











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
A habitat is the physical space where a species naturally occurs, and includes all the physical and biological characteristics (environmental factors) necessary to cover its basic needs. However, spatial and temporal variation in habitat conditions generate a selective pressure resulting in the use of different habitat components (Cody, 1985; Krauman, 1999) and this will vary according to the availability, richness, or utility of any resources that offer an advantage towards satisfying the requirements of a species (Hutton, 1985; Johnson, 1980).
Quantifying the abiotic and biotic resources that determine the presence of species in a habitat is complex, because the selection of resources by one species also depends upon the resource-use by the predators, co-predators, and prey species that occur there (McLoughlin et al., 2010; Slagsvold, 1980). In addition, the impact and changes that result from human activity on an ecosystem play an important role in habitat selection and resource-use by many animals (Balestrieri et al., 2009; McLoughlin et al., 2010).
The study and management of large carnivores has proven to be one of the best strategies for conserving their wider ecosystem because of their large home-ranges and extensive areas of activity. Therefore, in order to protect these predators, expansive areas of habitat are protected along with all those species that inhabit the same area. One such carnivore is the jaguar (Panthera onca), which is considered both a keystone and umbrella species, as well as an indicator of the proper functioning of its environment (Manterola et al., 2011; Miller & Rabinowitz, 2002; Núñez et al., 2002). Therefore, understanding the jaguar’s use and selection of resources is of high interest in terms of conserving this species and the ecosystems where it occurs. Despite the importance of the jaguar, its populations have been declining rapidly due to habitat loss in parts of Mexico and Central America (Caso et al., 2008; De la Torre et al., 2017) and it is considered to be a species in danger of extinction (Armella & Yáñez, 2011).
Current knowledge of resource selection by the jaguar suggests that it prefers forested areas near water and avoids disturbed or modified areas (Chávez, 2010; Conde et al., 2010; Cullen, 2006). The habitat components that play an important role in jaguar distribution include vegetation type and habitat continuity (De la Torre et al., 2017; Hidalgo-Mihart et al., 2017), land cover, water availability, prey (Cullen et al., 2013; Scognamillo et al., 2003), and anthropogenic activities (Núñez et al., 2002; Romeu, 1996; Sanderson et al., 2002; Zarza et al., 2007).
Current information about jaguar resource-use only covers a limited part of its geographical range. Studies from Mexico include one from the southern Yucatán Peninsula (Chávez, 2010), and another from the northern Yucatán Peninsula on their population density in the Ría Lagartos Reserve and surrounding areas including the Zapotal, Yum Balam, and Dzilam Reserves (Faller et al., 2007). The Yucatán Peninsula is considered a priority study area for jaguar (Ceballos et al., 2005; Sanderson et al., 2002) because it contains original native vegetation types that give an extensive continuity of habitat and maintain communication with the Maya Forest (De la Torre et al., 2017; Hidalgo-Mihart et al., 2017). Therefore, the aim of this study was to analyze resource-use by jaguars, as well as activity patterns, and information about associated species and environmental variables that influence their occurrence in the tropical forest in the north-eastern Yucatán Peninsula.
Materials and methods
The study was conducted in El Edén Ecological Reserve (EER) and surrounding areas in the municipality of Lázaro Cárdenas, Quintana Roo, in the north-east Yucatán Peninsula, Mexico (21°36’00”-20°34’00” N, 87°06’00”-87°45’00” W; Fig. 1a-e). The EER covers an area of 3,077 ha in the Yalahau region in the northernmost tropical forests of North America, at an elevation of 5-10 m asl (Gómez-Pompa et al., 2003, 2010; Lazcano-Barrero et al., 1992). This encompasses the Yum Balam protected area, which has extensive areas of medium-stature deciduous forest (MSF) and secondary forest or acahual (Navarro et al., 2007). MSF is characterized by trees reaching heights of 10-15 m. The most common species are Manilkara zapota, Lysiloma latisiliqua, Metopium brownei, and Thrinax radiata palm. Acahual is derived from MSF and characterized by trees of 7-10 m in height, typically M. brownei, Bursera simaruba, and Swartzia cubensis. There is also some flooded forest or tintal, dominated by Haematoxylon campechianum, Erythroxylon confusum, and Byrsonima bucidaefolia, plus small fragments of Annonaceae forest and savanna (Schultz, 2003) (Fig. 1).
Figure 1. The location of camera traps (large black dots) plotted according to year of study (a, 2008; b, 2010; c, 2011; d, 2012) showing major vegetation types (secondary forest; shaded with small dots on a white background), mixed farming (shaded in grey), medium-stature forest (shaded in white), and sink holes (small black dots), and e, the location of the study area at El Edén Ecological Reserve, Mexico (21°36’00”-20°34’00” N, 87°06’00”-87°45’00” W).
The study was conducted during July-September 2008, October-December 2010, May-July 2011, and August-December 2012 using camera traps (Cuddeback Expert, Capture, Capture IR, Moultrie, and Wildview). Camera placement followed the CENJAGUAR design (Chávez et al., 2007) with 2 or 3 sampling stations in 9 study plots each measuring 9 km2. The location (longitude and latitude) of each camera station was taken with a Global Positioning System (GPS-Garmin eTrex) and entered into ArcView 8 Geographical Information System (GIS).
The cameras were sited on forest paths or trails, firebreaks, and roads (about 6 m wide) at a distance of 1-3 km from one another. In 2008, there were 9 camera stations in MSF and 18 in acahual. In 2010, there were 10 in MSF and 14 in acahual, in 2011, 10 in MSF and 12 in acahual, and in 2012, 12 in MSF and 22 in acahual (Fig. 1a-d). Despite the variation in the number of cameras over the years, they occupied the same area. The cameras operated 24 h d and were checked and images downloaded once every 15 d.
Photographs were grouped by trap site to allow comparison of trends in the spatial distribution of individual felids, which were identified by their unique coat patterns. Photographic records were considered to be independent events when 1) the same individual was recorded at intervals of more than 30 min apart, 2) it was possible to distinguish between different individuals of the same species in consecutive photos, 3) a photograph showed several different individuals that could be clearly identified, and 4) where members of the same species could not be identified as being different individuals in consecutive photos, a new event was only recorded once in every 3 h (Ávila-Nájera et al., 2016).
Terrestrial or semi-terrestrial mammals and some birds of terrestrial habits that appeared in the photographs were considered to be associated species, and their activity was recorded. For some analyses they were grouped by taxon and weight into 5 classes: large mammals (8-26 kg), medium-sized mammals (3-8 kg), small mammals (1-3 kg), large birds (2-6 kg), and small birds (< 2 kg) (Davis et al., 2010). In addition, photographic records of human activity were included.
Photographs were grouped according to the time of the record: nocturnal (20:00-06:00), crepuscular (06:00-08:00, 18:00-20:00), and diurnal 08:00-18:00 (Monroy-Vilchis et al., 2011), and results were pooled into periods of 2 h (starting at 00:00) in order to detect peak times of activity. All species recorded by the camera traps were classified according to their activity pattern using the following rules: diurnal when ˂ 15% of observations were at night, mainly diurnal when 15-35% of observations at night, nocturnal when > 85% of observations were at night, and mainly nocturnal when 65-85% of observations were at night.
Traps were classified according to vegetation (MSF and acahual) and trail type (whether the camera was by a forest trail, firebreak, or dirt road), and the frequency of capture in the photos was used to determine jaguar activity patterns in each habitat.
Pearson’s chi-squared test (Chernoff & Lehmann, 1954; Siegel & Castellan, 1988) was used to test for differences in jaguar use of vegetation, trail type, and activity pattern between the study years. The test compared the observed activity (from photographs) and expected activity (percentage of cameras sited by each trail type), and the activity time of jaguars (from photographs) and an expected activity pattern of 10/24 h for diurnal/nocturnal and 4/24 h for crepuscular activity. A Monte Carlo simulation was used to estimate the significance levels (p-value; Hope, 1968).
Overall differences in jaguars’ use of resources and activity patterns were tested for
evidence of selection of type of vegetation (MSF or acahual), trail type (forest
trail, firebreak or dirt road), and time of activity using the equation:
H0: p
i
=p0, where the real proportion is
pi for the i-th category,
p0 is the expected proportion according to the null
hypothesis, at the observed proportion for the i-th category
We used 2 × 2 contingency tables to test the null hypothesis that different species’ records in the traps were independent of each other (at α = 0.05; Ludwig & Reynolds, 1988) and using Yates’ correction factor:
where a = number of sampling units where both species occur, b = number of sampling units
where a occurs, but not b, c = number of sampling units where b occurs, but not a, d
= number of sampling units where none of the 2 species were found, N= total number
of sampling units (N = a + b + c + d); m = a + b; n= c + d; r = a + c; s = b + d.
Therefore, if
To test for the overall effect of environmental and resources on jaguar occurrence we used Generalized Linear Models (GLM) (Sileshi, 2008), calculated using the glm.nb function in R. We assumed a negative binomial distribution and considered camera stations 1.5 km or farther apart to be independent sampling units. We also assumed that all vegetation and path types were equally accessible to jaguar and their prey and co-predators recorded in the traps (Table 1).
Table 1 List of variables selected according to the Akaike information criterion (AIC) (Burnham & Anderson, 2002) and incorporated into 18 models for each year and 18 models using all the data from the 4 years (90 models in total).
| Model name | Variables selected using AIC |
| m0 | Null |
| m2 | Co-predator activity pattern |
| m4 | Associated species records, adjusted for the presence of humans |
| m6 | Associated species, co-predator records and humans |
| m8 | Associated species records with activity pattern, adjusted for the presence of humans |
| m10 | Activity pattern of associated species and co-predator records, adjusted for the presence of humans |
| m12 | Environmental factors (distance to body water, vegetation types, trail type, longitude, latitude) |
| m14 | Environmental factors and associated species records, adjusted for the presence of humans |
| m16 | Associated species records by taxa and weight group |
| m18 | Associated species records by taxa and weight group and environmental factors |
The following variables were used in the GLMs: vegetation type, horizontal and vertical plant cover, path type, potential prey species, co-predators (puma and ocelot), human activity, activity patterns of potential prey and co-predators, distance to the nearest water body and town, latitude, and longitude. Horizontal plant cover is the percentage of vegetation capable of hiding a subject 15 m away, and vertical plant cover is the visibility through the tree canopy (Griffith & Youtie, 1988). Arc Map 10.3 (GIS) was used to determine the distance to the nearest water body and town. Significance was determined using an asymptotic t-test at α = 0.05 level, with Akaike’s Information Criterion (AIC) to select the best model (Burnham & Anderson, 2002), since this favors simpler models and acts to reduce the number of parameters.
Results
A total of 181 independent photographic records of jaguar were captured, and a total 12 individual jaguars were identified during the study. There were 96 photographs taken in 2008, 15 in 2010, 12 in 2011, and 58 in 2012, with 7, 5, 5, and 12 individual jaguars identified in each of the study years, respectively.
In some years, there were differences in the observed and predicted frequencies of jaguars in each vegetation and trail type, and by activity pattern (Table 2). Evidence of selective resource-use include significant results for vegetation and trail type in 2008 and 2011 (all at p < 0.001) and activity pattern in 2011 (p < 0.001) and 2012 (p = 0.01) (Table 2).
Table 2 Results of x2 Pearson tests with Monte Carlo estimate for selection vegetation type, trail type, and activity pattern (nocturnal, diurnal, crepuscular) by year on the observed frequency of jaguar records from camera traps in El Edén Ecological Reserve in Quintana Roo, Mexico.
| Year of study | ||||||||
| Variable | 2008 | 2010 | 2011 | 2012 | ||||
| x2 | p | x2 | p | x 2 | p | x 2 | p | |
| Vegetation type | 21.9 | 0.00 | 0.31 | 0.86 | 0.159 | 0.00 | 1.76 | 0.41 |
| Trail type | 36.9 | 0.00 | 0.61 | 0.89 | 229 | 0.00 | 1.78 | 0.62 |
| Activity pattern | 1.40 | 0.71 | 5.96 | 0.11 | 177 | 0.00 | 11.3 | 0.01 |
Most jaguar records were from acahual (80%) with only 20% from MSF. However, the frequency of jaguar records by vegetation type varied depending upon the year of study. There were more records from acahual in 2008, 2010, and 2012 (95%, 62% and 75%, total jaguar records by year of study, respectively) and more from MSF in 2011 (71%). In addition, the observed frequency of records in acahual was higher than expected in 2008 and 2011, but lower than expected in MSF (Table 3).
Table 3 Test of whether jaguar resource-use varied in El Edén Ecological Reserve, Quintana Roo, Mexico according to availability testing for the effect of vegetation type, trail type, and activity pattern with expected values estimated from 10,000 bootstrap simulations (where the test outcome shows Po < Pe, Po > Pe or no difference between Po and Pe (=), with corresponding Bonferroni confidence intervals).
| Resource | Classification type | Year | Observed proportion (Po) |
Expected proportion (Pe) |
Confidence interval range |
Test outcome (Po < > or = Pe) |
| Vegetation | Medium-stature forest | 2008 2010 2011 2012 |
0.04 0.38 0.21 0.25 |
0.33 0.5 0.45 0.34 |
0.00-0.12 0.07-0.69 0.18-0.25 0.11-0.40 |
< = < = |
| Acahual | 2008 2010 2011 2012 |
0.95 0.61 0.78 0.75 |
0.66 0.5 0.55 0.65 |
0.83-1 0.30-0.92 0.74-0.81 0.59-0.88 |
> = > = |
|
| Camera site | Forest trail | 2008 2010 2011 2012 |
0.04 0.38 0.22 0.25 |
0.37 0.5 0.5 0.34 |
0.00-0.12 0.07-0.76 0.18-0.26 0.11-0.40 |
< = < = |
| Firebreak | 2008 2010 2011 2012 |
0.06 0.15 0.13 0.22 |
0.12 0.18 0.1 0.20 |
0.00-0.15 0.00-0.46 0.00-0.16 0.08-0.37 |
= = = = |
|
| Road | 2008 2010 2011 2012 |
0.88 0.46 0.64 0.52 |
0.5 0.31 0.4 0.16 |
76.19-0.98 0.15-0.84 0.59-0.69 0.09-0.36 |
> = > > |
|
| Activity pattern | Crepuscular | 2008 2010 2011 2012 |
0.22 0.23 0.18 0.30 |
0.44 0.41 0.41 0.16 |
0.35-0.69 0.00-0.53 0.15-0.22 0.15-0.47 |
= = < = |
| Diurnal | 2008 2010 2011 2012 |
0.42 0.46 0.63 0.22 |
0.41 0.16 0.41 0.41 |
0.26-0.58 0.07-0.84 0.58-0.67 0.08-0.37 |
= = > < |
|
| Nocturnal | 2008 2010 2011 2012 |
0.34 0.30 0.18 0.47 |
0.41 0.41 0.16 0.41 |
0.19-0.50 0.00-0.69 0.14-0.22 0.30-0.64 |
= = = = |
Jaguars were active both day and night within the study area, although there were differences in activity patterns between the years, with more diurnal records and fewer crepuscular records in 2011. Overall, almost half (43%) of the records were nocturnal (20:00-06:00) with the main peaks of activity occurring between 21:00-22:00 and 02:00-03:00. About a quarter (25.5%) of the records were crepuscular, with most activity occurring between 19:00-20:00 and almost a third (31.3%) of the records occurring during the day (08:00-18:00) with fewer observations between 10:00-11:00 and 13:00-14:00. In 2011, the observed frequently of diurnal records was higher than expected, whilst for crepuscular records it was lower than expected (Table 3). There was no difference in the expected and observed frequency of nocturnal records over the 4 years of study (Table 3).
Figure 2(a-d) shows the annual relative abundance of other species recorded in the camera traps, grouped according to their taxa and size class, and whether they are co-predators (Fig. 2a) or potential prey (Fig. 2 b-d).
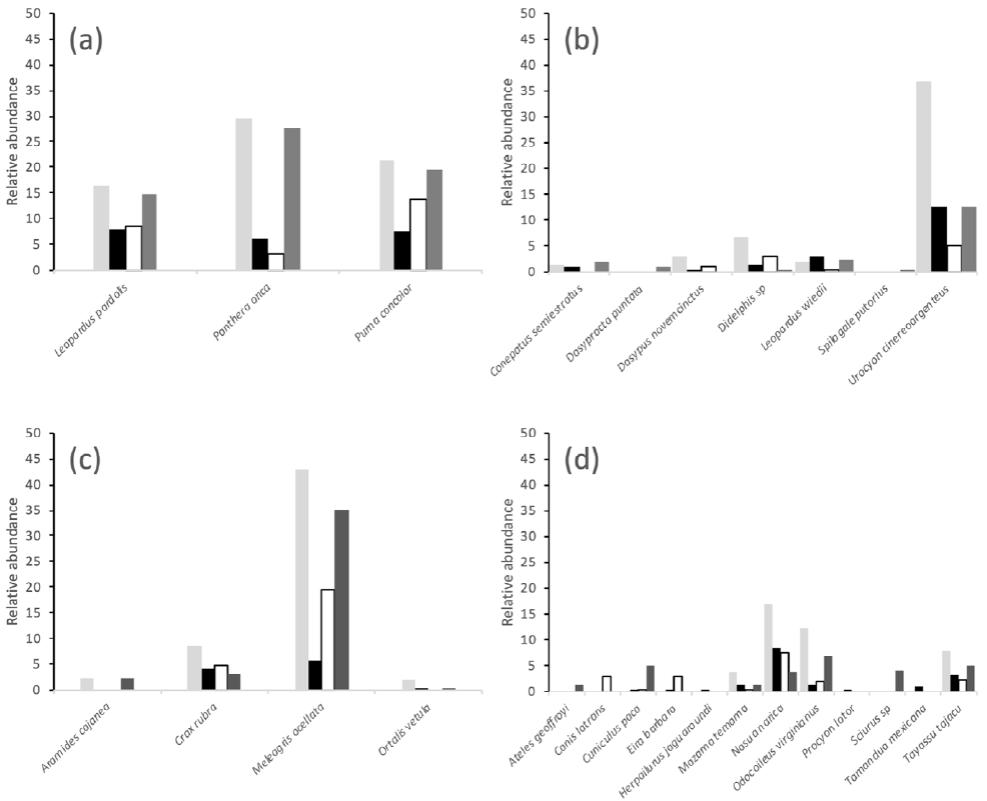
Figure 2. Annual relative abundance of the all animals recorded in the camera traps at El Edén Ecological Reserve, Quintana Roo, Mexico, grouped either by prey type: (a) co-predator, (b) large and medium mammals, (c) large and small birds, and (d) small mammals. Gaps in bars indicate that no observations were made for that species in a particular year (light grey (2008), black (2010), white (2011), dark grey (2012).
There was a significant association (x2 = 3.84, p < 0.05) between jaguar and some other species recorded in the camera traps including potential prey, Aramydes cajanea, Didelphis sp., Meleagris ocellata, Mazama temama, Nasua narica, Odocoileus virginianus, and Pecari tajacu, and the co-predator Leopardus pardalis. When camera trap results were grouped by vegetation type, there were some interesting associations between other species and jaguar. These varied depending upon the year of study. There was an association between jaguar records and traps sited in acahual in 2008 (x 2 = 3.84, p < 0.05), as well as for jaguar records and Dasypus novemcinctus and M. temama in 2010, L. pardalis in 2011 and for M. temama, O. virginianus, and Ortalis vetula in 2012. In MSF in 2008, there was a positive association between records of jaguar and Didelphis sp., and for N. narica in 2010, as well as for Leopardus wiedii, N. narica, P. tajacu, and Sciurus sp. in 2012, but there were no species associated with jaguar records from MSF in 2011.
Ninety GLMs were developed using the variables in Table 1 (18 per year and 18 for all 4 years combined). The effect of year was significant in all models, and GLMs based on environment (acahual, roads, and firebreaks) were the best at predicting jaguar presence. However, model 10 (292 AIC) (Table 4) had the lowest AIC value, and used the activity patterns of co-predators, associated species, and humans to predict the presence of jaguar.
Table 4 Model for prey species and co-predators by activity pattern and adjusted for human presence, after variable selection by AIC.
| Model 10 | Estimate | SE | z value | Pr (> |z|) |
| (Intercept) | 564 | 173 | 3.26 | 0.00 |
| Variable: year | -0.28 | 0.09 | -3.29 | 0.00 |
| Puma concolor diurnal activity | 0.42 | 0.12 | 3.60 | 0.00 |
| Leopardus pardalis crepuscular activity | 0.95 | 0.30 | 3.20 | 0.00 |
| Mazama temama crepuscular activity | -1.77 | 0.88 | -2.02 | 0.04 |
| Odocoileus virginianus diurnal activity | 0.98 | 0.25 | 3.99 | 0.00 |
| Urocyon cinereoargenteus diurnal activity | 0.32 | 0.12 | 2.60 | 0.01 |
| Aramides cajanea nocturnal activity | 0.37 | 0.17 | 2.10 | 0.04 |
| Crax rubra diurnal activity | -0.79 | 0.28 | -2.82 | 0.00 |
| Dasypus novemncinctus nocturnal activity | -3.21 | 0.97 | -3.31 | 0.00 |
Discussion
Although felids have been considered generalist species by many authors, some studies suggest they have specific habitat requirements and need certain components to be present in order to survive in an area (Chávez et al., 2010). In this study, there was evidence that jaguars selected and used resources according to their availability. However, they made higher than expected use of acahual and roads, although this varied by year, and their general presence within EER was defined by the abundance and activity patterns of associated species and co-predators. Jaguars changed their use of vegetation type depending upon the year of study, and this was particularly influenced by other factors such as the presence of humans or prey species. Secondary forest was a preferred habitat, and we believe that this is because it provides both horizontal and vertical vegetation cover, which are essential for jaguars’ permanence in a habitat (Conde et al., 2010).
Existing jaguar distribution data suggest that they inhabit many habitats (Zarza et al., 2007), favoring dense vegetation and areas that provide sufficient water and a diverse and abundant variety of prey (Manterola et al., 2011; Sanderson et al., 2002). They occur primarily in tropical rain forest, followed by tropical dry forest, and are at their lowest densities in arid environments and grasslands (Sanderson et al., 2002). Previous studies from the Yucatán Peninsula show that jaguars have a marked preference for areas with dense vegetation cover (Ceballos et al., 2005; Chávez et al., 2007). In EER, jaguars were recorded on roads, firebreaks, and forest trails, as reported by other authors (Maffei et al., 2004). Most records were from roads, and many authors suggest that jaguars use them to move along or for performing other activities such as scent marking. However, Conde et al. (2010) report a gender bias, with more records of male jaguars from roads, and this may in turn influence the gender of other road users (Maffei et al., 2011). In our study area there was very little traffic, making roads a relatively safe and undisturbed environment, often surrounded by dense vegetation cover, so that their open terrain would facilitate animal movement (Harmsen et al., 2010).
Most jaguar records in the EER were crepuscular or nocturnal, with 2 peaks in nocturnal activity, and crepuscular activity generally occurring around dusk. Similarly, Chávez et al. (2007) found that jaguars in the south of the Yucatán Peninsula (Calakmul) were mainly crepuscular or nocturnal, although peak activity occurred in the early morning. In other parts of their range, jaguars can be active 24 h d, although this varies with locality, and depends upon the availability and activity patterns of their prey (Carrillo, 2000; Scognamillo et al., 2003), and in some case they are mainly diurnal (Álvarez-Castañeda & Patton, 2000; Foster et al., 2013; Harmsen et al., 2009; Maffei et al., 2004, 2011; Rabinowitz & Nottingham, 1986). In the Chaco region of Bolivia, jaguar had 2 peak periods of activity, 03:00-06:00 and 17:00-22:00, but were less active around midnight (Maffei et al., 2004) reflecting the activity of prey such as Mazama sp. and Tayassu sp. (Barrientos & Maffei, 2000). The overall resource-use and nocturnal/crepuscular activity pattern seen in EER generally agrees with previous studies, although the diurnal activity was higher than expected.
The spatial association between jaguar and prey species in EER can be explained by the relative abundance of prey, for example D. novemcinctus, Didelphis sp., M. temama, and N. narica in 2008, and for O. vetula and Sciurus sp. in 2012. However, in 2010 there was a positive association between jaguar and E. barbara, L. wiedii, M. temama, N. narica, and T. tajacu recorded in the camera traps, despite these species not being at high relative abundance. The contingency tables and GLMs suggested that A. cajanea, D. novemcinctus, Didelphis sp., L. pardalis, Meleagris ocellata, M. temama, N. narica, O. virginianus, and P. tajacu were associated with jaguar records in the traps, and these are reported as important prey by Ceballos et al. (2005), Chávez (2010), Chávez et al. (2007), and Davis et al. (2010).
The environmental variables that influence the presence of the jaguar vary along its range alongside its resource-use (Davis et al., 2010). In EER, the year of study had a significant effect on jaguar occurrence in models for all 4 years and the combined models. The photographic catch rates of all species decreased significantly in 2011, following a fire around the reserve, probably inhibiting the activity of all animals nearby. There was also a flood in 2010, which is likely to have had considerable impact on animal mobility, and therefore to have reduced their relative abundance as estimated by the camera traps. The main environmental factor in the models that determined jaguar occurrence was a positive association with acahual, probably because it encompasses many roads, which allow jaguars to move easily in search of prey, as seen in a study from Belize (Davis et al., 2010). Water availability had no impact on jaguar occurrence because EER has large and permanent water sources throughout the year, so it is never a limiting factor.
There was a positive effect of small and large birds and small mammals on jaguar occurrence, and although large birds are a commonly recorded prey item in felid diets, other small vertebrates are not. Models based on the presence of co-predators found that the crepuscular ocelot and nocturnal puma activity had a significant effect on jaguar activity. This agrees with previous studies from this area, and is thought to represent a temporary lack of segregation of predators (Chávez, 2010), also seen in Belize (Davis et al., 2010). However, according to the Akaike value, the principal variables that determined the presence of the jaguar in the study were the activity patterns of their main prey species, such as Odocoileus virginianus (white-tailed deer), Mazama temama (brocket deer), Dasypus novemncinctus (armadillo), and Crax rubra (curassow).
Many researchers have noted the effect of humans on the presence of jaguar (Ávila-Nájera et al., 2011; Ceballos et al., 2005; Zarza et al., 2007), particularly in relation to the decline of prey species, with fewer jaguar occurring in areas of intensive hunting (Chávez, 2010; Montiel et al., 1999). In EER, humans were the most frequently recorded species in the camera traps, and were one of the most important factors that influenced jaguar occurrence in the GLM models, agreeing with similar findings by Davis et al. (2010). Although human activity is an important factor that negatively affects the occurrence and activity of jaguars, it did not seem to influence their presence in this area. This may be because most photographic records were of workers, who pose a minimum threat to jaguar and their prey.
Even though EER is not a large area, it has suitable environmental components including adequate vegetation cover, water availability, and potential prey that allow for the continued presence of the jaguar. There is sufficient primary and secondary MSF within the EER and the surrounding area, as well as sufficient key prey, including ungulates (brocket deer and peccary), opossums, raccoons, and oscillated turkeys. However, these need to be maintained at adequate populations to support sustainable populations of jaguar. Jaguars frequently used roads within the reserve, which received low-level traffic, and were not affected by the presence of humans within the area. However, the long-term survival of jaguars in small areas such as El Edén relies upon the connectivity with other suitable protected areas through the establishment and maintenance of biological corridors. These results provide further evidence to support the planning and management of jaguars in the northern Yucatán Peninsula and adjacent areas.

