











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Fouquieria columnaris Kellogg (cirio) is a relictual species of the Sonoran Desert (Fig. 1) that is solely found in northwest Mexico. Three geographically separated populations are known, one is located in the Baja California Peninsula, another in Ángel de la Guarda Island in the Sea of Cortez, and a third one at Punta Cirio in the Sierra Bacha along the Sonoran coast. The latter population is found along a 45 × 4 km strip of land beginning 8 km south of Puerto Libertad and extending southwards along the coastline of the Sea of Cortez (Hall, 1973; Humphrey, 1974; Webb et al., 2014).

Figure. 1 Population of cirios at sampling sites. a. Development on the northern hillside. b. Cirio growing on rocky soil. c-e. Adult specimens of Fouquieria columnaris.
Cirio populations appear to be declining. The maximum longevity of this species is about 300-400 years (Webb et al., 2014). Paleoclimatic records document the presence of cirios at Sierra Bacha as early as nearly 9,000 years ago (Van Devender et al., 1995). As juvenile individuals and vegetation changes over time are infrequent, the reproduction rate of cirios appears to be limited (Turner et al., 2003). In addition, cirios exclusively reproduce by seeds, yet their genetic variability is apparently low (Hall, 1973). The sexual reproduction of cirios is primarily limited by the extreme conditions of the desert areas where they grow. Soil surface temperatures in the Sonoran Desert can reach nearly 70 ºC, resulting in high seedling mortality and preventing germination despite the presence of adequate moisture (Turner et al., 2003). Furthermore, the threatened or endangered status of cirios warrants the protection of this species; currently, this species is included in the Convention on International Trade in Endangered Species (CITES) Appendix II (http://cites.org/eng/app/appendices.php), which lists species that are not presently threatened with extinction but may become extinct in the future. Cirios grown in nurseries are commercially available in the USA, and their selling price can reach nearly $1000/ft (https://arboretum.arizona.edu/boojum).
Given this context, plant biotic interactions are important means of survival for different organisms in extreme environments such as the Sonoran Desert (Franklin et al., 2016). Bacteria, actinobacteria, fungi including arbuscular mycorrhizal fungi (AMF), oomycetes, viruses, and archaea live in the rhizosphere and are attracted by and feed upon rhizodeposits (i.e., nutrients, exudates, and mucilage) released by plant roots. In this context, plant-associated microbial communities can play a major role in the adaptation of plants to biotic and abiotic stress (Philippot et al., 2013). Also, bacteria and fungi generate numerous additional benefits for plants, such as providing iron to plants via siderophores and improving the soil structure. In particular, AMF in the rhizosphere of plants facilitate nutrient and phosphorus acquisition, increase drought and salinity tolerance, enhance disease resistance, increase water uptake, and make roots more geometrically and physiologically efficient. In addition, AMF play a significant role in soil N and C cycles (Andrew et al., 2012; Montaño et al., 2007; Willis et al., 2013). Although AMF are prominent and abundant in arid ecosystems (Bashan et al., 2007; Carrillo-García et al., 1999), plant-AMF interactions are poorly understood in the Sonoran Desert. For the most part, studies on the rhizosphere microbes of arid and semiarid plants have solely examined the abundance of microbe groups, resulting in an incomplete understanding of the diversity, ecology, and evolution of microbes (Franklin et al., 2016; Plascencia, 2013; Reyes-Quintanar et al., 2000; Soto-Aquino et al., 2012).
Given the above context, the questions motivating the present microbial survey of the rhizophere of cirio in Sonora were: a) as plant-microbe interactions play an important role in the survival of plants in extreme environments, what is the abundance of rhizospheric bacteria and fungi associated with cirio in Sonora? b) as the geographic distribution of relictual cirios is significantly restricted in Sonora and the reproduction of cirios is only sexual, is the genetic variability of cirios also limited? As a first approach, the abundance of culturable free-living nitrogen-fixing bacteria, phosphorus-solubilizing bacteria, actinobacteria, and filamentous and mycorrhizal fungi in the rhizosphere of cirio at Punta Cirio, Sonora, was analyzed. Then, an amplified fragment length polymorphism (AFLP) analysis was performed to measure the genetic variability of cirio.
Materials and methods
This study was conducted during the spring at Punta Cirio, Sonora, which is located in the Sonoran Desert along the coastline of the Sea of Cortez. Three hills were selected as study sites and labelled as site I (SI; 29°49’43” N, 112°37’01” W), site II (SII; 29°49’42” N, 112°36’54” W), and site III (SIII; 29°49’41” N, 112°36’44” W). Cirios are mostly located on northern slopes (Fig. 1a). The distance between SI and SII is 200 m and between SII and SIII is 240 m. The elevations (m asl) of the hills were categorized as low (L: 205-210), medium (M: 220-230), and high (H: 240-250). The distance between the low and medium elevations was 90-110 m and between the medium and high elevations was 90-110 m. The slope of SI, SII, and SIII were 19°, 18°, and 17° respectively. Because of the similarity of the cirios distribution and the environmental conditions at the 3 studied hills, cirios from the base to the top of each hill were considered. Therefore, cirios were sampled at similar elevations and distances from one another within a similar area per hill. Nine cirios were sampled per hill to allow for sufficient degrees of freedom in the statistical analyses. The elevation and coordinates were taken with a Magellan ProMark GPS, and the slopes were determined using a clinometer.
Triplicate soil cores were collected from the rhizosphere of 9 cirios at each sampling site, resulting in a total of 27 samples. At each site, a minimum of 25 g of soil and roots were taken 10-40 cm from the base of each cirio up to a depth of 30 cm after removing the topsoil. In the laboratory, each sample was flooded with 250 mL of water. These suspensions were stirred for 30 min before diluting to 10-7. The total bacterial counts were performed using total plate count medium, the free-living nitrogen-fixing bacterial counts were performed using Jensen medium, the phosphorus-solubilizing bacterial counts were performed using Picovskaya medium, and the actinobacterial counts were performed using Czapeck medium. Dilutions of 10-5 to 10-7 were inoculated in triplicate on plates. For filamentous fungi, dilutions of 10-3 to 10-5 were inoculated in triplicate on potato dextrose agar plates added with rose bengal and tartaric acid. Dilutions for the total quantification of bacteria and filamentous fungi were determined by preliminary tests and, for the rest of the considered microbes, were based on the literature. Samples were incubated at 25 °C for 3-7 days depending on the microbe, prior examination, and colony counting. Controls were carried out to ensure the accuracy of the microbial counts. The filamentous fungi were determined using the keys provided by Domsch et al. (1980).
The spores of AMF were isolated according to the wet sieving and decanting method of Gerdemann and Nicolson (1963). The colonization of AMF (%) was determined based on the observation of 60 root segments (each ca. 1 cm long) of 27 randomly sampled plants. Roots were washed and cleared with 10% KOH, stained with 1% lactophenol cotton blue, and mounted on slides for observation under the microscope (Giovanetti & Mosse, 1980; Phillips & Hayman, 1970).
All data were analyzed using SAS v6.08. The Kurtosis test was used to test data for normality. Then, the Levene’s test was applied to assess the equality of variance of the considered variables among the different sites. The total percentages of root colonization were angular transformed for subsequent analyses. Data were submitted to analyses of variance (Anova) considering a completely randomized design and a two-way factorial arrangement (sites and elevations) to detect interactions with respect to total bacteria, nitrogen-fixing bacteria, actinobacteria, filamentous and mycorrhizal fungi, and roots colonized by AMF. The multiple-range Tukey test was applied at a significance level of p < 0.05 when appropriate.
To determine the chemical and physical soil parameters, the 9 rhizospheric soil subsamples per hill were mixed, and 1 sample per hill was analyzed. The samples were mixed because of the empirical similarity of the soil. The percentage of saturation was determined using the gravimetric method, pH using a pH-meter at a water: soil ratio of 2:1, and electrical conductivity using an atomic absorption spectrophotometer at a water: soil ratio of 2:1 based on a reference solution of 0.005 N KCl. The mineral content (K, Ca, Mg, Na, Fe, Cu) was determined in addition to the sodium adsorption ratio (SAR) (Na/(Ca+Mg)/2)1/2, the exchangeable sodium percentage (1.475 SAR/1+0.0127 (SAR), and the soil texture according to the Bouyoucos hydrometer method. Finally, pedregosity was determined by sieve analysis, nitrates with chromotropic acid, phosphates by the Olsen method, and organic matter by the Walkley-Black method (Castellanos et al., 2000).
All perennial plants were counted in 27 quadrats (each 20 × 20 m) that were randomly selected at each elevation per site. The floristic composition and the main species of the arboreal and shrub strata were determined according to density and frequency. The filamentous fungi and plant species associated with cirios were compared among sites and elevations according to several ecological indexes: species richness, Shannon’s diversity (H’), Pielou’s evenness (J’), and Simpson’s dominance (D2). The correlation between filamentous fungal abundance and chemical and physical soil parameters were tested by Pearson correlation coefficients and principal component analysis.
To analyze plant genetic variability, samples were taken from 9 specimens corresponding to 3 individuals per site. One plant was randomly selected from each elevation level. The experimental unit consisted of a branch 20 to 30 cm long. To extract the DNA, freeze-dried tissues were powdered and dispersed in 15 mL of extraction buffer and incubated at 60 °C for 30-60 min. Then, 10 mL of a 24:1 chloroform: octanol solution (vol/vol) was added according to the technique of Saghai-Maroof et al. (1984). For the AFLP analysis, the methodology of Vos et al. (1995) was followed, which consisted of 3 steps: 1) restriction of DNA and ligation of oligonucleotide adapters, 2) selective amplification of sets of restriction fragments, and 3) gel analysis of the amplified fragments. Five combinations of randomly selected primers were used (E-ACA/M-CTA, E-ACA/M-CTG, E-ACA/M-CAC, E-ACA/M-CAG, E-ACA/M-CAA). The advantages of AFLP analysis include the ability to use a universal protocol in combination with different restriction endonucleases and the choice of adding 1 or more selective nucleotides in the PCR-primers to achieve optimal results relatively quickly without prior knowledge of DNA sequences from a large variety of organisms (Fry et al., 2009).
Based on the resulting banding patterns, a data matrix of discrete values (0 = absence and 1 = presence) was developed. Genetic similarity coefficients between 2 samples were estimated using a matrix and the Nei and Li (1979) index. In particular, genetic relationship matrices were created in the integrated similarity for qualitative data (SIMQUAL) program to produce dendrograms according to a sequential, agglomerative, hierarchical nested cluster analysis (SAHN) using the pair group method with arithmetic mean (UPGMA) in the NTSYS-pc program version 2.0.2i. If a single band is absent in at least 1 individual, the sampled population is considered polymorphic. Meanwhile, the similarity index compares the number of bands shared between a pair of individuals given the total bands per individual. The dendrogram stability was also evaluated by the bootstrap approach using mono- and polymorphic bands. Means similarities and standard errors were calculated based on 100 re-samplings (Felsenstein, 1985) in the statistical package PhylTools version 1.23. Bootstrapping is based on random sampling with replacement, which allows measures of accuracy to be assigned to each comparison.
Results
Total bacteria
Total bacterial counts did not significantly differ among the sites (p > 0.4) and elevations (p > 0.4); no interaction was found between the site and elevation factors (p > 0.18, Table 1). The minimum bacterial count was 5.9 log10 CFU g-1 of soil (SII-H), and the maximum count was 6.7 log10 CFU g-1 of soil (SI-H).
Table 1 Abundance of rhizospheric bacteria and fungi associated with Fouquieria columnaris.
| Units | SI | SII | SIII | p values | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H | M | L | H | M | L | H | M | L | Sites | Elevations | Interaction | ||
| Total bacteria | LogCFU | 6.7a | 6.1 a | 6.3 a | 5.9 a | 6.2 a | 6.7 a | 6.7 a | 6.4 a | 6.5 a | 0.450 | 0.410 | 0.186 |
| Nitrogen-fixing bacteria | LogCFU | 1.9a | 1.7a | 1.6a | 0.0a | 2.3a | 3.4a | 5.9b | 5.6b | 4.9b | 0.002 | 0.752 | 0.458 |
| Actinobacteria | LogCFU | 5.8 a | 5.1 a | 5.5 a | 4.9 a | 5.7 a | 5.8 a | 4.4 a | 6.1 a | 5.3 a | 0.925 | 0.594 | 0.624 |
| Filamentous fungi | LogCFU | 4.5 a | 4.8 a | 5.0 a | 5.0 a | 4.8 a | 5.0 a | 4.9 a | 4.6 a | 4.8 a | 0.777 | 0.738 | 0.831 |
| Mycorrhizal fungi | spores/100g | 129 a | 144ab | 234b | 109 a | 224 ab | 245 b | 182 a | 178 ab | 121 b | 0.163 | 0.006 | 0.006 |
| Colonized roots | percentage | 73 a | 74 a | 78 a | 73 a | 69 a | 73 a | 80 a | 73 a | 65 a | 0.525 | 0.502 | 0.138 |
Sites I, II and III; elevations: high (H), medium (M) and low (L). Averages with different letters are significant p < 0.05 by a Tukey test at the same line and correspond to 3 replicates of rhizospheric soil sample of cirio.
Nitrogen-fixing bacteria
The number of CFUs of free-living nitrogen-fixing bacteria ranged from undetected (SII-H) to 5.9 log10 CFU g-1 of soil (SIII-H). The counts did not differ per elevation (p > 0.7) but did change between sites (p < 0.002); no interactions between these factors were found (p > 0.4).
The nitrogen-fixing bacteria were significantly higher in SIII, which had an average of 5.9 ± 0.5 log10 CFU g-1 of soil (Table 1).
Phosphorus solubilizing bacteria
Phosphorus-solubilizing bacteria were not detected in any of the analyzed samples.
Actinobacteria
Total actinobacterial counts ranged from 4.4 (SIII- H) to 6.1 (SIII-M) log10 CFU g-1 of soil. No significant differences were found among sites (p > 0.9) nor elevations (p > 0.5), and no significant interaction between these factors were present (Table 1).
Filamentous fungi
Total filamentous fungal counts in the rhizosphere did not vary significantly among sites (p > 0.7) nor elevations (p > 0.7); no interactions between these factors were found (Table 1). The minimum fungal count was 4.5 log10 CFU g-1 of soil (SI-H), and the maximum count was 5.0 log10 CFU g-1 of soil (SI-L, SII-LH). The species richness of rhizospheric filamentous fungi ranged from 10 (SII-L) to 14 (SII-MH), Shannon’s diversity from 0.58 (SI-M) to 2.21 (SII-H). Pielou’s evenness from 0.24 (SI-M) to 0.86 (SIII-H), and Simpson’s dominance from 0.13 (SII-H) to 0.76 (SI-M) (Fig. 2). Species of Aspergillus and Penicillium predominated in 48% and 28%, respectively, of the total average CFUs (Table 2). Fusarium, Colletotrichum, and Alternaria, which can be phytopathogenic species, were also found. No fungi were observed to cause root rot in the 1,620 root segments analyzed during the AMF colonization test.
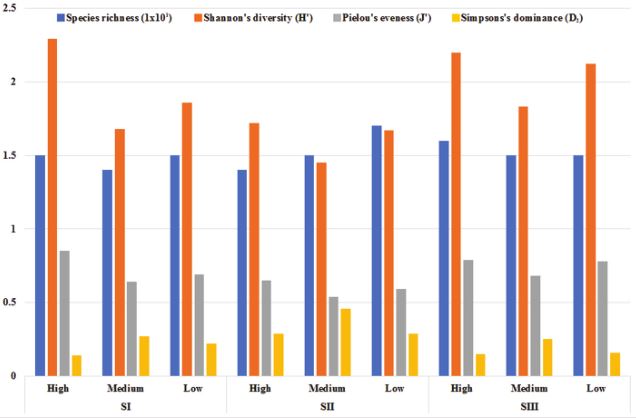
Table 2 Abundance of rhizospheric filamentous fungi associated with cirio.
| Fungi | Site I | Site II | Site III | ||||||
|---|---|---|---|---|---|---|---|---|---|
| H | M | L | H | M | L | H | M | L | |
| CFU x 10 3 /g of soil | |||||||||
| Alternaria sp. | 0.4 | ||||||||
| Aspergillus tamarii | 1.1 | ||||||||
| Aspergillus flavus | 2.2 | 2.6 | 41.3 | 20.0 | 9.1 | 20.5 | 30.0 | 6.8 | 154.6 |
| Aspergillus glaucus | 4.0 | 1.0 | 3.6 | 6.8 | 21.7 | 2.3 | 5.7 | 15.9 | 1.2 |
| Aspergillus niger | 6.5 | 18.5 | 42.7 | 23.0 | 9.3 | 55.4 | 15.8 | 3.1 | 41.9 |
| Aspergillus wentii | 0.8 | 0.3 | 3.3 | .1 | |||||
| Cladorrhinum sp. | 0.2 | ||||||||
| Cladosporium sp. | 8.9 | 1.1 | 1.1 | ||||||
| Colletotrichum sp. | 0.4 | 0.4 | 1.1 | 1.1 | 2.4 | ||||
| Fusarium spp. | 0.4 | 0.4 | 0.1 | 2.2 | 4.4 | 0.1 | 6.7 | ||
| Fusicoccum sp. | 0.8 | 1.4 | |||||||
| Geotrichum sp. | 1.3 | 1.1 | |||||||
| Gloeosporium sp. | 1.1 | 0.1 | 0.2 | ||||||
| Monilia sp. | 0.4 | 0.3 | |||||||
| Monocillium sp. | 3.3 | 11.3 | 11.1 | 2.2 | 13.3 | 1.3 | 6.7 | ||
| Sterile mycelium | 0.3 | 11.1 | 0.1 | 1.1 | 6.8 | ||||
| Mucor sp. | 0.67 | 0.1 | 1.3 | 0.2 | 16.2 | 1.3 | 0.2 | ||
| Penicillium spp. | 7.1 | 186.0 | 21.2 | 11.1 | 42.1 | 34.4 | 12.8 | 5.7 | 71.2 |
| Pestalotia sp. | 11.1 | 11.1 | |||||||
| Pleurostromella sp. | 0.2 | 0.6 | 13.8 | 13.3 | 1.1 | 12.4 | 3.3 | ||
| Stachybotrys sp. | 1.1 | 1.1 | |||||||
| Stemphylium sp. | 0.3 | 0.7 | 0.3 | 1.1 | 0.1 | 3.4 | |||
| Trichothecium sp. | 1.1 | 0.4 | 1.2 | ||||||
| Total | 33.6 | 215.0 | 133.7 | 114.5 | 104.9 | 118.8 | 100.4 | 48.9 | 297.4 |
Sites I, II and III; elevations: high (H), medium (M) and low (L). Averages of 3 replicates of rhizospheric soil sample of cirio.
Arbuscular mycorrhizal fungi
The number of AMF spores ranged from 109 (SII, H) to 245 (SII, L) per 100 g of soil, with a mean of 174 spores per 100 g of soil. No differences were observed among sites (p > 0.16). However, differences were observed according to elevation (p < 0.006), and interactions between factors were found (Table 1). The mean spore counts at high, medium, and low elevations were 140, 182, and 200, respectively, per 100 g of soil. Five morphospecies of Glomus spp. were found in the cirio rhizosphere. No significant interactions were found with respect to percentages of AMF colonization nor for the site and elevation factors (p > 0.05). Colonization percentages varied from 65% (SIII, L) to 80% (SIII, H), with a mean of 73%. Vesicles and arbuscles were consistenly observed.
Soil analysis
The pH was slightly alkaline, ranging from 8.0 to 8.25. The electrical conductivity varied from 0.5 (SII) to 0.94 dS m-1 (SI). The highest value for Ca + Mg (8 cmol kg-1) was observed at SI (Table 3). The Na content varied from 1 (SII) to 1.8 mg L-1 of soil (SIII). The SAR also ranged from 0.7 (SI, II) to 1.2 (SIII). The exchangeable sodium percentage varied from free (SI, II) to 0.6 (SIII). High sand content and pedregosity were observed and had corresponding values ranging from 79% to 89% and 47% to 55%, respectively. Nitrates and phosphates ranged from trace amounts up to 3.1 mg kg-1 and 23.2 mg kg-1, respectively. The organic matter content was low and represented 0.28% (SI) to 0.35% (SII, III) of soil.
Table 3 Physical and chemical analysis of rhizospheric soil of cirio.
| Soil parameters | Site I | Site II | Site III |
|---|---|---|---|
| Percentage of saturation | 30 | 30 | 27 |
| pH | 8.0 | 8.2 | 8.2 |
| EC (dS m-1) | 0.94 | 0.50 | 0.58 |
| Ca + Mg (mg L-1) | 8.0 | 4.0 | 4.0 |
| Na (mg L-1) | 1.4 | 1.0 | 1.8 |
| SAR | 0.70 | 0.71 | 1.27 |
| ESP | Free | Free | 0.6 |
| % Sand | 89.8 | 79.3 | 85.3 |
| % Silt | 8.2 | 14.5 | 11.5 |
| % Clay | 2.0 | 6.2 | 3.2 |
| Texture | Sandy | Loamy sand | Loamy sand |
| Percentage of pedregosity | 47.0 | 55.0 | 53.0 |
| NO- 3 (mg kg-1) | 3.13 | Traces | Traces |
| PO≡ 4 (mg kg-1) | 12.3 | Traces | 23.2 |
| % Organic matter | 0.28 | 0.35 | 0.35 |
EC: electrical conductivity; SAR: sodium adsorption ratio;
ESP: exchangeable sodium percentage.
The principal component analysis indicated that the data variability was explained by 2 factors, the first with 59.5% and the second with 40.4%. The abundance of filamentous fungi was correlated with the following soil parameters in factor 1: electrical conductivity (0.36), Ca + Mg (0.36), nitrate (0.36), texture (-0.36), and organic matter (-0.36), and factor 2: sodium (0.43) and phosphate (0.43) (Table 4).
Table 4 Correlation coefficient between the soil parameters and the main factors in the abundance of rizospheric filamentous fungi of cirio.
| Soil parameters | Factor 1* | Factor 2* |
|---|---|---|
| Percentage of saturation | -0.41 | |
| Electrical conductivity | 0.36 | |
| Calcium + magnesium | 0.36 | |
| Sodium | 0.43 | |
| Sodium adsorption ratio | 0.41 | |
| Exchangeable sodium percentage | 0.41 | |
| Sand | 0.33 | |
| Silt | -0.34 | |
| Clay | -0.30 | |
| Texture | -0.36 | |
| Nitrate | 0.36 | |
| Phosphate | 0.43 | |
| Organic matter | -0.36 |
*Factors 1 and 2 explained 59.5 and 40.4% of the data variability, respectively.
Vegetation analysis
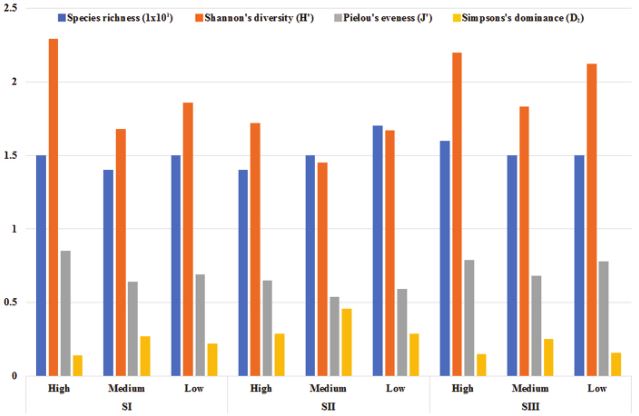
A total of 31 plant species were recorded. The species richness was similar among sites, ranging from 14 (SI-M, SII-H) to 17 (SII-L). The main species by frequency and abundance in the arboreal strata were Bursera microphylla A. Gray (Importance Value [IV]: 44%), Parkinsonia microphylla Torr. (IV: 8%), and Fouquieria columnaris (IV: 6%). The main species in the shrub strata were Ambrosia dumosa (A. Gray) W.W. Payne (IV: 52%), Encelia farinosa A. Gray & Torr. (IV: 32%), and Sphaeralcea sp. (IV: 20%). SI-H had the highest Shannon’s diversity (2.29) and Pielous’s evenness (0.85) and the lowest Simpson’s dominance (0.14), while SII-M had the lowest diversity (1.45) and evenness (0.54) and the highest dominance (0.46) (Fig. 3).
Genetic variability of cirio
The AFLP patterns generated via 5 primer combinations were similar among the 8 studied individuals. One sample was eliminated because of decomposition. Four hundred and forty-seven bands ranging between 100 and 500 bp, which were the product of 5 primer combinations, were analyzed. Of these, 264 were monomorphic, indicating a 40.9% polymorphism. The similarity coefficient varied between 0.86 and 0.92, with an average value of 0.89 (Fig. 4).
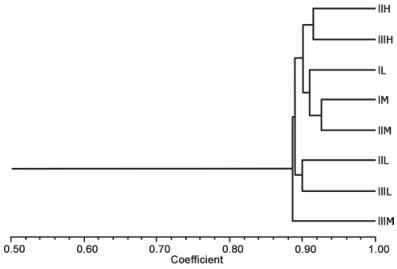
Figure. 4 Fouquieria columnaris dendrogram generated for samples collected at different sites (I, II, and III) and elevations (H = high, M = medium, L = low) with 5 primer combinations (E-ACA / M-CTA; E-ACA / M -CTG; E-ACA / M-CAC; E-ACA / MCAG; E-ACA / M-CAA). The scale corresponds to the coefficient of Nei and Li (1979).
Discussion
Environmental soil conditions, mainly moisture, aeration, temperature, organic matter, acidity, and availability of inorganic nutrients, affect microbiota density and composition. Microbial community size is directly related to the amount of organic matter in the soil (Vega et al., 2010). The pH was slightly alkaline at Punta Cirio, which is typical of arid zones and is usually due to the presence of free CaCO3. Humphrey (1974) identified a range of pH from 6.3 to 8.4 at 15 cirio sites, with values (8.0 to 8.4) similar to those in Punta Cirio at 6 localities. Esqueda et al. (2012) determined a range of pH from 6.3 to 7.9 at ten sites of the Plains of Sonora subdivision of the Sonoran Desert in central Sonora. High sand content and predregosity are common in desert areas where granite rock disintegrates to form a gravelly soil, which leads to the entrapment of moisture-retaining mulch (Webb et al., 2014). A more accurate physical and chemical soil analysis of each cirio would be interesting because the abundance of filamentous fungi was correlated with some soil parameters.
Andrew et al. (2012) observed that microbial communities in Sonoran Desert soils are primarily shaped by soil characteristics associated with geographic location, while rhizosphere associations are secondary factors. The operational taxonomic units suggested that the community of soil and rhizosphere microbes contain shared species and several common and abundant taxa, but the vast majority of taxa are rare and unique. Several soil properties, particularly pH and carbon content, were significantly correlated with community diversity.
Reyes-Quintanar et al. (2000) studied the total bacterial count of arid soils with and without erosion associated with Prosopis laevigata (Humb. & Bonpl. ex. Willd.) M.C. Johnst. and Neobuxbamia tetetzo (J.M. Coult.) Backeberg. In soils with and without erosion, the total mean bacterial counts varied from 5.6 to 7.3 log10 CFU g-1 of soil and from 5.8 to 7.8 log10 CFU g-1 of soil, respectively, wherein the lower means were related to low amounts of soil organic matter (1.1%). In the same locality, Soto-Aquino et al. (2012) measured bacteria and found 10.5 to 13.8 log10 CFU g-1 of soil; these authors attributed the significant variability in the total bacterial counts to the particular plant species and seasonal environmental conditions.
The low amount of nitrogen in the soil and the presence of nitrogen-fixing bacteria suggest that nitrogen-fixing bacteria could significantly contribute nitrogen to cirios. However, the abundance of nitrogen-fixing bacteria at Punta Cirio was not correlated with nitrate content. In a prior study, the nitrogen-fixing bacterium Azospirillum brasiliense was found to significantly promote growth in young cacti such as Pachycereus pringlei Britton & Rose, Stenocereus thurberi (Engelm.) Buxb., and Lophocereus schottii (Engelm.) Britton & Rose in desert areas with soil erosion (Bashan et al., 1999). The authors of this study also demonstrated the feasibility of bacterial inocula for increasing the establishment of cacti in disturbed areas and for potentially stabilizing soils. Therefore, the identification of nitrogen-fixing bacteria species associated with cirios is important, as these species could have potential applications.
Meanwhile, phosphorus-solubilizing bacteria were undetected in the cirio rhizosphere. The presence of phosphorus-solubilizing microfungi, which are abundant in arid soils, may compensate for this absence (Vera et al., 2002). Mycorrhizae also play a key role in the assimilation of soil phosphorus; plants with mycorrhizal associations can increase phosphorus absorption by 3 to 5 times (Montaño et al., 2007). In a later study, the absence of phosphorus-solubilizing bacteria in the cirio rhizosphere in Sonora may be confirmed using a lower dilution factor and a higher incubation temperature (30 °C) for the samples (Gupta et al., 2012).
With respect to actinobacteria, Plascencia (2013) found average counts of 4.4 to 4.9 log10 CFU g-1 in the rhizosphere of Bouteloua curtipendula Torr. in several arid areas of Mexico. These values are similar to those recorded in the cirio rhizosphere. Meanwhile, in the arid soils of P. laevigata and N. tetetzo, the average actinobacterial counts were 4.7 vs. 6.4 log10 CFU g-1 of soil in areas with vs. without erosion (Reyes-Quintanar et al., 2000). The CFUs of the eroded areas were similar to those at Punta Cirio.
In addition, in the rhizosphere of P. laevigata and N. tetetzo, filamentous fungal counts of 2.6 and 3.1 log10 CFU g-1 of soil, respectively, were found in arid soils with erosion vs. 3.7 and 3.9 log10 CFU g-1 of soil, respectively, in arid soils without erosion (Reyes-Quintanar et al., 2000). Meanwhile, in the rhizosphere of Viguiera dentata (Cav.) Spreng. and Ferocactus latispinus Britton & Rose, average filamentous fungal counts of 10.9 and 13.7 log10 CFU g-1, respectively, were determined. In the latter case, these relatively high counts in the dry season could be due to the higher content of organic matter of these soils (Soto-Aquino et al., 2012). Finally, Plascencia (2013) quantified 2 to 2.8 log10 CFU g-1 of filamentous fungi in the rhizosphere of B. curtipendula in several arid soils of Mexico.
The predominance of Aspergillus and Penicillium species has been recorded in other sites of the Sonoran Desert by Bácame-Valenzuela et al. (2015), Padilla et al. (2006), and Ranzoni (1968). This prevalence is common in arid zones because the species of these genera tolerate wide ranges of temperature, pH, and soil salinity (Domsch et al., 1980; Klich, 2002; Vera et al., 2002). Notably, the xerophilous species of the subgenus Aspergillus and many species of the Nidulantes and Circumdati section occur more frequently than expected in desert soils (Klich, 2002). Interestingly, some species of these genera can solubilize phosphorus making it more available to plants. This is highly relevant as phosphorous is the limiting factor for plant growth in desert regions. At Punta Cirio, the soil texture has a high sand content, which leads to low moisture retention (10 to 20%), loss of nutrients, and a high-water infiltration rate in addition to continuous erosion problems. The sodium concentration of soil was low at Punta Cirio. However, Humphrey (1974) found high levels of sodium (average of 607 mg kg-1) in 11 cirio localities in the Vizcaíno subdivision of the Sonoran Desert in central Baja California, whereas Esqueda et al. (2012) reported 70-198 mg kg-1 in 10 sites of the Plains of Sonora subdivision of the Sonoran Desert in central Sonora.
The number of AMF spores encountered in the soils of the present study is high compared to the cirio soils analyzed in Baja California where less than 5 spores per 100 g of soil were found (Rose, 1981). Spore abundance can also be influenced by seasonality (Schachtman et al., 1998) and soil fertility conditions (Bethlenfalvay et al., 1984). Among the most important limiting factors of vegetation in arid areas are the lack of water and nutrients, especially phosphorus. One study in the California Desert documented the colonization by AMF of 38 perennial species belonging to 19 families (Bethlenfalvay et al., 1984). The sandy soils at these prior desert localities were poor in phosphorus and nitrogen, and mycorrhizae distribution was not random but dependent upon host plants and climatic and soil conditions. Given these characeristics, the inoculum potential of AMF in desert soils is relatively low but remains possible in the upper layers of soil, although hyphal fragments should be used rather than spores (Bashan et al., 2000).
Rose (1981) reported that Glomus macrocarpus var. geosporus and G. microcarpus were associated with cirios in Baja California; these fungi developed vesicles and arbuscules in the roots of cirios. In another study conducted in Baja California, Bethlenfalvay et al. (1984) found that all plants sampled in 4 arid and semiarid sites (38 species belonging to families such as Agavaceae, Cactaceae, Leguminosae, etc.) were colonized by AMF (6 species). Among these, Fouquieria splendens Engelm. was in symbiosis with Glomus epigaeum and Glomus sp. Meanwhile, Bashan et al. (2007) found 23 mycorrhizal species in the rhizosphere of cirios in Baja California belonging to the genera Glomus (12 spp.), Appendicispora (3 spp.), Scutellospora (3 spp.), Archaeospora (1 sp.), Entrophospora (1 sp.), Intraspora (1 sp.), Pacispora (1 sp.), and Paraglomus (1 sp.); in this latter study, consistent colonization of cirio roots by hyphae, vesicles, and arbuscules was observed.
In another Sonoran Desert locality, Carrillo-García et al. (1999) found that all perennial plants belongning to 46 species and 23 families, including Burseraceae, Cactaceae, Euphorbiaceae, Fabaceae, and Zygophyllaceae, were mycorrhized at a colonization rate ranging from < 10% to > 70%. In disturbed areas, the roots of pioneer plants showed high mycorrhization. In contrast, columnar cacti, e.g., Pachycereus pringlei Britton & Rose, Stenocereus gummosus (Engelm.) A.C. Gibson & K.E. Horak, and S. thurberi (Engelm.) F. Buxb., were colonized at a rate of less than 10% and were typically found in association with nurse plants such as Prosopis articulata S. Watson and Olneya tesota A. Gray.
DNA polymorphism was low considering that cirio only spread by seed. Although the relationships between samples varied according to the utilized initiators during the DNA analysis, the similarity coefficient was high. In addition, the dendrogram reveals limited variability. The distribution analysis of samples for each pair of primers indicated no genetic correlation between individuals of the same hill or elevation. Considering the 5 pairs of primers, a greater correlation seems to be found with elevation than geographical location. The similarities between the lowest and highest coefficients and the average similarity of each evaluation in the bootstrap analysis suggest that the cirio population of Sonora has a high level of genetic homogeneity.
The limited genetic variability of F. columnaris was also observed by Hall (1973), who studied 21 localities in Baja California and Sonora by empirically separating them into 4 phenotypic groups that were genetically analyzed by isozymes. In this latter case, the amount of protein and tissue weight of seeds and seedlings of 24 h decreased with increasing latitude, and analyses of amylase and esterase isozymes indicated high polymorphism. Even though the variation between groups was high, some isozyme patterns were similar at the intragroup level, reflecting genetic homogeneity. Of the 4 phenotypic groups present in Baja California, 1 genetically matched the germplasm from Sonora. The cirios restricted distribution at Punta Cirio, e.g., to a 45-km strip, and limited genetic variability suggest the occurrence of inbreeding. Endemic plant species with extremely narrow geographical ranges (< 100 km2) often have a few small-sized populations and tend to be more vulnerable to extinction because of genetic drift and inbreeding effects (Solórzano et al., 2016).
Further integrative studies in soil microbiology are required, especially those that would holistically consider the various mechanisms at play in the rhizosphere. Such studies would undoubtedly improve the prediction and management of the microbiota in the rhizosphere. Clearly, going “back to the roots” of natural plant communities holds great promise for conserving biodiversity (Philippot et al., 2013). Do cirio-bacteria-fungi interactions, play an important role in the maintenance of species diversity? The pursuit of greater knowledge of species interactions in desert ecosystems is only beginning.

