











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El uso de híbridos trilineales de maíz (Zea mays) en México, incluyendo los Valles Altos (2 200 a 2 600 m de altitud), es generalizado en la actualidad; su inicio data de 1986, cuando se identificaron ventajas a este tipo de híbridos: facilitan la producción de semillas; se requieren menor número de lotes de progenitores para llegar a la obtención de semilla certificada; el control de la calidad genética es accesible al poderse desmezclar fácilmente la línea progenitora macho, así Rowe y Andrew (1964) mencionan en general que poblaciones homogéneas de maíz (líneas puras y cruzas simples) muestran menor estabilidad que poblaciones heterogéneas (cruzas dobles y trilineales).
En el periodo de 1943 a 1985, los híbridos de cruza doble de progenitores de baja endogamia, fue el tipo de conformación de híbridos más utilizado en México, con base en un objetivo relacionado con la intención de que tuviesen mayor adaptabilidad; sin embargo, en general mostraban baja heterosis, dificultades para la producción de semilla, más esfuerzo para su producción en ciclos de cultivo, y mayores costos de producción para la obtención del híbrido final (Espinosa et al., 1986). Los híbridos de cruza simple que se han generado para la zona de producción de Valles Altos no han tenido el potencial productivo esperado, pero tampoco se cuenta con líneas hembras homocigóticas altamente rendidoras; estos híbridos presentan alta interacción genotipo*ambiente (GE). Estas cruzas simples son la clave para la obtención de híbridos trilineales, ya que la cruza simple que participa como hembra es de alta productividad (Espinosa et al., 1998; Sierra et al., 2006; Márquez, 2009).
En los programas de mejoramiento genético, es importante definir cuáles son los genotipos sobresalientes en cuanto a rendimiento, adaptación y estabilidad, evaluando estas propiedades en multi-ambientes para recomendar su uso de manera comercial (Crossa et al., 2006). La adaptación en sentido amplio, se refiere al mejor comportamiento relativo de un genotipo en la mayoría de los ambientes de prueba, mientras que, en sentido específico, la muestra el genotipo con mejor comportamiento relativo en un determinado ambiente de prueba (Cooper et al., 1999; Fuentes et al., 2005). Al respecto, Hanson (1970), define al genotipo estable como aquel que tiene la variabilidad mínima posible, cuando se desarrolla en diferentes ambientes.
El estudio de la interacción genotipo*ambiente (GE) en el mejoramiento genético vegetal es muy importante, pues esta es el resultado de la respuesta de cada genotipo frente a las variaciones ambientales (Crossa et al., 2006). Para lograr mayores avances en el mejoramiento genético de una especie, es necesario establecer correctamente las metodologías a utilizar para la evaluación de la interacción genotipo*ambiente (GE), para estimar con precisión la respuesta diferencial de los genotipos a través de los ambientes de prueba.
Entre las diversas metodologías para el estudio de la estabilidad en cultivos, destaca el modelo de Eberhart y Russell (1966), modificado del método de Finlay y Wilkinson (1963), utilizando la media aritmética de los datos reales y señalando que el coeficiente de la regresión de los efectos ambientales sobre los efectos fenotípicos puede utilizarse como estimador para medir la respuesta de cada cultivar a los índices ambientales, y la estabilidad de producción es posible medirla por la magnitud de la desviación a partir de la regresión lineal; es decir, por medio del cuadrado medio de la desviación de regresión. En este modelo una variedad con media alta, coeficiente de regresión βi=1 y desviación no significativamente diferente de cero (
Por otra parte, en los métodos multivariados, el modelo de efectos principales aditivos e interacción multiplicativa (AMMI), consiste esencialmente en combinar las técnicas del análisis de varianza y el análisis de componentes principales (ACP) en un sólo modelo, aquí el análisis de varianza permite estudiar los efectos principales de genotipos y ambientes, en tanto que, la interacción genotipo x ambiente (IGA) se trata de forma multivariada mediante el ACP, donde se realiza una reparametrización del modelo de regresión para mejorar la interpretación de la interacción (Zobel, 1988).
Para interpretar los resultados obtenidos del análisis AMMI, de acuerdo con Yan et al. (2000), es utilizar los efectos combinados de genotipos (G) y de la interacción genotipo por ambiente (IGE) en la evaluación del rendimiento, obteniéndose los gráficos denominados GGE (genotipo G + interacción genotipo*ambiente GE) biplot, que facilitan la identificación visual de los genotipos y los ambientes de evaluación. Generalmente, los gráficos GGE biplot se confeccionan utilizando los dos primeros componentes principales (CP1 y CP2). Así, el genotipo que está en el vértice es el que responde mejor en los ambientes de evaluación (Yan et al., 2001).
Partiendo de lo mencionado anteriormente, el objetivo de este trabajo fue identificar híbridos trilineales de maíz que presenten mejor estabilidad del rendimiento de grano, utilizando dos modelos para evaluar la estabilidad parámetros de estabilidad de Eberhart y Russell y AMMI, así como definir al modelo que mejor describa la interacción genotipo*ambiente. Con base en los resultados, se pretende recomendar la liberación comercial posterior en el caso de los mejores híbridos trilineales identificados en este trabajo.
Materiales y métodos
Este trabajo se realizó en el ciclo primavera-verano 2012, en dos localidades; la primera fue la FESC-UNAM (CuautitlánUNAM), municipio de Cuautitlán Izcalli, Estado de México, a una altitud de 2 274 m; la segunda localidad fue en el predio Santa Lucía de Prías, del Campo Experimental Valle de México (CEVAMEX), del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), municipio de Texcoco, Estado de México, a una altitud de 2 240 m. En ambas localidades se sembraron dos experimentos en dos fechas de siembra. En Cuautitlán-UNAM la primer fecha de siembra fue el 21 de mayo de 2012, y segunda el 1 de junio de 2012, mientras que en el CEVAMEX-INIFAP, la primer fecha de siembra fue el 18 de mayo de 2012 y la segunda el 29 de mayo de 2012.
Se utilizaron los híbridos trilineales Puma 1183 AEC1 y Puma 1183 AEC2 de la UNAM, H-53 (Espinosa et al., 2012a) y H-57 (Espinosa et al., 2012b) del INIFAP, todos en su versión androestéril (AE) y fértil (F) en dos densidades de población, 55 000 y 70 000 plantas por hectárea. Densidades que se recomiendan por el las características fenotípicas de las plantas de genotipos evaluados por cuestión de competencia. Los híbridos trilineales, las dos fechas de siembra y las dos densidades de población en las dos localidades se individualizaron para conformar los ambientes, generándose así ocho ambientes (dos localidades x dos fechas de siembra x dos densidades de población).
En todos los ambientes, en las diferentes fechas de siembra se aplicó un riego después de la siembra. Para el control de malezas se utilizó Gesaprim (Atrazina) a una dosis de 2 kg ha-1 y Hierbamina (2-4D amina) a una dosis de 2 L ha-1. La cosecha se realizó los días 10 y 17 de diciembre de 2012, en Cuautitlán Izcalli y Santa Lucía, respectivamente. En ambas localidades se utilizó un diseño experimental de bloques completos al azar con tres repeticiones, donde la parcela experimental fue de un surco de 5 m de largo y 0.8 m entre surcos, la parcela útil tuvo un área de 4 m2. Se utilizó un diseño factorial para el análisis estadístico del rendimiento, donde los factores de variación para los híbridos trilineales fueron ambientes, genotipos y las interacciones entre los factores de variación. La comparación de medias se hizo con la prueba de Tukey al 0.05 de probabilidad. Para evaluar la interacción genotipo*ambiente se utilizó como variable el rendimiento de los híbridos trilineales.
Para evaluar la respuesta de los ambientes, los genotipos y la IGA se empleó el modelo de efectos principales aditivos e interacción multiplicativa (AMMI), se realizó el análisis de varianza para determinar los efectos de genotipos, de ambientes, y de sus interacciones. Se consideraron los dos primeros componentes principales para la explicación de la interacción genotipo*ambiente. La representación gráfica se obtuvo por un Biplot, el cual mostró el patrón producto de la interacción genotipos*ambientes.
Para el análisis se emplearon las rutinas de programación AMMI descritas por Vargas y Crossa (2000), utilizando el siguiente modelo matemático:
Donde: Yij= rendimiento del i-ésimo genotipo en el j-ésimo ambiente; µ= media general; gi= efecto del i-ésimo genotipo; ej= efecto del j-ésimo ambiente; λk= raíz cuadrada del vector característico del k-ésimo eje del ACP; αik= calificación del ACP para el k-ésimo eje del i-ésimo genotipo; γjk= calificación del ACP para el k-ésimo eje del j-ésimo ambiente; Eij= valor del error.
Se obtuvieron los parámetros de estabilidad de Eberhart y
Russell (1966) para poder medir la interacción genotipo x ambiente, y se
utilizó la clasificación de la adaptabilidad de las variedades propuesta por Carballo y Márquez (1970), donde
bi es el coeficiente de regresión y
Donde: Yij= promedio del genotipo i en el ambiente j; μi=promedio del genotipo i en todos los ambientes; βi= coeficiente de regresión que mide la respuesta del genotipo i al variar los ambientes; Ij=índice ambiental del ambiente j-ésimo, que se calcula como la desviación del promedio de los genotipos en un ambiente dado a partir del promedio general; δij= desviación de la regresión.
Los datos utilizados en ésta metodología se analizaron aplicándoles un algoritmo computacional para obtener los indicadores de estabilidad de Mastache y Martínez (1996).
Todos los análisis estadísticos se realizaron mediante el programa SAS versión 9.0 (SAS Institute, 2002).
Resultados y discusión
El análisis de varianza combinado del AMMI (Cuadro 1) detectó diferencias altamente significativas entre ambientes y entre genotipos para rendimiento, mientras que la interacción genotipo*ambiente resultó no significativa. El coeficiente de variación fue 12.7%, considerado un valor aceptable en cuanto a la condición del experimento, por los valores obtenidos de r-cuadrada y por las aplicación del diseño experimental. La diferencia entre ambientes para rendimiento indica que las condiciones ambientales y sus efectos sobre los genotipos fueron diferentes en todos los ambientes de prueba. La diferencia en rendimiento entre genotipos fue el resultado de la divergencia genética entre los híbridos evaluados; es decir, el rendimiento de los genotipos no fue el mismo porque éstos tuvieron un origen genético diferente, debido a que los progenitores se derivaron de diferentes poblaciones de maíz. La ausencia de significancia de la interacción genotipo*ambiente (IGE) para rendimiento, el cual es un carácter de altamente cuantitativo, y por lo tanto muy influenciado por las condiciones ambientales, indica que estadísticamente no hubo una respuesta diferencial del rendimiento de los genotipos a través de los diferentes ambientes de prueba.
Cuadro 1 Análisis de varianza AMMI para el rendimiento de cuatro híbridos evaluados en ocho ambientes FESC-UNAM y Santa Lucía INIFAP, primavera - verano 2012.
| Fuente de Variación | G.L | SC | CM |
| Ambientes (a) | 7 | 18081507.04 | 2583072.43 ** |
| Genotipos (g) | 3 | 54167006.91 | 18055668.97 ** |
| Interacción (gxa) | 21 | 14316864.75 | 681755.5 |
| CP1 | 9 | 8885107.4 | 987234.15 |
| CP2 | 7 | 3944506.8 | 563500.97 |
| Error | 62 | 39381766.9 | 635189.8 |
| Total Corregido | 95 | 127620558 | |
| CV (%) | 12.7 | ||
| Promedio | 6293.4 |
*, **: Significancia (p≤ 0.05, 0.01); GL= grados de libertad; SC= suma de cuadrados; CM= cuadrados medios; CV= coeficiente de variación; CP1 y CP2= componente principal.
Con respecto a la cantidad de la suma de cuadrados total, el efecto de ambientes contribuyó en 14.2%, la interacción genotipos x ambientes en 11.2%, y 42.4% del total correspondió al efecto de genotipos. Lo anterior indica que los genotipos, en su conjunto, contribuyeron en mayor proporción a la variación del rendimiento en comparación con los factores ambientales y de la interacción genotipo*ambiente, aun cuando hubo diferencia altamente significativa entre ambientes. Este resultado contrasta con el obtenido en otras investigaciones, en las que el factor ambiental y de interacción genotipo*ambiente fueron superiores a los efectos de genotipos (Alejos et al., 2006; Palemón et al., 2012).
Los genotipos que tuvieron el mayor rendimiento fueron H-53 y H-57 en todos los ambientes de prueba. Los ambientes con los mayores rendimientos fueron los de Cuautitlán en la segunda fecha de siembra (1 de junio de 2012), con densidad de población de 50 000 plantas por hectárea y Cuautitlán en la segunda fecha de siembra (01 de junio de 2012), con densidad de población de 70 000 plantas por hectárea. Resultado de las condiciones ambientales benéficas en la localidad de Cuautitlán, de precipitaciones y las propiedades del suelo para retener humedad aprovechable para la planta. Los genotipos que obtuvieron valores absolutos más bajos del CP1, es decir, que interaccionaron menos con el ambiente, fueron, H-57, con 14.72, y Puma 1183 AEC2 con -17.93, que al presentar los valores del CP1 más cercanos a cero, se consideraron los más estables a través de ambientes (Medina et al., 2002; Alejos et al., 2006; Palemón et al., 2012).
Los ambientes con mejor comportamiento (no podemos decir que un ambiente es estable, es incorrecto por eso se agrega que tiene buen comportamiento) con respecto a los valores absolutos del CP1 fueron: Cuautitlán en la segunda fecha de siembra, con densidad de población de 70 000 plantas por hectárea (A5), con -0.28; Santa Lucía en la primer fecha de siembra, con densidad de población de 50 000 plantas por hectárea (A3), con 5.12; Santa Lucía en la segunda fecha de siembra, con densidad de población de plantas por hectárea (A7), con 9.15, y Cuautitlán en la primer fecha de siembra, con densidad de población de plantas por hectárea (A2), con -9.3 (Cuadro 2).
Cuadro 2 Rendimiento medio en kg ha-1 de cuatro genotipos de maíz evaluados en ocho ambientes y valores de los dos primeros componentes principales para genotipos y ambientes. FESC-UNAM y Santa Lucía INIFAP, primavera - verano 2012.
| Genotipo | Ambiente | Media (kg ha-1) | CP1 | CP1 | |||||||
| Cuautitlán | Cuautitlán | Santa Lucía | Santa Lucía | Cuautitlán | Cuautitlán | Santa Lucía | Santa Lucía | ||||
| F1 D1 | F1 D2 | F1 D1 | F1 D2 | F2 D1 | F2 D2 | F2 D1 | F2 D2 | ||||
| A1 | A2 | A3 | A4 | A5 | A6 | A7 | A8 | ||||
| Puma 1183 | 5624.5 | 5709.6 | 4880 | 4723.9 | 6007.6 | 5859.7 | 5147.4 | 5090.3 | 5380.4 | -22.66 | -16.4 |
| AEC1 | |||||||||||
| Puma 1183 | 5793.5 | 6384.4 | 5146.2 | 5698.1 | 6372.3 | 7543.1 | 5727.8 | 6191.4 | 6107.1 | -17.93 | 12.9 |
| AEC2 | |||||||||||
| H-53 | 6128.9 | 7409.4 | 7180.9 | 7714.6 | 7634.2 | 7507.4 | 7775.3 | 8156.6 | 7469.6 | 25.87 | -17.03 |
| H-57 | 5118.6 | 6091.8 | 5391 | 6785.5 | 6737.8 | 7356.2 | 5937.4 | 6313.9 | 6216.5 | 14.72 | 20.5 |
| Media (kg ha-1) | 5728.7 | 6398.8 | 5649.5 | 6230.5 | 6766.1 | 6971.35 | 6147 | 6438.1 | |||
| CP1 | -25.8 | -9.3 | 5.12 | 21.4 | -0.28 | -14.3 | 9.15 | 14.02 | |||
| CP2 | -10.6 | -3.1 | -14.3 | 11.1 | 8.8 | 22.2 | -10.64 | -3.52 | |||
El híbrido H-53 mostró un rendimiento promedio superior a la media, mientras que los otros tres híbridos presentaron rendimientos menores a la media (Figura 1). El híbrido Puma 1183 AEC1 mostró el rendimiento promedio más bajo. Los híbridos con valores bajos en el CP1 y que interaccionan menos con el ambiente fueron, H-57 y Puma 1183 AEC2, mientras que los genotipos con mayor interacción con el ambiente por sus valores altos de CP1 fueron, H-53 y Puma AEC1. De manera más precisa, se logró identificar a los ambientes A5, A3, A7 y A2 como los de mejor comportamiento, por sus valores de rendimiento y del CP1, por estar más cercanos a cero; es decir, los que mostraron la menor variación entre ellos, y considerando que influyó en cada ambiente de la densidad de población, la localidad y la fecha de siembra, en los ambientes A5, A3, A7 y A2 se presentaron las mejores condiciones para obtener los mejores rendimientos.
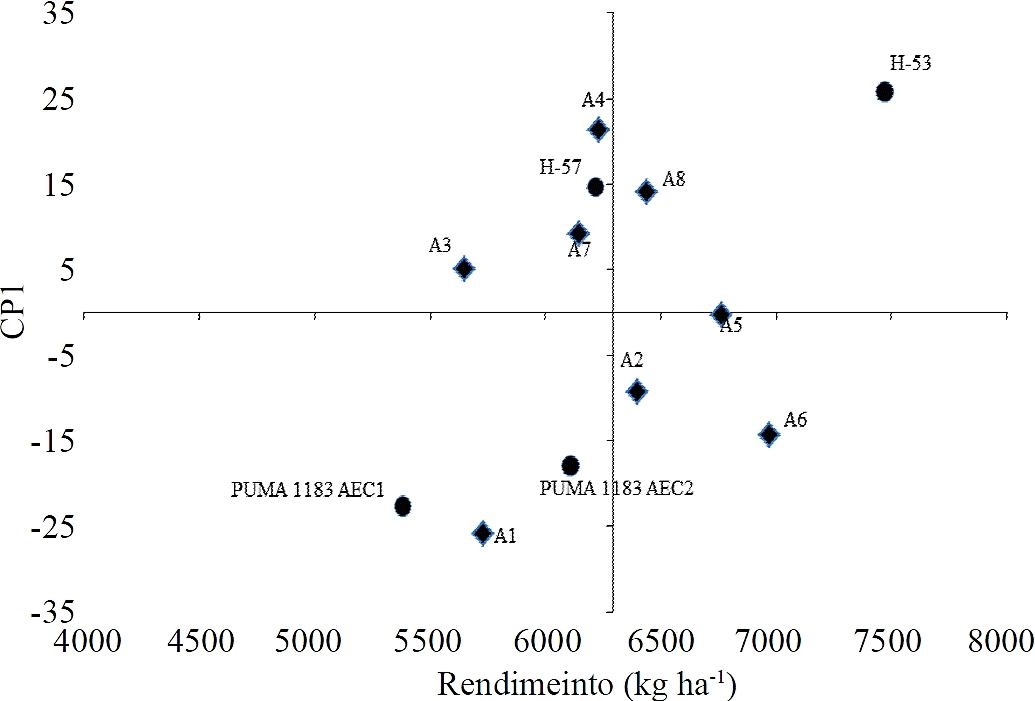
Los círculos representan los genotipos y los rombos a los ambientes.
Figura 1 Biplot del CP1 en función del rendimiento promedio de cuatro híbridos trilineales de maíz evaluados en ocho ambientes en la FESC-UNAM y Santa Lucía INIFAP, primavera - verano 2012.
En el análisis AMMI, el primer componente principal (CP1) explicó 60.5% de la suma de cuadrados, y el segundo componente principal (CP2) observó 26.8% de la suma de cuadrados, por lo que entre los dos componentes principales describieron 87.3% el efecto de la interacción genotipo x ambiente. Por otro lado, con respecto a los ambientes, Yan et al. (2000) señala que los ambientes cuyos ángulos sean menores a 90° clasificarán a los genotipos de manera semejante. En este caso, en particular se formaron dos grupos de ambientes con comportamientos similares; en el primer grupo se encontraron los ambientes, A1, A2, A5 y A6; en el segundo se ubicaron los ambientes A3, A4, A7 y A8, siendo los ambientes A1, A4 y A6, los que mejor discriminaron a los genotipos.
Por su parte, Eeuwijk (2006) indica que un ángulo menor de 90° o mayor de 270° entre el vector de un cultivar y un vector de sitio indican que el cultivar tiene una respuesta positiva al sitio de prueba. Una respuesta negativa del cultivar es indicada con ángulos mayores de 90° y menores de 270°. Lo anterior coincide con los resultados del gráfico biplot (Figura 2), donde H-53 se respondió mejor en los ambientes A7, A8, A3 y A4, mientras que H-57 mostró una mejor respuesta en los ambientes A4, A8 y A6. Por su parte, Puma 1183 AEC2 mejoró en los ambientes A5, A6, y A2, mientras que Puma 1183 AEC1 presentó mejor respuesta en los ambientes A5, A2 y A1. Así, los genotipos que se encontraron más cerca al origen en el gráfico biplot, fueron los que interaccionan menos con el ambiente y por lo tanto los más estables; por el contrario, los más alejados mostraron mayor variación en su comportamiento (Yan et al., 2000). Estos resultados muestran que los genotipos más estables fueron Puma 1183 AEC2 y H-57, mientras que los genotipos que más interaccionaron con el ambiente fueron H-53 y Puma 1183 AEC1.
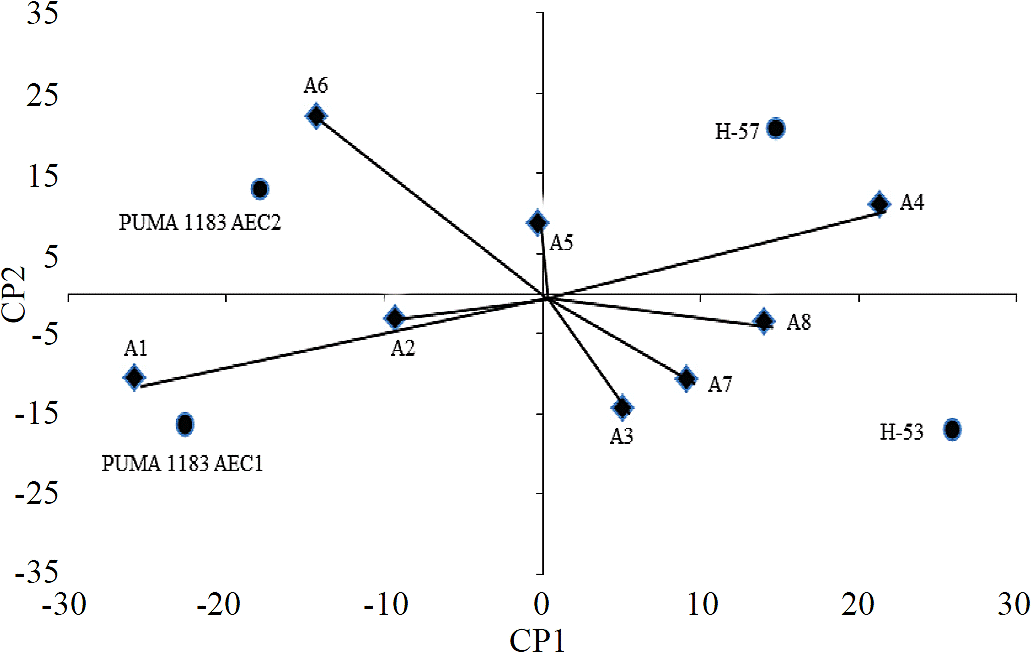
Los puntos representan a los híbridos trilineales y los vectores representan a los ambientes.
Figura 2 Biplot AMMI para cuatro híbridos trilineales de maíz.
El análisis de varianza de los Parámetros de Estabilidad de Eberhart y Russell (1966), para rendimiento, no mostró significancia entre ambientes, entre genotipos, y para la interacción genotipo*ambiente. Esto indica que entre ambientes, las condiciones ambientales tuvieron un comportamiento similar, lo que causó un efecto no significativo en el rendimiento, mientras que para genotipos, aun cuando provinieron de diferente origen genético, su comportamiento promedio en todos los ambientes fue muy parecido. La interacción genotipo x ambiente, al no ser significativa indica que no hubo una respuesta diferencial del rendimiento de los genotipos a través de los diferentes ambientes de prueba.
De acuerdo con los parámetros de estabilidad estimados, el coeficiente de regresión (bi), desviación de regresión (
Cuadro 3 Rendimiento medio, coeficiente de regresión (bi), desviaciones de regresión (
| Genotipo | Rendimiento (kg ha-1) | bi |
|
| PUMA 1183 AEC1 | 5380 | 0.5286 | -213063.97 |
| PUMA 1183 AEC2 | 6107 | 1.3304 | -292066.93 |
| H-53 | 7438 | 0.7117 | -121018.54 |
| H-57 | 6217 | 1.4293 | -309803.83 |
*, **= significancia (p≤ 0.05, 0.01); SC= suma de cuadrados; CM= cuadrados medios; CV= coeficiente de variación.
Para definir la mejor metodología para caracterizar a los genotipos más estables y a los ambientes donde los genotipos presentan una mejor respuesta en el rendimiento, para esto, se compararon los valores del primer componente principal (CP1) correspondientes a la metodología AMMI, y los valores de los índices ambientales correspondientes a la metodología de los parámetros de estabilidad. En el Cuadro 4 se muestra que el modelo AMMI identificó como ambientes con mejor respuesta por parte de los genotipos para rendimiento a, A5, A3, A7 y A2, por tener valores del CP1 más cercanos a cero, mientras que los ambientes con mala respuesta de los genotipos en el rendimiento fueron, A1, A4, A6 y A8, por tener los valores del CP1 más alejados de cero. El modelo de los parámetros de estabilidad identificó como los mejores ambientes para rendimiento a, A6, A5, A8 y A2, por los valores de sus índices ambientales positivos; es decir, superiores a la media general, y los ambientes con mal comportamiento fueron, A1, A3, A7 y A4, por tener índices ambientales negativos; es decir, menores a la media general. La correspondencia que existió entre ambos modelos fue de 50%, ya que coincidieron ambos modelos en determinar a dos ambientes estables y dos ambientes inestables, de los ocho ambientes de prueba.
Cuadro 4 Comportamiento medio del rendimiento en los ocho ambientes evaluados, componente principal uno (CP1) e índices ambientales. FESC UNAM y Santa Lucía INIFAP, primavera - verano 2012.
| Ambientes | Rendimiento (kg ha-1) | CP1 | Índice ambiental |
| A1 | 5728 | -25.8 | -619.4 |
| A2 | 6399 | -9.3 | 113.2 |
| A3 | 5649 | 5.1 | -636.1 |
| A4 | 6230 | 21.4 | -55.1 |
| A5 | 6766 | -0.28 | 402.4 |
| A6 | 6971 | -14.3 | 781 |
| A7 | 6147 | 9.1 | -138.6 |
| A8 | 6438 | 14 | 152.5 |
*, **= significancia (p≤ 0.05, 0.01); GL= grados de libertad; SC= suma de cuadrados; CM= cuadrados medios; CV= coeficiente de variación; CP1 y CP2= componente principal.
En la comparación del modelo AMMI con el de Eberhart y Russell para identificar a los genotipos más estables (Cuadro 5), con el modelo AMMI los genotipos más estables fueron H-57 y Puma 1183 AEC2, por sus valores del CP1 más cercanos a cero. Con el modelo de los parámetros de estabilidad, todos los genotipos fueron estables. La correspondencia entre ambos modelo fue 50%, ya que sólo dos de los cuatro genotipos evaluados coincidieron como estables con ambos modelos, resultados similares a los de Córdova (1991), quien obtuvo 40% de coincidencia al utilizar ambas metodologías.
Cuadro 5 Respuesta del rendimiento de cuatro híbridos evaluados, componente principal uno (CP1) y coeficiente de regresión. FESC-UNAM y Santa Lucía INIFAP, primavera - verano 2012.
| Genotipo | Rendimiento (kg ha-1) | CP1 | bi |
| PUMA 1183 AEC1 | 5380 | -22.7 | 0.5286 |
| PUMA 1183 AEC2 | 6107 | -17.9 | 1.3304 |
| H-53 | 7438 | 25.9 | 0.7117 |
| H-57 | 6217 | 14.7 | 1.4293 |
*, **= significancia (p≤ 0.05, 0.01); GL= grados de libertad; SC= suma de cuadrados; CM= cuadrados medios; CV= coeficiente de variación; CP1 y CP2= componente principal.
Conclusiones
Los híbridos trilineales H-57 y Puma 1183 AEC2 tuvieron la mejor estabilidad y el mayor rendimiento. Los ambientes con mejor comportamiento de los genotipos en el rendimiento de los híbridos trilineales fueron, Cuautitlán en la segunda fecha de siembra, con densidad de siembra de 50 000 plantas por hectárea (A5); Santa Lucía en la primer fecha de siembra, con densidad de siembra de 50 000 plantas por hectárea (A3); Santa Lucía en la fecha de siembra 2, con densidad de siembra de 50 000 plantas plantas por hectárea (A7) y, Cuautitlán en la primer fecha de siembra, con densidad de siembra de 70 000 plantas por hectárea (A2).
El mejor modelo para analizar la interacción genotipo*ambiente fue el modelo AMMI, en comparación con los parámetros de estabilidad de Eberhart y Russell. Lo anterior se debe a que la interacción genotipo*ambiente conviene analizarla como un efecto multiplicativo y no sólo como un efecto lineal, además de que al hacer uso del Biplot, se explica fácilmente la interacción genotipo*ambiente, utilizando los valores de los componentes principales y del rendimiento de los genotipos.

