










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.8 no.1 Texcoco ene./feb. 2017
https://doi.org/10.29312/remexca.v8i1.77
Articles
Effect of potassium on the phenolic content and antioxidant capacity of Ocimum basilicum L.
1Universidad Politécnica de Gómez Palacio. Carretera Gómez Palacio-Chihuahua, km 6.5. Ejido el Vergel. Gómez Palacio, Durango, México. CP. 35 120. (pepe_dimaslopez@hotmail.com; lsalas@upgop.edu.mx; ramonvalzs@hotmail.com; vborroel@upgop.edu.mx).
2Instituto Tecnológico de Torreón. Carretera Torreón-San Pedro, km 7.5. Torreón, Coahuila, México. CP. 27170. (ppreciador@yahoo.com.mx).
3Universidad del Papaloapan, Campus Loma Bonita. Ferrocarril s/n, San Antonio. Loma Bonita, Oaxaca, México. CP. 68 400. (ana_ramirez04@hotmail.com).
In the last decade, numerous efforts have been made to increase the content of phenolic compounds and the antioxidant capacity of plants that go directly to human consumption, using for this several methods of cultivation; however, the results have been discrepant about the quality of the fruits and the yield obtained, concluding that the high content of phenolic compounds and the antioxidant capacity has not been a guarantee of high crop yields. Therefore, the objective of this study was to determine a possible increase in total phenolic compounds, total antioxidant capacity and yield of three basil genotypes (Sweet Lemon, Red Rubin and Cinnamon) produced under different levels of potassium (7,9 , 11 and 13 mM). In addition, dry matter, plant height and number of leaves were evaluated. The work was carried out in the Comarca Lagunera of Durango in the year 2014. The analysis of variance showed significant differences (p≤ 0.001) due to genotype and potassium levels in almost all variables. It was found interaction in total phenolic compounds, antioxidant capacity, yield and height of the plant. The correlation (R2) (p≤ 0.05) was also found between the potassium level and all variables. The results indicate that potassium control in the nutrient solution may be a viable option to improve nutraceutical qualities in terms of total phenolic compounds content and antioxidant capacity without affecting basil yield.
Keywords: Ocimum basilicum L.; antioxidants; potassium concentration; yield
En la última década, se han realizado numerosos esfuerzos para incrementar el contenido de compuestos fenólicos y la capacidad antioxidante de plantas que vayan directo al consumo humano, empleando para esto diversos métodos de cultivo; sin embargo, los resultados han sido discrepantes acerca de la calidad de los frutos y el rendimiento obtenido, concluyendo que el alto contenido de compuestos fenólicos y la capacidad antioxidante no ha sido garantía de alto rendimiento de los cultivos. Por lo tanto, el objetivo de este estudio fue determinar un posible aumento del contenido en compuestos fenólicos totales, capacidad antioxidante total y rendimiento de tres genotipos de albahaca (Sweet Lemon, Red Rubin y Cinnamon) producidos bajo diferentes niveles de potasio (7, 9, 11 y 13 mM). Adicionalmente, se evaluó materia seca, altura de planta y número de hojas. El trabajo se llevó a cabo en la Comarca Lagunera de Durango en el año 2014. El análisis de varianza mostró diferencias significativas (p≤ 0.001) por efecto de genotipo y nivel de potasio en casi todas las variables. Se encontró interacción en compuestos fenólicos totales, capacidad antioxidante, rendimiento y altura de la planta. También se encontró correlación (R2) (p≤ 0.05) entre el nivel de potasio y todas las variables. Los resultados indican que el control de potasio en la solución de nutrientes puede ser una opción viable para mejorar las cualidades nutracéuticas en cuanto al contenido de compuestos fenólicos totales y la capacidad antioxidante sin afectar el rendimiento de albahaca.
Palabras clave: Ocimum basilicum L.; antioxidants; concentración de potasio; rendimiento
Introduction
In recent years, many efforts have been made to increase the content of phenolic compounds and the antioxidant capacity of plants that go directly to human consumption, using different methods of cultivation such as the application of organic and conventional fertilization; in open fields and in greenhouses (Kokaly and Skidmore, 2015). However, the results have been discrepant about the quality of the plant and the yield obtained, concluding that the high content of phenolic compounds and antioxidant capacity is not a guarantee of high crop yields (Salama et al., 2013). This circumstance should be considered in view of the growing global population, high costs of crop production, and issues related to environmental pollution, which creates the need to improve and select more efficient production systems for maintaining food production (Henderson et al., 2016).
In addition to the environmental impacts caused by agricultural production systems, the effects on human health due to new eating habits characterized by nutrient and stress disorders have increased the number of chronic diseases such as obesity, diabetes, cancer, cardiovascular diseases and neurological disorders among others (Lee and Scagel, 2009). Therefore, it is necessary to unite in a global effort on food, health and the environment, in order to produce safe foods that promote health and reduce the risk of developing chronic diseases (Tilman and Clark, 2014). In this sense, it is important to propose protocols based on biological systems controlling the application of nutrients in adequate nutritive solutions to improve the nutritional and nutraceutical quality of plants (Kocira et al., 2016).
In this context, in addition to obtaining quality harvests in the phenolic content and antioxidant capacity, it is important to obtain acceptable yields to avoid economic losses, so that the production of medicinal plants and the control of nutrients in the nutrient solution could be an alternative to the issues raised. In particular, potassium (K) is one of the mineral nutrients required in larger quantities by plants and has physiological implications in cell maintenance and stomatal opening (Nguyen et al., 2010). In addition, K promotes the rapid accumulation and translocation of carbohydrates and activates the processes that are necessary for the conservation of the water state of the plant and the turgidity pressure of the cells (Pottosin et al., 2005). This element is essential for many physiological processes, such as photosynthesis, osmoregulation, cell turgor and homeostasis in the plant cell (Mohd Zain and Ismail, 2016).
Some studies indicate that K, besides promoting the synthesis of amides and proteins, is also an enzymatic activator (Inthichack et al., 2012). Its role as an enzyme activator has been linked to the production of phytonutrients and therefore has implications for the biochemical synthesis of secondary metabolism products of plants (Mohd Zain and Ismail, 2016; Inthichack et al., 2012). Some authors point out that K favors the increase of phenolic compounds (Ibrahim et al., 2012), which are a large and heterogeneous group of secondary metabolites, which are distributed throughout the plant kingdom (Abbasi et al., 2015). The phenolic compounds are among the most desirable phytochemicals due to their antimicrobial and antiviral activity; as well as its anti-inflammatory and antioxidant properties (Olennikov et al., 2011).
On the other hand, one of the plants widely reported for its content of phenolic compounds and antioxidant capacity is basil (Ocimum basilicum L.), which is an ornamental, culinary and medicinal species belonging to the family Lamiaceae, cultivated worldwide and produced under a wide variety of growth conditions (Kwee and Niemeyer, 2011).
Basically, basil is a plant with a wide variety of food and medicinal applications (Flanigan and Niemeyer, 2014; Koca and Karaman, 2015); however, there is little information that this species can increase its phenolic content and antioxidant capacity without affecting its performance. Therefore, the objective of this study was to determine a possible increase in the content of phenolic compounds, antioxidant capacity and yield, in addition to other agronomic parameters such as dry matter, plant height and leaf number in three basil genotypes (Sweet Lemon, Red Rubin and Cinnamon) by applying different concentrations of potassium.
Materials and methods
The work was carried out in the Comarca Lagunera, located between the states of Coahuila and Durango, Mexico, during the spring of 2014 in the Locality “El Quemado”, which belongs to the municipality of Gomez Palacio, Durango, Mexico. The experiment was carried out in a greenhouse with semi-automatic temperature regulation, with values between 25-30 ºC and 70-80% for temperature and humidity, respectively.
Three genotypes of basil (Ocimum basilicum L.) were selected: Sweet Lemon, Red Rubin and Cinnamon because they represented a wide range of morphological and genetic characteristics (Lee and Scagel, 2009). The basil seeds were obtained as a donation from the Universidad Juarez of the State of Durango (UJED). The seeding was carried out in 250-well polystyrene germination trays (three seeds per well) filled with perlite (Environment®). The trays were watered twice a day with tap water 21 days after sowing, the seedlings showed 3 to 4 true leaves and a height of 0.1 to 0.15 m, therefore, the seedlings were transferred to black plastic pots (one gallon capacity) containing a mixture of sand: perlite (80:20).
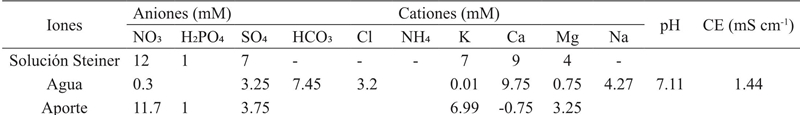
The preparation of the nutrient solution was performed using Steiner’s universal method, which indicates a balance between anions and cations (Steiner, 1961) (Table 1).
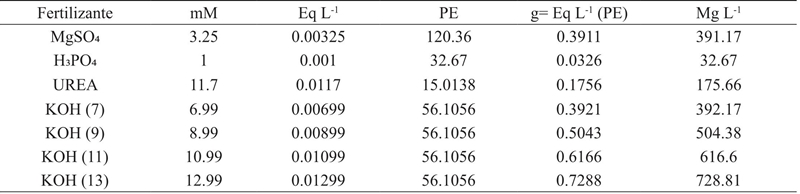
On the other hand, Table 2 shows the calculations made for the application of K (7, 9, 11 and 13 mM). The KOH was used as the source of potassium and the requirements of the anions and cations were calculated from the equivalent weight of the fertilizer (Favela et al., 2006). Once the seedlings were transferred and the nutrient solutions were prepared, the pots were placed in a completely randomized 3*4 factorial block design with six replicates per treatment (n= 72 plants).
The nutrient application started the day of transplantation and was repeated twice a day (9:00 am and 6:00 pm). The harvest was performed 40 days after the transplant. The harvested plants were transferred to the Laboratory of Biochemistry of the Polytechnic University of Gomez Palacio for its preparation. The fresh basil was washed with 3% sodium hypochlorite solution and placed on brown paper for drying at room temperature (25 ± 2 ºC) for 15 days.
The dried samples were ground in a blender (Hamilton Beach) and stored at 5 °C for subsequent extraction. A solid-liquid extraction was performed by the addition of 20 g of sample in 100 ml of ethanol. A “Stuart” stirrer was used to keep the mixture under stirring for 24 h at 30 °C. After 24 h, the sample was dried under vacuum and the ethanol was rotary evaporated (Buchi, Model-210) and a water bath at 35-40 °C. The obtained extracts were stored in refrigeration until their respective analysis.
The total phenolic content was measured using a modification of the Folin-Ciocalteu method (Esparza et al., 2006). 30 μl of extract was mixed with 270 μl of distilled water in a test tube. Then 1.5 ml of Folin-Ciocalteu reagent (Sigma-Aldrich, St Louis MO, EE.UU.) diluted (1:15) was added to the mixture, with vortexing for 10 s. After 5 min 1.2 ml of sodium carbonate (7.5% w/v) was added and stirred for 10 s. The solution was placed in a water bath at 45 °C for 15 min, and then allowed to cool to room temperature. The absorbance of the solution was read at 765 nm on a UV Genesys 10 spectrophotometer. The phenolic content was calculated by a calibration curve using gallic acid as standard and the results were recorded in mg gallic acid equivalent per 100 grams extract (mg AGE/100 g extract).
The determination of the total antioxidant capacity of the different samples was carried out based on the method Brand-Williams et al. (1995) with slight modifications. The free radical solution 1.1-diphenyl-2-picrylhydrazyl (DPPH) (Aldrich, St. Louis MO, USA) was prepared in a flask completely covered with DPPH+ foil (5 mg/100 ml ethanol grade analytical). The mixture was shaken vigorously and the flask kept covered to prevent rapid degradation. 300 μl sample of the extract diluted in triplicate test tubes and 1 200 μl of distilled water was shaken at 3000 rpm for 10 s. 1 ml of DPPH+ was added and vortexed again at 3000 rpm for 10 s. The readings were performed at 517 nm after 90 min. The total antioxidant capacity was calculated using a standard curve with the reference antioxidant Trolox and the results were expressed in μM Trolox per 100 grams extract (μM Trolox/100 g extract).
After harvesting, plant height was measured from the basal to the apical part with a retractable tape measuring 5 m in length (Moncayo et al., 2015). Subsequently, the plants were cut from the base for performance measurement. The yield values were obtained by weighting the plant on a granataria scale (OHAUS) and reported as kilograms per plant on a fresh weight basis (kg planta/PF). To determine the number of leaves, they were separated from the stems and quantified. The dry matter (MS) was quantified according to the method established by the AOAC (2005), placing 12 g of the fresh sample in aluminum boxes in a forced air oven at 70 °C to constant weight. All analyzes and measurements were performed in triplicate.
The data of the variables for factors and interactions were analyzed by an analysis of variance using the statistical software SAS (1999). In addition, a correlation and regression of the obtained data was obtained. The Tukey’s test was used for the media comparisons (p< 0.05).
Results and discussion
The interest in the determination of phenolic compounds in basil in this study was to elucidate the mechanisms of biosynthesis as a result of the change in potassium (K) concentration. The results of this study showed highly significant differences (p≤ 0.001) for total phenolic compounds (CFT). These were affected by genotype (G) and potassium level (NK) (Figure 1). As can be seen in Figure 1a Sweet Lemon cultivar showed the highest phenolic content (1 353 mg AGE/100 g extract) followed by Cinnamon and Red Rubin cultivars (1 251 and 230 mg AGE/100 g of extract, respectively). Javanmardi et al. (2003) studied total phenolic compounds in 23 different basil cultivars and reported values of 22.9 to 65.5 mg AGE/g dry weight (PS).
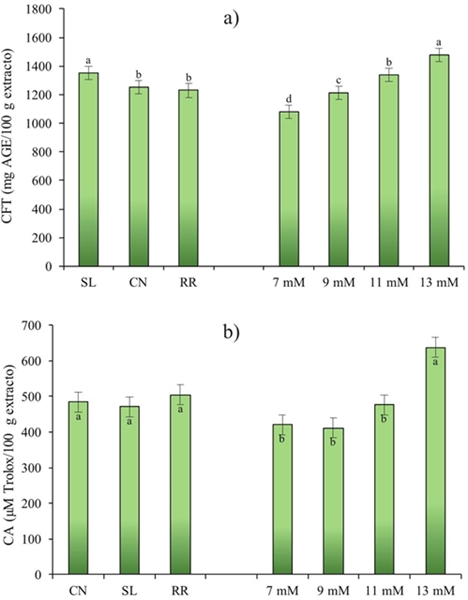
Figure 1 Effect of cultivar (C) and level of K (NK) on the content of total phenolic compounds (a); and antioxidant capacity (b), from Ocimum basilicum L. CN= Cinnamon;SL= Sweet Lemon; RR= Red Rubin.
On the other hand, Kwee and Niemeyer (2012) in a study of 15 basil cultivars indicated ranges from 3.47 to 17.58 mg AGE/g PS. On the other hand, Flanigan and Niemeyer (2014) in 10 cultivars found values of 13.1 to 26.9 mg AGE/g PS. The mentioned authors indicated that the phenolic content in plants is determined not only by the cultivar, but also by the conditions and place of cultivation, in addition the concentration of total and individual phenolics like flavonoids and phenolic acids can vary by said parameters. According to the NK factor, it was observed that the higher the potassium level the higher the total phenolic compounds content. Thus, the highest content of phenolic compounds (1 180 mg AGE/100 g extract) was obtained with 13 mM (Figure 1a).
These results are in agreement with other data reported in the literature. For example, Knuyen et al. (2010) evaluated concentrations of K in the range of 1 to 5 mM and obtained values of phenolic compounds from 7 to 16 mg AGE/g PS. In another study, Ibrahim et al. (2012) reported an increase of 1.22 to 1.82 mg AGE/g extract by applying 0 to 270 kg of K ha-1 in Labisia pumila Benth. These results coincide with this study because they confirm a correlation between potassium uptake and phenolic compounds content in the plant (Table 3), with a significant positive correlation in all cultivars (R2= 0.972) between NK and CFT. In addition, an interaction between genotype and potassium level (G*NK) was found (Figure 2a). The interaction shown suggests a linear increase in the phenolic content of the three genotypes in response to increased potassium level. Generally, the K+ ion regulates the internal balance for the purpose of absorption and translocation of carbohydrates that indirectly affect the formation of the basic structures of phenylpropanoids (Kuum et al., 2015). However, it is not conclusive that the increase of the phenolic compounds content in the plant can always be linear with the increase of the application of K, therefore, it could be interesting to continue investigating the range of application of K, with the possibility of find a quadratic response to relate the level of K and the production of total phenolic compounds.
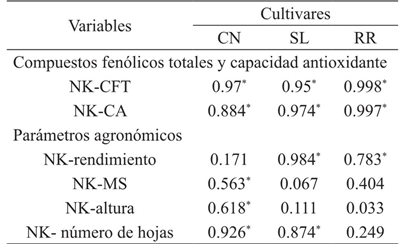
Table 3 Correlation coefficient R2 between potassium concentration and total phenolic compounds content, antioxidant capacity and agronomic parameters in Ocimum basilicum L.
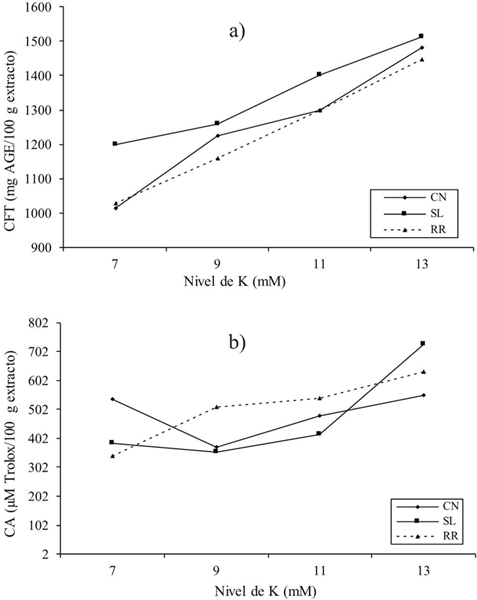
Figure 2 Interaction of the cultivar and level of K (C*NK) for the content of total phenolic compounds (a); and antioxidant capacity (b) from Ocimum basilicum L.CN= Cinnamon; SL= Sweet Lemon; RR= Red Rubin.
Regarding antioxidant capacity (CA) in this study no significant difference was found between genotypes, but for potassium level (p≤ 0.05) (Figure 1b). The antioxidant capacity of basil has been reported in numerous studies (Szymanowska et al., 2015), indicating that regardless of genotype, basil is a potential source of bioactive compounds with antioxidant capacity. On the other hand, the results showed that the antioxidant capacity of basil was higher with the increase of the potassium level, demonstrating this with the significant positive correlation obtained (R2= 0.951) between NK and CA in the three genotypes studied (Table 3 ).
The interaction G*NK for phenolic compounds was highly significant (p≤ 0,001) (Figure 2b), indicating that one or more levels of potassium used affect the genotypes under study. Red Rubin showed a linear response with increasing NK, although the response was similar at 9 and 11 mM, and a higher antioxidant capacity was obtained at 13 mM. Meanwhile Cinnamon presented this linear response in the range of 9 to 13 mM; however, it is of interest to note that level 7 obtained a value similar to level 13. Finally, Sweet Lemon has similar antioxidant capacity in the range of 7 to 11 and increases considerably by 13 mM.
On the other hand, a correlation between total phenolic compounds and antioxidant capacity (R2= 0.65) was obtained (Figure 3), suggesting that 65% of the antioxidant capacity obtained is due to the phenolic compounds content.
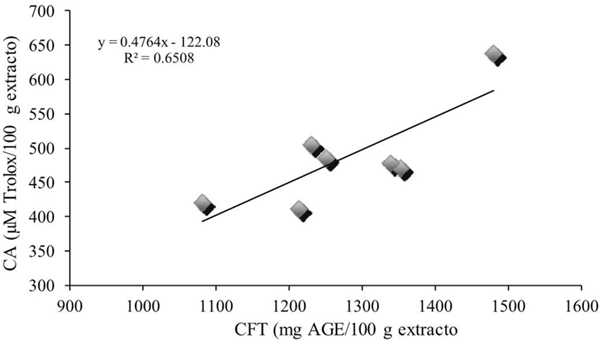
Figure 3 Correlation between the content of total phenolic compounds and the antioxidant capacity of Ocimum basilicum L.
The performance values showed highly significant differences (p≤ 0.001) for G and NK (Figure 4a). The highest yield value was obtained with Cinnamon (0.938 kg plant-1 PF), followed by Sweet Lemon and Red Rubin (0.9 and 0.87 kg plant-1 PF, respectively). For NK factor, yields were statistically similar with the levels of 9, 11 and 13 mM, the lowest yield was 7 mM.
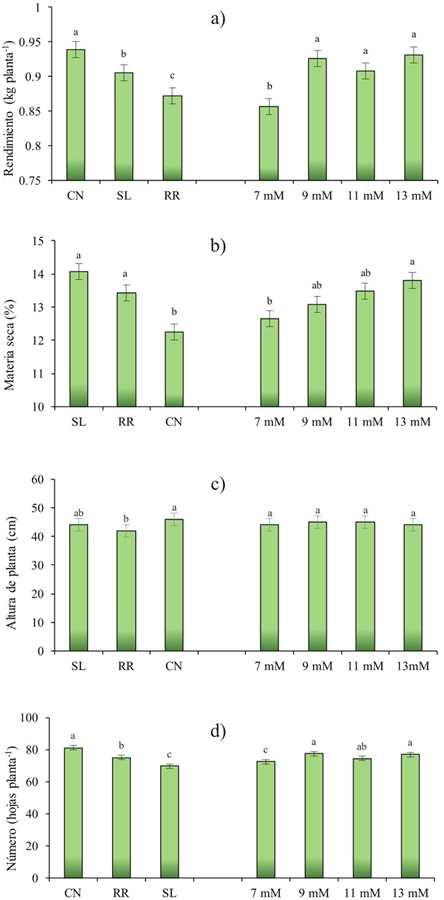
Figure 4 Effect of cultivar (C) and level of K (NK) on yield (a); dry matter (b); plant height (c) and number of leaves (d); from Ocimum basilicum L. CN= Cinnamon; SL=Sweet Lemon; RR= Red Rubin.
For plant height, analysis of variance showed highly significant differences (p≤ 0.001) due to genotype (G), but there was no difference for potassium (NK) factor (Figure 4c). These results could indicate that NK does not affect the height of the plant, but this variable is related to the genotype (Al-Kateb and Mottram, 2014), being a factor to consider when choosing a cultivar with this characteristic. As can be seen in Figure 4c Cinnamon exhibits greater height (46 cm), next to Sweet Lemon (44 cm) and these are followed by Red Rubin (42 cm).
In addition, a highly significant interaction (p≤ 0.001) was obtained for plant height due to G and NK (G*NK) (Figure 4). In the Figure 4c shows that in Cinnamon the height of the plant was higher in the range of 9 mM and 11 mM and the lowest in 7 and 13 mM. Sweet Lemon have similar values at all levels of K; and Red Rubin showed similar values in the range of 9 to 13 mm and higher in 7 mM. The Height is a parameter that is influenced by the genetic characteristics of the plant; however, their own genetic characteristics produce different biological responses that define a particular state of the organism in an optimal range (Li et al., 2016).
Rubin is a variety of dark opal basil, being a crop that has reddish-purple leaves in color and a stronger flavor than sweet basil (Bekhradi et al., 2015). Possibly its lower height could be related to a physiological and biochemical response in the primary and secondary metabolism due to the mobilization and translocation of the nutrients in the plant, producing a lower height and directing the response to pigment concentration (Landi et al., 2013). The number of leaves resulted in highly significant differences (p≤ 0.001) for G and NK (p≤ 0.05) (Figure 4). The Figure 4d shows that Cinnamon had the highest number of leaves (81.67 leaves per plant) followed by Red Rubin and Sweet Lemon (75 and 69 leaves per plant, respectively). The number of leaves was found within the ranges reported in other studies, as indicated by Moncayo et al. (2015) in a study with white basil, where values ranged from 66.5 to 103 leaves per plant.
In this study it was observed that with 9 mM of K, more leaves per plant were obtained (77.73), followed by the level of 13 mM (76.88) and 11 mM (74.33). Finally the lower number of leaves was obtained with 7 mM. Some authors suggest that the development of the leaf is initially related to the appearance of the primordial leaf at the apex, which may depend on the genetic characteristics of the plant, then depends on environmental and nutritional factors (Uzun, 2006). In a study in the melon it was found that there are differences in the number of leaves by the concentration of K, indicating that the physiological importance of this nutrient is in a greater area of impact for the photosynthesis and, therefore, a greater production of carbon skeletons, which will be used or stored for later and transported to demand sites (Preciado et al., 2002). The effect of potassium on the number of leaves can be demonstrated by the correlation obtained (Table 2) in Cinnamon and Sweet Lemon where it could be suggested that 92 and 87% (respectively) of the number of leaves in these cultivars is due to the increase in the number of leaves concentration of potassium.
Conclusions
In this work a manipulation of the potassium concentration in the nutrient solution was carried out to increase the production of phenolic compounds and the antioxidant capacity of basil as an option to create added value to the crop. It was shown that the content of the phenolic compounds and the antioxidant capacity of the basil could be improved by selecting cultivars and increasing the potassium concentration during plant growth. A significant linear correlation was obtained between the applied potassium concentration and the total content of phenolic compounds and the antioxidant capacity for the three basil genotypes investigated. For the yield it is concluded that the results show that this is affected by the increase in the potassium concentration and that it varies by the selection of the genotype.
Literatura citada
AAbbasi, A. M.; Guo, X.; Fu, X.; Zhou, L.; Chen, Y.; Zhu, Y.; Yan, H. and Liu, R. H. 2015. Comparative assessment of phenolic content and in vitro antioxidant capacity in the pulp and peel of mango cultivars. Int. J. Mol. Sci. 16:13507-13527. [ Links ]
Al, K. H. and Mottram, D. S. 2014. The relationship between growth stages and aroma composition of lemon basil Ocimum citriodorum Vis. Food Chem. 152: 440-446. [ Links ]
AOAC (Association of Oficial Analytical Chemist). 2005. Official methods of analysis 18th (Ed.). AOAC International.Gaithersburg, MD, EEUU. http://www.aoac.org. [ Links ]
Bekhradi, F.; Luna, M. C.; Delshad, M.; Jordan, M. J.; Sotomayor, J. A.;Martínez, C. C. and Gil, M. I. 2015. Effect of deficit irrigation on the postharvest quality of different genotypes of basil including purple and green Iranian cultivars and a Genovese variety.Postharvest Biol. Technol. 100:127-135. [ Links ]
Brand, W. W. and Berset, C. 1995. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. u. Technol. 28:25-30. [ Links ]
Bugarín, M. R.; Galvis, S. A.; Sánchez, G. P. y García, P. D. 2002.Acumulación diaria de materia seca y de potasio en la biomasa aérea total del tomate. Terra Latinoam. 20(4):401-409. [ Links ]
Delgado, R.; Núñez, U. and Velásquez, L. 2004. Acumulación de materia seca, absorción de nitrógeno, fósforo y potasio por el maíz en diferentes condiciones de manejo de la fertilización nitrogenada. Agron. Trop. 54(4):371-390. [ Links ]
Esparza, R. J. R.; Stone, M. B.; Stushnoff C.; Pilon, S. E. and Kendal, P.A. 2006. Effects of ascorbic acid applied by two hydrocooling methods on physical and chemical properties of green leaf lettuce stored at 5 C. J. Food Sci. 71(3):S270-S276. [ Links ]
Favela, Ch. E.; Preciado, R. P. y Benavidez, M. A. 2006. Manual para la preparación de soluciones nutritivas. Universidad Autónoma Agraria Antonio Narro (UAAN). 543 p. [ Links ]
Flanigan, P. M. and Niemeyer, E. D. 2014. Effect of cultivar on phenolic levels, anthocyanin composition, and antioxidant properties in purple basil (Ocimum basilicum L.). Food Chem 164: 518-526. [ Links ]
Henderson, B.; Godde, C.; Medina, H. D.; Van, W. M.; Silvestri, S.;Douxchamps, S.; Stephenson, E.; Power, B.; Rigolot, C.;Cacho O. and Herrero, M. 2016. Closing system-wide yield gaps to increase food production and mitigate GHGs among mixed crop- livestock smallholders in Sub-Saharan Africa.Agric. Syst. 143:106-113. [ Links ]
Ibrahim, M. H.; Jaafar, H. Z.; Karimi E. and Ghasemzadeh, A. 2012.Primary, secondary metabolites, photosynthetic capacity and antioxidant activity of the Malaysian Herb Kacip Fatimah (Labisia pumila Benth) exposed to potassium fertilization under greenhouse conditions. Int. J. Mol. Sci. 13:15321-15342. [ Links ]
LInthichack, P.; Nishimura Y. and Fukumoto, Y. 2012. Effect of potassium sources and rates on plant growth, mineral absorption, and the incidence of tip burn in cabbage, celery, and lettuce. Hortic.Environ. Biotechnol. 53:135-142. [ Links ]
Javanmardi, J. 2003. Antioxidant activity and total phenolic content of Iranian Ocimum accessions. Food Chemistry 83:547-550. [ Links ]
Koca, N. and Karaman, S. 2015. The effects of plant growth regulators and L-phenylalanine on phenolic compounds of sweet basil.Food Chem 166:515-521. [ Links ]
Kocira, A.; Świeca, M.; Kocira, S.; Złotek U. and Jakubczyk, A. 2016.Enhancement of yield, nutritional and nutraceutical properties of two common bean cultivars following the application of seaweed extract (Ecklonia maxima). Saudi J. Biol. Sci. [ Links ]
Kokaly, R. F. and Skidmore, A. K. 2015. Plant phenolics and absorption features in vegetation reflectance spectra near 1.66 μm. Inter.J. Appl. Earth Observ. Geoinf. 43:55-83. [ Links ]
Kuum, M.; Veksler V. and Kaasik, A. 2015. Potassium fluxes across the endoplasmic reticulum and their role in endoplasmic reticulum calcium homeostasis. Cell Calcium. 58:79-85. [ Links ]
Kwee, E. M. and Niemeyer, E. D. 2011. Variations in phenolic composition and antioxidant properties among 15 basil (Ocimum basilicum L.) cultivars. Food Chem. 128:1044-1050. [ Links ]
Landi, M.; Pardossi, A.; Remorini D. and Guidi, L. 2013. Antioxidant and photosynthetic response of a purple-leaved and a green-leaved cultivar of sweet basil (Ocimum basilicum) to boron excess.Environ. Exp. Bot. 85:64-75. [ Links ]
Lee, J. and Scagel, C.F. 2010. Chicoric acid levels in commercial basil (Ocimum basilicum) and Echinacea purpurea products. J. Func.Foods 2:77-84. [ Links ]
Li, F.; Chen, B.; Xu, K.; Gao, G.; Yan, G.; Qiao, J.; Li, J.; Li, H.;Li, L.; Xiao, X.; Zhang, T.; Nishio T. and Wu, X. 2016. A genome-wide association study of plant height and primary branch number in rapeseed (Brassica napus). Plant Sci.242:169-177. [ Links ]
Mohd Zain, N. A. and Ismail, M. R. 2016. Effects of potassium rates and types on growth, leaf gas exchange and biochemical changes in rice (Oryza sativa) planted under cyclic water stress. Agric.Water Manag. 164:83-90. [ Links ]
Moncayo, L. M. D. R.; Alvarez, R. V. D. P.; González C. G.; Salas P.L. y Chávez S. J. A. 2015. Producción orgánica de albahaca en invernadero en la comarca lagunera. Terra Latinoam.33(1):69-78. [ Links ]
Nguyen, P. M.; Kwee E. M. and Niemeyer, E. D. 2010. Potassium rate alters the antioxidant capacity and phenolic concentration of basil (Ocimum basilicum L.) leaves. Food Chem. 123:1235-1241. [ Links ]
Olennikov, D. N.; Kashchenko, N. I.; Chirikova N. K. and Kuz’mina, S.S. 2015. Phenolic profile of Potentilla anserina L. (Rosaceae) herb of siberian origin and development of a rapid method for simultaneous determination of major Phenolics in P. anserina pharmaceutical products by microcolumn RP-HPLC-UV. Molecules 20:224-248. [ Links ]
Peil, R. M. and Galvez, J. 2012. Reparto de materia seca como factor determinante de la producción de las hortalizas de fruto cultivadas en invernadero. Current Agric. Sci. Technol. 11(1). [ Links ]
Pottosin, I. I.; Muñiz J. and Shabala, S. 2005. Fast-activating cannel controls cation fluxes across the native chloroplast envelope.J. Membrane Biol. 204:145-156. [ Links ]
Preciado, R. P. ; Baca, C. G. A.; Tirado, T. J. L.; Kohashi-Shibata J.;Tijerina, CH. L. y Martínez, G. A. 2002. Nitrógeno y potasio en la producción de plántulas de melón. Terra. 20(3):267-276. [ Links ]
Salama, Z. A.; El Baz, F. K.; Gaafar A. A. and Zaki, M. F. 2015. Antioxidant activities of phenolics, lavonoids and vitamin C in two cultivars of fennel (Foeniculum vulgare Mill.) in responses to organic and bio-organic fertilizers. J. Saudi Soc. Agric. Sci. 14:91-99. [ Links ]
SAS (Statistical Analysis System) Institute. 2001. SAS user’s guide.Statistics. Version 9.0. SAS Inst., Cary, NC. USA. Quality, and elemental removal. J. Environ. Qual. 19:749-756. [ Links ]
Steiner, A. A. 1961. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil. 15(2):134-154. [ Links ]
Szymanowska, U. 2015. Anti-inflammatory and antioxidative activity of anthocyanins from purple basil leaves induced by selected abiotic elicitors. Food Chem. 172:71-77. [ Links ]
Tilman, D. and Clark, M. 2014. Global diets link environmental sustainability and human health. Nature 515:518-522. [ Links ]
Uzun, S. 2006. The quantitative effects of temperature and light on the number of leaves preceding the first fruiting inflorescence on the stem of tomato (Lycopersicon esculentum, Mill.) and aubergine (Solanum melongena L.). Scientia Hortic. 109:42-14 [ Links ]
Received: December 2016; Accepted: February 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
