Vit C= vitamin C; FT= total phenols; CAPs= capsaicin; NE= number of spores; % Col= colonization percentage; SM= without mycorrhizae; CM= with mycorrhizae.
Conclusions
Blue mesh with the lowest RFA had a positively impacted on the agronomic, quality and response variables to the number of spores and percentage of colonization. The SNL ecotype influenced quality variables such as SST, Vit C, phenols and NE, and RTZ on agronomic variables and % colonization. The inoculation with the mycorrhizae consortium produced favorable changes in agronomic characteristics and quality of the piquin chili fruit.
Literatura citada
AOAC. 2000. Association of Official Analytical Chemists. Official Methods, Assoc. Off. Anal. Chem. Int. (AOAC), Arlington, VA, USA. 2 000 p. [ Links ]
Arthurs, P. S.; Stamps, R. H and Giglia, F. F. 2013. Environmental modification inside photoselective Shadehouses. Hortscience. 48(8):975-979. [ Links ]
Ayala-Tafoya, F.; Yáñez-Juárez, Z. M. G.; Partida-Ruvalcaba, L.; Ruiz-Espinosa, F. H.; Campos-García H.; Vásquez-Martínez, O.; Velázquez-Alcaraz, T. de J. y Díaz-Valdés. 2015. Producción de pepino en ambientes diferenciados por mallas de sombreo fotoselectivo. Información Técnica Económica Agraria. 111(1):3-17. [ Links ]
Bennet, D. J. and Kirby, G. W. 1965. Colorimetry of total phenolics with phosphomolibdic-phpsphotungstic acid reagents. Ame. J. Enology and Viticulture. 16(3):144-158. [ Links ]
Cardona, G.; Peña, C. y Arcos, A. 2008. Ocurrencia de hongos formadores de micorriza arbuscular asociados a ají (Capsicum sp.) en la Amazonia colombiana. Agronomía Colombiana. 26(3):459-470. [ Links ]
Caruso, C.; Maucieri, C.; Barco, A.; Barbera, A. C. and Borin, M. 2018. Effects of mycorrhizal inoculation and digestate fertilisation on triticale biomass production using fungicide-coated seeds. Irish J. Agric. Food Res. 57(1):42-51. [ Links ]
Castellón-Martínez, E.; Carrillo-Rodríguez, J. C; Chávez-Servia, J. L. y Vera-Guzmán, A. M. 2014. Variación fenotípica de morfotipos de chile (Capsicum annuum L.) nativo de Oaxaca, México. Phyton. Inter. J. Exp. Bot. 83(2): 225-236. [ Links ]
García-Federico, A. S.; Montes-Hernández, J. A.; Rangel-Lucio, E.; García-Moya. y Mendoza-Elos, M. 2010. Respuesta fisiológica de la semilla de chile piquín [Capsicum annuum var. glabriusculum (Dunal) Heiser & Pickersgill] al ácido giberélico e hidrotermia. Rev. Mex. Cienc. Agríc. 1(2):203-216. [ Links ]
Gerdemann, J. W. and Nicolson, T. H. 1963. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Transactions of the British Mycological Society. 46(2):235-244. [ Links ]
Hernández-Verdugo, S.; González-Sánchez, R. A.; Porras, F.; Parra-Terraza, S.; Valdez-Ortiz, A.; Pacheco-Olvera, A. and López-España, R. G. 2015. Phenotypic plasticity of populations of wild chile (Capsicum annuum var. glabriusculum) in response to light availability. Bot. Sci. 93(2):231-240. [ Links ]
Kelly, J. W. G.; Landhäusser, S. M. and Chow, P. S. 2015. The impact of light quality and quantity on root-to-shoot ratio and root carbon reserves in aspen seedling stock. New Forests. 46(4):527-545. [ Links ]
Kim, H. J.; Chen, F.; Wang, X. and Rajapakse, N. C. 2006. Effect of ethyl jasmonato on secundary metabolites of sweet basil (Ocimum basilicum L.) J. Agric. Food Chem. 54(6):2327-2332. [ Links ]
Koyama, A.; Pietrangelo, O.; Sanderson, L. and Antunes, P. M. 2017. An empirical investigation of the possibility of adaptability of arbuscular mycorrhizal fungi to new hosts. Mycorrhiza. 27(6):553-563. [ Links ]
McGonigle, T. P; Miller, M. H.; Evans, D. G.; Fairchild, G. L. and Swan, J. A. 1990. A new method which gives an objective measure of colonization of roots by vesiculararbuscular mycorrhizal fungi. New Phytol. 115(3):495-501. [ Links ]
Mujica, P. Y. 2012. Inoculación de hongos micorrízicos arbusculares (HMA) por dos vías diferentes en el cultivo del tomate (Solanum lycopersicum L). Cultivos Tropicales. 33(4):71-76. [ Links ]
Pagán, I.; Betancourt, M.; Miguel, J.; Piñero, D.; Fraile, A. and García-Arenal, F. 2010. Genomic and biological characterization of chiltepín yellow mosaic virus, a new tymovirus infecting Capsicum annuum var. aviculare in Mexico. Archives of Virology. 155(5):675-684. [ Links ]
Peixoto, B. V.; Pereira, C. C. M. A. e Ferreira, R. R. 2014. Emergência e crescimento inicial de Tabebuia heptaphylla (Vell.) Toledo en ambientes contrastantes de luz. Revista Árvore. 38(3):523-531. [ Links ]
Phillips, J. M. and Hayman, D. S. 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society. 55(1):158-161. [ Links ]
Quiñones-Aguilar, E. E.; López-Pérez, L; Hernández-Acosta, E; Ferrera-Cerrato, R y Rincón-Enríquez, G. 2014. Simbiosis micorrízica arbuscular y fuentes de materia orgánica en el crecimiento de Carica papaya L. Interciencia. 39(3):198-204. [ Links ]
Rodríguez, L. A.; Sánchez, R. D. L. C. and Silva, M. M. S. 2005. Effect of sunlight regimes on growth and yield of piquin pepper (Capsicum annuum L. var. aviculare). Rev. Chapingo Ser, Hortic. 11(2):357-359. [ Links ]
Rueda, P. E. O.; Murillo, A. B.; Castellanos, C. T. J. L. G. H.; Tarazón, H. M. A.; Moreno, M. S. and Gerlach, B. L. E. 2010. Effects of plant growth promoting bacteria and mycorrhizal on Capsicum annuum L. aviculare ([Dierbach] D'Arcy ans Eshbaugh) germination under stressing abiotic conditions. Plant Physiol. Biochem. 48(8):724-730. [ Links ]
Sánchez-Sánchez, A.; Salcedo-Martínez, S.; Mendoza-Villarreal, R.; Pinedo-Espinoza, J. and Moreno-Limón, S. 2018. Aislamiento e identificación de micorrizas arbúsculares (MA) asociadas a la rizósfera del chile piquín (Capsicum annuum var. aviculare L.). Investigación y Desarrollo en Ciencia y Tecnología de Alimentos. 3(4):86-91. [ Links ]
Shahak, Y.; Ratner, K.; Giller, Y. E.; Zur, N.; Or, E.; Gussakovsky, E. E. and Greenblat-Avron, Y. 2006. Improving solar energy utilization, productivity and fruit quality in orchards and vineyards by photoselective netting. In: XXVII International Horticultural Congress-IHC2006: International Symposium on Enhancing Economic and Environmental. 772(1):65-72. [ Links ]
Wei, Z. F.; Luo, M.; Zhao, C. J.; Li, C. Y.; Gu, C. B.; Wang, W.; Zu, Y. G.; Efferth, T. and Fu, Y. J. 2013. UV-Induced changes of active components and antioxidant activity in postharvest pigeon pea [Cajanus cajan (L.) Millsp.] leaves. J. Agric. Food Chem. 61(6):1165-1171. [ Links ]
Received: January 01, 2021; Accepted: February 01, 2021
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
