










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.8 no.3 Texcoco abr./may. 2017
https://doi.org/10.29312/remexca.v8i3.26
Articles
Germination of cariopsids classified by size and diaspores of four pastures for semi-arid rain-fed
1 Campus Montecillo-Colegio de Postgraduados. Carretera México-Texcoco, km 34.5. Montecillo, Texcoco, Estado de México, México. CP. 56230. Tel. 01(595) 95201200, ext. 1712 y 1714. (queroadrian@colpos.mx, perpdgo@colpos.mx, helasamy@colpos.mx, garciag@colpos.mx).
2Universidad Tecnológica de Tulancingo. Camino a Ahuehuetitla núm. 301, Col. Las Presas, Tulancingo, Hidalgo, México. CP. 43642. Tel. 01(775) 7558210, 01(800) 8491888. Fax. 01 (775) 7552760.
3Posgrado de Agroindustrias, Universidad Autónoma Chapingo. Carretera México-Texcoco, km 38.5. Texcoco, Estado de México, México. CP. 56230. Tel. (595) 9521500. (patylanda@gmail.com).
4Universidad Autónoma Metropolitana. Calzada del Hueso 1 100, Col. Villa Quietud, Del. Coyoacán, CDMX. CP. 04960. (sramirezs@correo.xoc.uam.mx).
In Poaceae, the commercial seed or propagule consists of the spikelet or diaspora: caryopsis (Cs) + accessory bracts: usually glumes, lemurs, paleas and sometimes species specific, modified branches, which have an effect on germination, and influence the management required and therefore the biological quality of the seed. The removal of accessory bracts with or without Cs classification by size is an undocumented alternative in the establishment of rain-fed praires. The objective of this paper was to evaluate germination by sowing every 30 days, Cs classified from pasture, two native and two introduced, obtained from commercial diaspores at the beginning of the experiment (Cs base -CsB) or every 60 days (Cs control-CsT), for 15 months; similarly, to evaluate the emergence of seedlings from diaspores. The propagules (diasporas or Cs) were evaluated from February 2010 to May 2011, in laboratory. Native and introduced species included Bouteloua gracilis, B. curtipendula, Cenchrus ciliaris and Chloris gayana, respectively. From diaspores, accessory bracts were removed and the resulting Cs were separated with sieves into large (CsG), medium (CsM) and small (CsCh). They were sown based on viable pure seed, the different sizes of Cs, on wet paper, as diaspores at 1 cm depth, in peat moss. The behavior of the germination was analyzed by logistic regression with the Logistic routine of SAS. In the evaluated species, larger Cs size was associated with higher germination and the covariables benefit and storage time were significant (p< 0.001). Germination in native pastures tended to decrease with time, contrary to introduced ones and germination was higher in direct relation to the Cs size. The viability is best preserved by maintaining the Cs with accessory bracts; but in introduced species germination increased when they were eliminated.
Keywords: Bouteloua gracilis; Bouteloua curtipendula; classified caryopsids; grass seed
En Poaceae, la semilla o propágulo comercial consiste de espiguilla o diáspora: cariópside (Cs) + brácteas accesorias: normalmente glumas, lemas, paleas y, en ocasiones, específico de especie, ramas modificadas, las cuales tienen efecto sobre la germinación, influyen el manejo requerido y por tanto la calidad biológica de la semilla. La eliminación de brácteas accesorias con o sin clasificación de Cs por tamaño es una alternativa no documentada en el establecimiento de praderas de temporal. El objetivo del estudio fue evaluar la germinación sembrando cada 30 días, Cs clasificadas de pastos, dos nativos y dos introducidos, obtenidas a partir de diásporas comerciales al inicio del experimento (Cs base -CsB) o cada 60 días (Cs testigo-CsT), durante 15 meses; similarmente, evaluar la emergencia de plántulas a partir de diásporas. Los propágulos (diásporas o Cs) se evaluaron de febrero 2010 a mayo 2011, en laboratorio. Las especies nativas e introducidas incluyeron Bouteloua gracilis, B. curtipendula, Cenchrus ciliaris y Chloris gayana, respectivamente. A partir de diásporas, se eliminaron brácteas accesorias y las Cs resultantes, se separaron con tamices, en grandes (CsG), medianos (CsM) y chicos (CsCh). Se sembraron, en base a semilla pura viable, tanto los diferentes tamaños de Cs, sobre papel húmedo, como diásporas, a 1cm de profundidad, en peat moss. El comportamiento de la germinación se analizó mediante regresión logística con la rutina Logistic de SAS. En las especies evaluadas, mayor tamaño de Cs se relacionó con mayor germinación y las covariables beneficio y tiempo de almacenamiento fueron significativas (p< 0.001). La germinación en pastos nativos tendió a disminuir con el tiempo, contrariamente a introducidos y la germinación fue mayor en relación directa con el tamaño de la Cs. La viabilidad se conserva mejor manera al mantener la Cs con brácteas accesorias; pero, en especies introducidas eliminar éstas, incrementó la germinación.
Palabras clave: Bouteloua gracilis; Bouteloua curtipendula; cariópsides clasificados; semilla de pastos
Introduction
Commercial grass seed for arid zones is marketed as a dispersal or propagule structure (diaspora or spikelet); which consists of the caryopsid (Cs) contained in accessory bracts: glume, lemma, palea and modified twigs, depending on the species; making its sawing management difficult due to the type of structures that contain Cs that influences both the physical purity and the biological quality of the seed to be used in planting (Enríquez-Quiroz and Quero-Carrillo, 2006). The increase in the density and vigor of desirable perennial grasses in the settlement is a felt need in the arid zones of Mexico (Quero-Carrillo, 2015), due to the deterioration of the soil in response to the reduction of the vegetable cover In these areas, resulting from a number of causes, including overgrazing, deforestation and land opening for crops (SEMARNAT, 2008).
In the Chihuahuan Desert of Mexico, grasslands cover approximately 15% or less of their original area (PMAR, 2012). Therefore, in trying to recover pasture areas, the seed industry must offer the producer maximum genetic, physical and biological quality, which are aspects influenced by the production environment, harvesting, profit and storage (Probert and Hay, 2000). A solid indicator of quality includes physical purity (percentage of botanical seeds -Cs- in 100 diaspores), a cultural value impossible to see with the naked eye, unlike other crops marketed in Mexico and the world (Quero et al., 2007).
In low-precipitation zones in Mexico, the most representative native pastures of the Chihuahuan Desert include Banderita and Navajita (Rzedowski and Rzedowski, 2001), and among the introduced ones, the most widely propagated is Buffel, with approximately four million hectares (Alcalá, 1995); of which, in Sonora, 2.4 million hectares have been established (Ibarra et al., 2005) and with great potential to invade, by sowing or not, regions smaller than 800 masl, of the Chihuahuan Desert.
Another important factor is seed latency; which is characteristic of wild plants to survive adverse environments (Hilhorst, 1995; Zhongnan and Norton, 2009) and indicates the state in which the plant or organ retains its development under adequate growth conditions (Hilhorst, 1995). Grass latency can be expressed in the embryo or imposed by surrounding tissue (Simpson, 1990; Hilhorst, 1995; Tian et al., 2002). In Tripsacum dactyloides L. Tian et al. (2003) tested the influence of the pericarp and accessory bracts on latency and found that when Cs were scarified, all viable germs were able to germinate. In pastures introduced as Buffel, where latency is present both in the embryo and by influence of floral structures, germination after 18, 48 and 12 months of storage was 94%, 35% and zero (Winkwoth, 1963; Palma-Rivero, 2000; Martínez et al., 2014), respectively.
In order to determine new opportunities for improvement of pasture establishment (Poaceae), the germination by size of Cs with differential time of benefit, regarding harvest time, in comparison to diaspores, has not been documented. The objective of this paper was to evaluate the germination, every 30 days, in Banderita, Navajita and Buffel and Rhodes in beneficiated and classified Cs, both at the beginning of the experiment and every 60 days, similarly, to evaluate the emergence, every 30 days, of the sowing of diasporas, both in two native pastures and in two introduced pastures.
Materials and methods
The experiment was carried out from February 2010 to May 2011 under laboratory conditions at Montecillo Campus, Postgraduate College, Estado de Mexico. The experimental material consisted of two native species of the Chihuahuan Desert: Navajita Bouteloua gracilis (Willd. Ex Kunth) Lag. Ex Griffiths and Banderita Bouteloua curtipendula (Mich.) Torr. and on the other hand, two species introduced from Africa to Mexico, Buffel Cenchrus ciliaris L., American variety, Sin. Common or T4464 and Rhodes Chloris gayana Kunth, Bell variety. Seed, both native and introduced, was acquired as marketed on the market in January 2010; therefore, it was planted on the basis of viable pure seed, to standardize the germination or emergency opportunity (Table 1).
Table 1 Germination treatments using different dispersion unit at different time of benefit, as well as viability and germination in forage pastures.

Diásporas= cariópsides más brácteas accesorias: glumas, lema, palea, ramillas modificadas y aristas, según la especie; cariópside= CsB, material base, obtenidos al inicio del experimento y almacenados; cariópsides testigo= CsT, obtenidos cada 60 días a partir de material experimental original.
Preparation of the experimental material
At the beginning of the research, the seeds were characterized for moisture, which was 11.5 to 12%, a range considered suitable for commercial storage (FAO, 2011). Later, with a rug and a pad of corrugated rubber and manual friction, accessory bracts were removed from the Cs: gluma, lemma, palea, modified twigs, according to the species, until at the beginning of the experiment 0.5 kg of Cs (Cs Base, CsB) And 100 g every 60 days (control CsT), of each species were obtained, which were placed in plastic containers with lid and stored under laboratory conditions. To classify Cs, a considerable number of botanical seeds were taken randomly to measure length, wide and thick (mm in diameter) and were separated with manual vernier and using sieves; for Buffel and Banderita, sieve measurements were 0.5, 0.59 and 0.7 mm for small (CsCh), medium (CsM) and large (CsG) Cs, respectively. For Navajita and Rhodes, sieve sizes were 0.42, 0.5 and 0.59 mm, for CsG, CsM and CsCh, respectively.
Once the Cs were classified, either in CsB or CsT, those in good condition were selected and damaged ones were removed and grated using a stereoscopic microscope. Subsequently, both CsB, CsT, diaspores and plastic containers with lid were desinfected with commercial hypochlorite solution at 0.6% for 3 min, rinsed with distilled water and dried in the shade with light air flow for 24 h, then they were arranged on paper in work tables under laboratory environmental conditions. Subsequently, the propagules were placed by species and, if appropriate, by size, in identified containers in the shade, in a dry and fresh environment.
Subsequently, of each size and species per propagule, 50 botanical seeds were taken in four replicates to determine their viability with the 0.1% tetrazolium technique; for which the 200 Cs were soaked in distilled water for 10 h and with the aid of a stereoscopic microscope, each Cs was dissected in such a way that the embryo structures were exposed to the tetrazolium solution. The reading was performed 12 h after contact with the solution and were evaluated according to the staining shown: intense red to pink: viable; pale pink to white: not viable. The viability result for CsB was valid for germination tests at four 30-day intervals; in CsT, it was valid for two 60-day intervals and, in diaspores, valid for four 30-day intervals (Andreoli and Vilela, 2007).
Once viability was known by treatment, four replicates of 100 seeds were carried out on the basis of viable pure seed, either in classified Cs or diaspores. Before sowing, the plastic trays were disinfected with commercial hypochlorite at 0.6% for 3 min and dried in the shade for 3 h. Sowing in classified Cs was carried out in transparent plastic boxes with a cover (20*10*7 cm) and with light air flow, with the technique on wet filter paper. Diaspora seeding was carried out in trays (40 *25*10 cm) without a lid, 1.0 cm deep, in wet peat moss. The trays seeded with classified Cs and diaspores were kept in controlled environment chamber with light air flow, 24 h of fluorescent light at 22 ±2 °C. Classified Cs were irrigated every 24 h, with 8 mL of distilled water underneath the filter paper and, to diasporas, every 72 h with 100 mL of distilled water, by means of sprinkling.
Establishment of the experiment and data analysis
Sixteen germination tests were carried out in CsB and diaspores with a 30 days interval; while, for CsT, eight tests were performed with a 60-day interval. Twelve treatments (Table 1) were evaluated in CsB, 6 with the combination of two species (Banderita and Navajita) with three sizes of Cs (CsCh, CM and CG) and, on the other hand, six in CsT with the combination of two native pastures (Banderita and Navajita) with three sizes of Cs, obtained every 60 days. Twelve treatments with CsB, six with CsB with the combination of two pastures (Buffel and Rhodes) with three sizes of Cs (small, medium and large) and six with CsT, with the combination of Buffel and Rhodes, with three sizes of Cs (small, medium and large). Other treatments consisted of Cs still wrapped with accessory bracts or complete dispersion units in two native pastures (Banderita and Navajita) and on the other hand, two introduced grasses (Buffel and Rhodes).
The seedling response variable with classified Cs was normal seedling germination, which was recorded every 24 h for 15 days (ISTA, 1996). In sows with diaspores, the response variable was emergence of plums and it was recorded every 24 hours for 25 days (ISTA, 1996). Normal seedlings were those that germinated with plumule and radicle, even with slight defects and, in diaspores, with satisfactory plumule development (ISTA, 2012). The response variable was modeled using a random variable with Bernoulli (p) distribution, since for each test seed there are only two possible outcomes (the seed germinates or does not germinate).
The Bernoulli distribution depends on the parameter p, which in this case corresponds to the germination probability. The probability of germination may depend on covariates, in this case time, benefit (removal of accessory bracts) and pasture species. The problem is reduced to determine if the probabilities of germination change according to these covariates. A widely used model for this type of problem is logistic regression (Hosmer and Lemeshow, 2000).
Where: x= vector of covariables (eg. time, seed size, Cs, etc.) and β= vector of unknown parameters and correspond to the effect of covariables. The regression model was adjusted using the logistic routine of the SAS statistical package (SAS Institute Inc., 2009). The goodness of model fit was tested using the Somers D statistic (Somers, 1962). The significance of regression coefficients associated with covariates was tested using the generalized likelihood ratio test and the Wald test (α= 0.05).
Results and discussion
Native species
Germination showed a difference between treatments (p< 0.001; Figure 1). After 16 intervals of 30 days, the germination rate decreased for both CsB and CsT in each size of Cs (CsCh, CsM and CsG). Germination decreased by 39, 26 and 24 percentage units (UP) in CsB (Figure 1a) and the reduction in germination was lower in CsT than in CsB (Figure 1b) with 33, 22 and 16 UP, respectively. For CsB (Figure 1c), this reduction was 35, 19 and 4 UP, and in CsT (Figure 1d); Similarly, germination loss was lower, with 20, 15 and 7 UP, respectively. The above is indicative that the germination of the Cs stored with accessory bracts, until the time of their sowing, in CsM and CsG, maintains higher germination levels for all the sowing dates in the evaluated period.
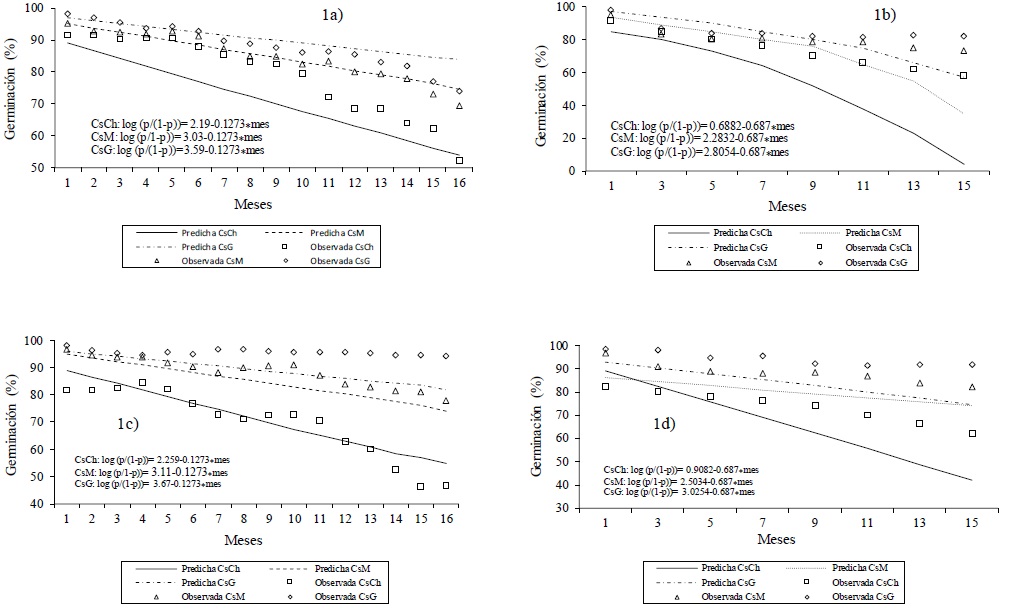
Figure 1 Observed and predicted germination of three caryopsid sizes of Bouteloua curtipendula (1a and 1b) and B. gracilis (1c and 1d) for 16 months, with a 30 day interval and for control caryopsids (obtained every two months) in laboratory.CsCh = small caryopsids; CsM = medium caryopsids; CsG = large caryopsids.
The germination curves predicted by the logistic model in native pastures along 16 germination tests (Figure 1), indicate that the germination will decrease to a greater extent, to a smaller size of Cs, either in Navajita or Banderita, or in CsB or CsT; i.e. larger size of Cs is an advantage at the moment of sowing, by the quality reached by the seed. Therefore, managing the seed production and its benefit with selection processes by size of Cs results in greater germination at the time of planting.
In native pastures, as the storage time increased, lower germination was observed at lower Cs size, for both CsB and CsT and by species evaluated. Springer et al. (2001), in T. dactyloides, indicate that viability and germination in Cs of greater weight and size, is higher, compared to that observed in CsCh; likewise Pérez et al. (2006), in maize, found that at higher weight and seed length, greater vigor, which is reflected in higher amount of dry matter of the aerial part of seedlings during the first days of establishment. Similarly, López- Castañeda et al. (1996), in wheat plants, showed that a higher weight of Cs yield in plants with greater root and leaf weight, which can result in greater seedling survival in the intra-drought period, in addition to higher initial growth; theoretically it can increase the success of establishment of plants in rain-fed meadows.
The logistic model in Navajita grass showed a difference in germination between seed sizes and, time and benefit covariables were significant (p< 0.001; Figure 1a; Figure 1b); similarly, it showed a coefficient of data adjustment of 83%, with the same covariables; on the other hand, the germination of CsG exceeded that observed for CsCh and CsM in 23 and 2.7 times, respectively. The effect of benefiting (release of Cs) improved germination four times, also the model indicates the decrease of germination of a UP every 30 days. In Banderita, the model was able to correctly classify 75% of the data, given the time and benefit covariables (Figure 1c and Figure 1d); In addition, differences in germination between sizes and covariates were significant (p< 0.001). Given the covariates mentioned, when germination is compared between Cs sizes, the germination in CsG is 5.7 times higher, compared to Cch and 1.3 times higher, compared to CsM. On the other hand, the germination of Banderita grass was higher in comparison to Navajita grass in 1.3 times (p< 0.001).
Introduced species
Contrary to what was observed for natives, germination increased according to the time of evaluation, in each size of Cs (Figure 2), differences between treatments were observed (p< 0.001). For CsB in Buffel (Figure 2a), germination (initial minus final) for Csch, CsM and CsG increased 51, 67 and 66 UP, respectively; the above, is indicative of marked training of the embryo; while for CsT of Buffel (Figure 2b), germination increased at 26, 62 and 64 UP for CsCh, CsM and CsG, respectively, which is slightly lower, compared to CsB (p< 0.001). The above is indicative that in Buffel Cs there is no difference between the benefit of the seed at the moment of planting or from the beginning of the experiment, storing the Cs in fresh and dry conditions, such as those in the laboratory, in this case.
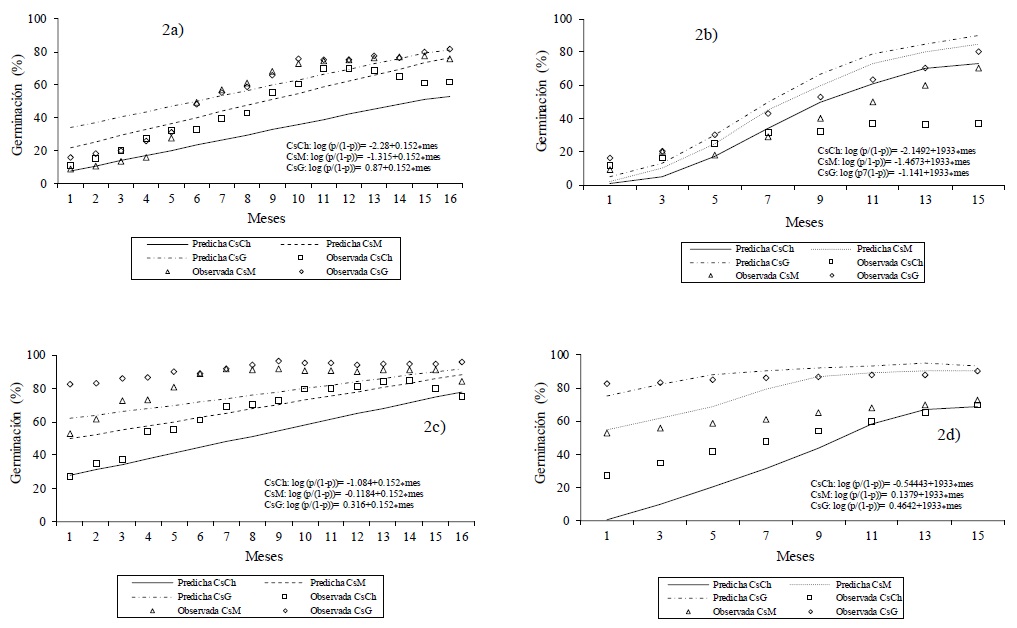
Figure 2 Observed and predicted germination of three caryopsid sizes in Cenchrus ciliaris (2a and 2b) and Chloris gayana (2c and 2d) for 16 months, 30 days interval and for caryopsids obtained every two months in the laboratory. CsCh= small caryopsids; CsM= medium caryopsids; CsG= large caryopsids.
In Rhodes, germination increased by removing accessory bracts from the beginning of the experiment (CsB; Figure 2c) which, for CsCh, CsM and CsG were 48, 31 and 13 higher UP, respectively, whereas in CsT (Figure 2d), germination was also increased for CsCh, CsM and CsG at 42, 20 and 7 UP, respectively. Therefore, higher germination occurred in CsB in all three Cs sizes, in both Buffel and Rhodes; however, for a longer storage time both Buffel and Rhodes, CsT loses latency and, at the end of germination tests, the amount of normal seedlings is slightly lower, compared to those observed for CsB. Contrary to the other three evaluated species, in Rhodes, greater germination was observed in CsB and CsT to smaller size of Cs; the above may be informative about Rhodes' ability to consolidate viable seed despite the stress conditions that the crop could face in the reproductive phase during the field phase in seed production.
The germination was higher during storage in introduced pastures and when the accessory bracts were removed at the beginning of the experiment (CsB), germination increased (Tian et al., 2002); The above, is indicative of the existence of inhibitors of germination in the embryo (Simpson, 1990); In addition, the accessory bracts are a hydrophobic barrier where lignin, phenolic polysaccharides and pectins contribute to determine the impermeable nature of the seed coat and prevent the diffusion of water for imbibition (Castillo and Guenni, 2001) and longer storage time (more than 18 months), secondary dormancy decreases in pastures like Buffel (Butler, 1985).
The removal of accessory bracts has been evaluated by Anderson (1985), in T. dactyloides, where seed was obtained and later scarified it physically with mat and corrugated rubber pad and found that germination increased from 5 to 40% and 10 to 90%. In the present study, eliminating accessory bracts increased germination, which is desirable when good planting conditions (agronomic and climatic) are combined, since the seedlings would maximize the use of moisture to reach adulthood and avoid being exposed to rapid evaporation in addition to developing photosynthetic structures and efficient nutrient assimilation for possible competition with weeds of larger size and growth capacity (Zhuang et al., 2010).
In the case of Cs Ch, CsM and CsG, the germination of Cs Ch, CsM and CsG in Navajita was marked by differences in Cs T, with 58, 73 and 82%, and for Banderita 62, 82 and 91%, respectively; while the emergence when planting at 30-day intervals was 78% and in Banderita, 87%. Tian et al. (2002), in T. dactyloides pete variety, found, at 14 days post sowing, greater germination by scarifying Cs (80%), contrasting with uncured Cs (8%) and diaspores (2%). Poor seed germination in small native grasses is associated with low vigor (Culleton et al., 1991); while, in introduced ones, this was associated with seed latency. Thus, for field plantings Ellis and Roberts (1980), Naylor (1982), Larsen and Bibby (2004), mention that the production process, harvest, storage and seed benefit are important to maintain the best biological quality of the seed and to obtain more quantity of seedlings per m2 already established in the field.
In introduced pastures, analysis with the logistic regression model indicated differences in germination between Cs sizes, time and benefit covariables were significant (p< 0.001) and germination of normal seedlings was higher as storage time progressed (Butler, 1985). In Buffel, the model manages to correctly classify 75% of the data, given the time and benefit covariables, germination increases by 2.2 times when accessory bracts are removed and increased by 1.2 times every 30 days. Also in Buffel, when comparing germination between Cs sizes, CsG showed 1.8 and 1.5 times higher germination in comparison to Cch vs CsM, respectively. In Rhodes, the model manages to explain 82% of the behavior of the data in the germination curves given the covariables time and benefit, the germination increased 6.5 times when eliminating accessory bracts and increases 1.1 times every 30 days. The germination of CsG, compared to Csch and CsM, was 16 and 2.7 times higher; therefore, to a bigger size of Cs there is greater germination. The germination observed for Rhodes was 1.4 times higher than that observed in Buffel.
The germination curves predicted by the logistic model along 16 intervals (Figure 2aa, 2d), indicate that germination increased to a larger size of Cs in both Buffel and Rhodes and at the last interval of having evaluated the seed, the highest germination in both pastures is shown; however, according to the predicted germination curves for both species, the probable germination curves will decrease.
Diaspore emergence
Differences in emergence were observed in native species (Figure 3a); which decreased with the time of storage (p< 0.001); in Navajita, it decreased six UP and in Banderita, 10 UP. The emergence of Banderita was 2.6 times higher, compared to that of Navajita. In this regard, Simpson (1990) and FAO (2011) mention that the more time of storage would affect germination, and the greater the number of abnormal seedlings will germinate. In introduced pastures (Figure 3b), difference in emergence (p< 0.001) and a positive slope were observed, as storage time (p< 0.001) advanced; Buffel increased 38 UP and Rhodes, 43 UP. The Rhodes emergency was 1.9 times greater than that of Buffel, Harty et al. (1983), in Panicum máximum and Butler (1985), in Buffel mention that the germination in grasses with latency, the germination increases after four months of storage, in Panicum spp., after 12 months and in Buffel, after 18 Months.

Figure 3 Observed and predicted emergence of seed with accessory bracts, in Bouteloua curtipendula (3a) and B. gracilis (3a);as well as in Cenchrus ciliaris (3b) cv. Common and Chloris gayana cv. Bell (3b), along 16 tests with 30 days intervals in laboratory.
In native pastures, the model managed to classify 63% of each of the cases of seed as emerged or not and in introduced, the logistic model properly adjusted, 80% of each of the cases of seed as germinated or not. The germination curves predicted in native pastures to be diminished to a small extent, indicating that the viability of the botanical seed is conserved when stored with accessory bracts, whereas in introduced pastures the emergency curves indicate that the latency imposed, both by the embryo as by accessory bracts, is smaller as the storage time grows i.e. training for germination occurs.
Conclusions
Germination of normal seedlings in native pastures was greater with a larger caryopsid size and this decreased with longer storage time. In native species, more seedlings are obtained by longer storage of the caryopsids within the accessory bracts. Banderita pasture had greater germination at each size of caryopsis, compared to Navajita. In introduced pastures, eliminating accessory bracts at the beginning of the experiment was manifested in higher germination compared to the conservation of bracts and the germination percentages were higher to longer storage time and larger caryopsid size. Pasto Rhodes showed higher germination compared to Buffel. The management of the latency of the propagule to be used influences the germination and increases the germination in native and introduced species.
Literatura citada
Alcalá, G. C. H. 1995. Origen y distribución mundial. Guía práctica para el establecimiento, manejo y utilización del zacate Buffel. PATROCIPES. Hermosillo, Sonora, México. 9-14pp. [ Links ]
Anderson, J. 1985. Aspects of the germination ecology and biomass production of eastern gama grass (Tripsacum dactyloides L.).Botanical Gazzette. 146:353-364. [ Links ]
Andreoli, C. and Vilela, de A. R. 2007. Seed longevity chart to predict viability of corn seed during open storage. Revista Brasileira de Milho e Sorgo. 6(2):247-255. [ Links ]
Butler, J. E. 1985. Germination of buffel grass. Seed Sci. Technol.13(3):583-591. [ Links ]
Castillo, R. y Guenni, O. 2001. Latencia de semillas de Stylosanthes hamata (Leguminosae) y su relación con la morfología de la cubierta seminal. Rev. Biol. Trop. 49(1):287-299. [ Links ]
Culleton, N.; McCarthy, V. and McGilloway, D. 1991. A note on the germinability and early seedling growth of Lolium perenne.Irish J. Agric. Res. 30:159-161. [ Links ]
Ellis, R. H. and Roberts, E. H. 1980. Towards a rational basis for testing seed quality. In: Hebblethwaite, P. D. (Ed.) Seed production.Butterworths, London. 605-635 pp. [ Links ]
Enríquez, Q. J. F y Quero, C. A. R. 2006. Producción de semillas de gramíneas y leguminosas forrajeras tropicales. INIFAPCIRGOC.CE Cotaxtla. Libro Técnico Núm. 11. Veracruz,Veracruz, México. 109 p. [ Links ]
FAO (Organización de las Naciones Unidas para la Alimentación y la Agricultura). 2011. Semillas en emergencias, manual técnico.Grupo de Semillas y Recursos Fitogenéticos de la División de Producción y Protección Vegetal (AGPMG) en colaboración con la División de Operaciones de Emergencia y Rehabilitación (TCE). Roma, Italia. 83 p. [ Links ]
Harty, R. L.; Hopkinson, J. M.; English, B. H. and Alder, J. 1983.Germination, dormancy and longevity in stored seed of Panicum maximum. Seed Sci. Technol. 11(2):341-351. [ Links ]
Hilhorst, H. W. M. 1995. A critical update on seed dormancy I: Primary dormancy. Seed Sci. Res. 6:61-73. [ Links ]
Hosmer, D. W. and Lemeshow, S. 2000. Applied logistic regression. 2th(Ed.). Wiley, New York. 392 p. [ Links ]
Ibarra, F. F.; Martín, M. H. R.; Moreno, S. M.; Denogean, F. G. B y Gerlach,B. L. E. 2005. El zacate Buffel como una alternativa para incrementar la rentabilidad de los ranchos en la zona serrana de Sonora. Rev. Mex. Agro. 9:521-529. [ Links ]
International Seed Testing Association (ISTA). 1996. International rules for seed testing. Seed Science and Technology 24. Supplement.243 p. [ Links ]
ISTA (International Seed Testing Association). 2012. International rules for seed testing. Seed Sci. Technol. 27:27-32. [ Links ]
Larsen, S. U. and Andreasen, C. 2004. Light and heavy seeds differing germination percentage and mean germination thermal time.Crop Sci. 44:1710-1720. [ Links ]
Larsen, S. U and Bibby, B. M. 2004. Use of germination curves to describe variation in germination characteristics in three turfgrass species. Crop Sci. 44:891-899. [ Links ]
López, C. C.; Richards, R. A.; Farquar, G. D. and Williamson, R. E. 1996.Seed and seedling characteristics contributing to variation in early vigor in early vigor among temperate cereals. Crop Sci.36:1257-1266. [ Links ]
Martínez, S. J.; Villegas, A. Y.; Enríquez, del V. J. R.; Carrillo, R. J.C. y Vásquez, D. M. A. 2014. Estrategias de escarificación para eliminar latencia en semillas de Cenchrus ciliaris L. y Brachiaria brizantha cv. Marandú. Rev. Mex. Cienc. Agríc.6:1263-1272. [ Links ]
Naylor, R. E. L. 1982. Differences between cultivars of perennial ryegrass in laboratory germination and field emergence. Ann. Appl.Biol. 100:106-107. [ Links ]
Palma, R. M. P.; López, H. A y Molina, M. J. C. 2000. Condiciones de almacenamiento y germinación de semillas de Cenchrus ciliaris L. y Andropogon gayanus Kunth. Agrociencia. 34:41-48. [ Links ]
Pérez, M. C.; Hernández, L. A.; González, C. F. V.; García de los S. G.;Carballo, C. A.; Vásquez, R. T. R. y Tovar, G. M del R. 2006.Tamaño de semilla y relación con su calidad fisiológica en variedades de maíz para forraje. Agric. Téc. Méx. 32(3):341-352. [ Links ]
PMAR (Plan Maestro de la Alianza Regional). 2012. Plan maestro de la alianza regional para la conservación de los pastizales del Desierto Chihuahuense. In: Guzmán-Aranda, J. C.; Hoth, J. y Berlanga, H. (Eds.). Comisión para la Cooperación Ambiental.Montreal. Canadá 64 p. [ Links ]
Probert, R. J. and Hay, F. R. 2000. Keeping seed alive. In: Bewley, D. J., and Black M. (Eds.). Seed technology and its biological basis.CRC press LLC. Great Britain. 390-393 pp. [ Links ]
Quero, C. A. R.; Enríquez, Q. J. F. y Miranda, J. L. 2007. Evaluación de especies forrajeras en América tropical, avances o status quo.Interciencia. 32:566-571. [ Links ]
Quero, C. A. R. Pastoreo actual y profesionalizado. Importancia e impacto.In: memorias del VI congreso internacional de manejo de pastizales. Conferencia magistral. SOMMAP, MVZ, UJED,septiembre 23 a 25, 2015. Durango, Durango. 6 p. [ Links ]
Rzedowski, G. C. y Rzedowski J. 2001. Flora fanerogámica del valle de México. 2a (Ed.). Instituto de Ecología y Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Pátzcuaro,Michoacán, México. 504 p. [ Links ]
SEMARNAT (Secretaría del Medio Ambiente y Recursos Naturales).2008. Informe de la situación del medio ambiente en México.Compendio de estadísticas ambientales. México, D. F. 357 p. [ Links ]
Simpson, G. M. 1990. Seed dormancy in grasses. Cambridge University Press, Cambridge, UK. 296 p. [ Links ]
Somers, R. H. 1962. A new asymmetric measure of association for ordinal variables. Am. Sociol. Review. 27(6):799-811. [ Links ]
SAS (Statistical Analysis System). SAS Institute Inc. 2009. SAS/STAT® 9.3.1 User’s Guide. Cary, NC, USA. 906 p. [ Links ]
Springer, T. L.; Dewald, C. L and Aiken, G. E. 2001. Seed germination and dormancy in Eastern Gamagrass. Crop Sci. 41:1906-1910. [ Links ]
Tian, X.; Knapp, A. D.; Moore, K. J.; Brummer, E. C. and Bailey, T. B.2002. Cupule removal and caryopsis scarification improves germination of eastern gamagrass seed. Crop Sci. 42:185-189. [ Links ]
Tian, X.; Knapp, A. D.; Gibson, L. R.; Struthers, R.; Moore, K. J.; Brummer,E. C. and Bailey, T. B. 2003. Response of eastern gamagrass seed to gibberellic acid buffered below its Pka. Crop Sci. 43:927-933. [ Links ]
Winkworth, R. E. 1963. The germination in buffel grass (Cenchrus ciliaris) seed after burial in a Central Australian soil. Austr. J.Exp. Agric. Animal Husbandry. 3(11):326-328. [ Links ]
Zhongnan, N. and Norton, M. R. 2009. Stress tolerance and persistence of perennial grasses: the role of the summer dormancy trait in temperate Australia. Crop Sci. 49:2405-2411. [ Links ]
Zhuang, J.; McCarthy, J. F and Perfect, E. 2007. Soil water hysteresis in water-stable microaggregates as affected by organic matter.Soil Sci. Soc. Am. J. 72:212-220. [ Links ]
Received: January 2017; Accepted: March 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
