











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Successive cambium is a cambial variant that involves the development of more than one cambium, resulting in 2 or more vascular rings of secondary xylem and phloem. Successive cambia have been reported in 34 vascular plant families, predominantly in the order Caryophyllales (Robert et al., 2011). In the Amaranthaceae, which belongs to this order, successive cambia have been described in both the stem and root, and in both organs, they form concentric vascular rings, either complete or incomplete (Carlquist, 2003, 2007; Joshi, 1937; Rajput, 2002).
Tamaio, Neves, Brandes, and Vieira (2011) showed that multiple cambia generate several vascular rings that differ between species with successive cambium from those with composite vascular rings. According to these authors, when composite vascular rings are present, all cambia remain active and have a determined number. This is why species with composite vascular cambia show little difference in the dimensions of the tracheary elements and fibers (axial cell elements) that constitute their secondary xylem. However, when successive cambia are present, the internal cambia (closer to the pith) cease their activity, while those closer to the stem periphery remain active. When not all of the successive cambia remain active, the dimensions of axial system cells vary significantly. For example, from pith to periphery, they tend to decrease in size in Stegnosperma Benth. (Phytolaccaceae; Horak, 1981), they can be of equal size to slightly longer in Cycas L. (Cycadaceae; Terrazas, 1991) and they tend to increase in size in Phytolacca dioica L. (Phytolaccaceae; Wheat, 1977), Abuta grandifolia (Mart.) Sandwith (Menispermaceae; Tamaio & Brandes, 2010) and Cocculus hirsutus (L.) Diels (Menispermaceae; Rajput & Rao, 2003). It has been hypothesized that in species with successive cambia, tracheary elements and fiber lengths are longer in the most internal ring, which comes from the fusiform initials of vascular cambium, while the tracheary elements and fibers that originate from parenchyma cells would be shorter. The alternative hypothesis is that the tracheary element and fiber lengths increase from pith to the stem periphery, as has been described for other plants with only one vascular ring (Panshin & de Zeeuw, 1980).
Iresine latifolia (M.Martens & Galeotti) Benth. & Hook.f. was selected as a scandent shrub of the Amaranthaceae and one of the species of Iresine P. Browne that is widely distributed in Mexico, ranging from tropical deciduous forest up to the cloud forest. It is assumed that this species, like other Amaranthaceae, has successive cambia. If all successive cambia are active in I. latifolia, it is expected that the lengths of tracheary elements and fibers will increase from the inner vascular ring toward the stem periphery. The aims of this study were to describe the wood of I. latifolia and to evaluate the lengths of tracheary elements and fibers in the different vascular rings.
Materials and methods
Three samples of I. latifolia were collected (Table 1); samples, each 10 cm long, were cut with a saw at 5 cm above the ground. The samples were fixed in formalin-acetic acid-ethanol (Ruzin, 1999) for 3 days and stored in glycerine-95% ethanol-water (1:1:1) solution until further processing. Two cylinders, each 2 cm long, were taken from each sample. The first of these stem cylinders was used to obtain transverse, radial and serial tangential sections (20-35 μm) using a sliding microtome. These sections were stained with safranin-fast green (Ruzin, 1999) and mounted in synthetic resin.
Table 1 Iresine latifolia samples collected.
| Individual | Height (m) |
Diameter (cm) |
Locality, state | Elevation (m a.s.l.) |
Vegetation | Collector |
|---|---|---|---|---|---|---|
| 1 | 2.0 | 4.6 | Parque ecoturístico Los Trocones, Cañón de la Libertad, Ciudad Victoria, Tamaulipas. |
338 | Scrub | Zumaya-Mendoza & Mora-Olivo 96 |
| 2 | 1.50 | 1.6 | Arroyo el León, Sierra Las Cacachilas, La Paz, Baja California Sur. |
496 | Tropical deciduous forest | Zumaya-Mendoza, León de la Luz & Domínguez 130 |
| 3 | 4.0 | 2.3 | Cañada El Muñiz, San Fernando, Chiapas. |
874 | Tropical subdeciduous forest | Samain et al., 2014-009 |
We added hydrochloric acid and phloroglucinol (Ruzin, 1999) to the transverse plane of the second stem cylinder to highlight the lignified tissue. Once the surface acquired a reddish color, we counted the number of secondary xylem rings and measured their width with the help of an American Optical 570 stereoscopic microscope. Next, secondary xylem was separated from each of the vascular rings using a disposable microtome knife. Each of the secondary xylem rings was macerated using Jeffrey's solution (Berlyn & Miksche, 1976) at 56 °C for 20-90 min to gather vessel element and fiber lengths; no tracheids were seen. Macerations were washed with tap water, and temporary slides were prepared for measurements of 50 fibers and 100 vessel elements for each vascular ring using Image-Pro Plus version 6.01 (Media Cybernetics) adapted for an Olympus BX-51 optical microscope. Variance analyses were performed using the general linear model (SAS, 2008) to evaluate whether there are significant differences between the rings within each sample.
Results
The wood of I. latifolia has a variable number of concentric vascular rings with conspicuous secondary xylem and phloem bands (Fig. 1A). The wood has growth rings abruptly delimited by vessel and fiber wall thickness. Porosity is diffuse. The vessels are in radial multiples of 2, with 16-22 vessels, sometimes 2-3 parallel radial multiples in each vascular fascicle, and occasional vessels are solitary (Fig. 1B); vessels are rounded in outline and occluded by gums, especially in the rings closer to the pith. Vessel diameter ranges from 23 to 78 μm with a mean of 44 μm and a wall thickness of 3 μm. The vessel elements are morphologically heterogeneous (Fig. 2), varying in length from 120 to 309 μm, and they have simple perforation plates from slightly inclined to almost horizontal end walls, with alternate intervascular pits (Fig. 1C) and pit diameter varies from 3.3 to 4.3 μm. The libriform fibers are nucleated and have simple pits, with no helical thickenings or septa (Fig. 1D, 2). Mean fiber length varies from 249 to 571 μm, diameter from 10.9 to 32.3 μm and wall thickness from 2.8 to 3.3 μm. The parenchyma is scanty and vasicentric. Additionally, it forms concentric unlignified bands (conjunctive tissue) of more than 3 cells wide and alternating with bands of secondary xylem. Rays are heterocellular, uni- and multiseriate, 4-10 cells wide, with short procumbent, erect and square cells. In the multiseriate rays, there is a center with meristematic activity, where xylem and phloem are differentiated (Fig. 1E).
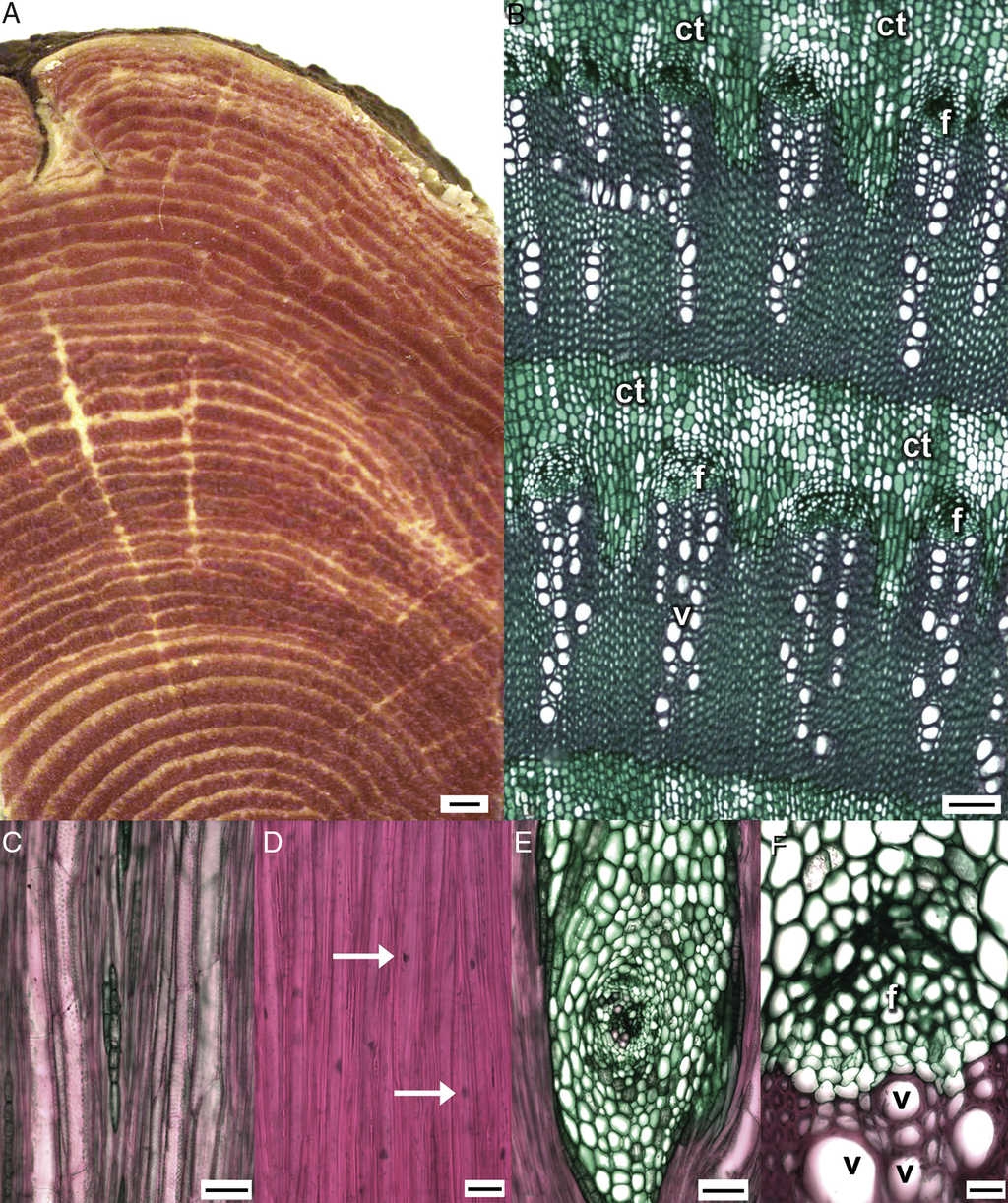
Figure 1 Wood of Iresine latifolia with successive cambium. Transverse (A, B, F) and tangential (C-E) sections. A, Stem showing successive cambia with abundant vascular rings; B, detail of vascular rings, showing the vessels in radial multiples, phloem, and conjunctive unlignified tissue; C, uniseriate ray and vessel elements with simple perforation plates and intervascular alternate bordered pits; D, nucleated libriform fibers; E, multiseriate unlignified ray with meristematic center; F, cambial zone with 2-3 layered cambial cells. Bars are 1mm in (A), 100μm in (B), 50μm in (C, E), and 20μm in (D, F). ct: conjunctive tissue; f: fiber, v: vessel; arrows: nuclei.

Figure 2 Diversity in morphology and sizes of vessel elements and fibers of Iresine latifolia wood. Bar is 100μm.
Additionally, the vascular cambium of each ring is distinctively active and differentiates new derivatives (Fig. 1F). The secondary phloem is composed of 2 regions: the first is a functional region of parenchyma cells and angular sieve tube elements with simple sieve plates and nucleated companion cells (Fig. 1F); the other is a non-functional region that forms a casket over the functional phloem, and only certain parenchyma cells expand between the obliterated sieve tube elements. Prismatic crystals were observed in ray cells, and sandy and prismatic crystals are present in the pith and the parenchyma cells (conjunctive tissue) between the vascular rings.
Vascular rings
The number of concentric secondary xylem rings is highly variable between samples, and the larger the stem diameter, the higher the number of secondary xylem rings (Fig. 3A), except for sample 3, which had fewer rings. In the 3 samples, the first 8 secondary xylem rings have a width of at least 1 mm. However, in sample 1, secondary xylem rings of 1 mm in width are present between the intermediate secondary xylem rings (Fig. 3A). The secondary xylem rings closer to the stem periphery were the narrower ones (<0.4 mm), and the separation between them is minimal, joining a vascular ring to another through parenchyma cells, which anastomose.
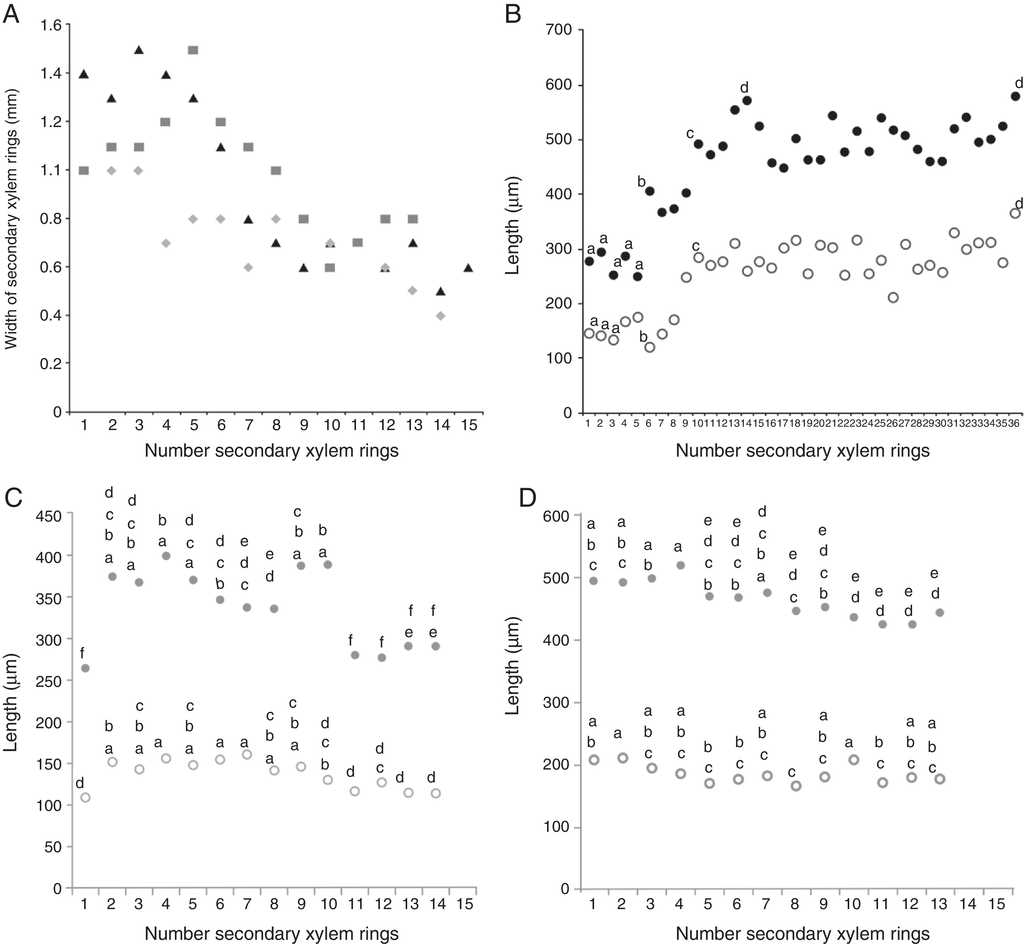
Figure 3 Widths of xylem rings and lengths of vessel elements and fibers in Iresine latifolia. A, Widths of xylem rings in sample 1 (triangles), sample 2 (diamonds), and sample 3 (squares); B-D, lengths of vessel elements (open circles) and fibers (close circles) from pith to periphery. B, Sample 1; C, sample 2; D, sample 3. Each point represents the mean of 50 measurements for fibers and 100 for vessel elements. Different letters indicate statistically significant differences.
Vessel element length
The length was almost constant in 2 samples, with no apparent differences between most of the secondary xylem rings (p > 0.05, Tukey), and differences between most secondary xylem rings were found only in sample 1, in which the length increased from pith to periphery (Fig. 3B-D).
Fiber length
The fibers are almost twice the length of vessel elements, due to intrusive growth. In samples 1 and 2, the length increases from pith toward periphery, and some significant differences were found between some rings (p < 0.05, Tukey), but in sample 3, the fibers decrease in length in the vascular ring closer to the periphery, and there are almost no differences between secondary xylem rings (Fig. 3D).
Discussion
Variation in the width of the vascular ring and the length of its axial cells
Variance analyses confirmed significant differences in the length of vessel elements and fibers only for some of the secondary xylem rings from pith to periphery for each of the I. latifolia samples studied. For vessel elements, there are no major differences between the first ring, which was formed from vascular cambium, and those that are closer to the periphery, which come from parenchyma cells that would correspond to rings 13 or 14. This lack of size difference between the first and last vascular rings formed in 2 of the samples (Fig. 3) suggests that the parenchyma cells from which the new cambium is differentiated reach the initial fusiform size early in the differentiation process due to their short length (<309 μm). In sample 1, there was always an increase in vessel element length from pith to periphery, confirming that when the environment is favorable to the plant growth, more wood is accumulated and larger cell sizes are therefore achieved, as is the case in other plant groups (Panshin & de Zeeuw, 1980). Moreover, the results in vessel element lengths support the assertion of Robert et al. (2011), who noted that the successive cambia are interlinked and remain active, which is why they reach the initial fusiform cell sizes.
In fibers, there were significant differences between those differentiated from vascular cambium and those derived from successive cambia, with no clear pattern. For example, in sample 1, their length increased, but in samples 2 and 3, fiber lengths decreased. In the 3 samples, the fibers became twice the length of vessel elements as a result of intrusive growth, similar to reports for C. hirsutus (Rajput & Rao, 2003), which also has successive cambia.
The variation in vessel element and fiber lengths in the 3 samples of I. latifolia is more diverse than reported for other species with successive cambia. Only in one of the samples studied was there an increase in the length of both types of cells from pith to periphery, as has been observed in several species of Cycadaceae, Menispermaceae or Phytolaccaceae (Rajput & Rao, 2003; Tamaio & Brandes, 2010; Terrazas, 1991; Wheat, 1977), or else it is maintained or decreased in the other 2 samples of I. latifolia, as in Stegnosperma (Phytolaccaceae; Horak, 1981). On the other hand, no association between the width of the vascular ring and the size of the vessel elements and fibers was found in any of the 3 samples. For further comparative studies of the wood in other Amaranthaceae species, we recommend measuring the length of the vessel elements and fibers in any of the 2 most external vascular rings, especially when the complete stem cylinder is not available.
Concentric successive cambia may develop complete vascular rings, discontinuous rings or an anastomosing network. The presence of anastomosis between the most external vascular rings has been observed in other Amaranthaceae species and appears to be a common trait (Rajput & Marcati, 2013) in species with successive cambia (Robert et al., 2011).
The wood
It is confirmed that I. latifolia has concentric successive cambia (secondary xylem bands alternating with secondary phloem bands) as observed in other Amaranthaceae species, as well as diffuse porosity, vessels in radial multiples, vessel elements with simple perforation plates, alternate intervascular pits, scanty-vasicentric parenchyma and nucleated libriform fibers (Carlquist, 2003; Rajput & Marcati, 2013; Rajput & Rao, 2000). This Iresine species has uniseriate rays with erect, square and short procumbent cells but also multiseriate rays with a meristematic center similar to the multiseriate rays reported for Hebanthe eriantha (Poir.) Pedersen (Amaranthaceae; Rajput & Marcati, 2013) and Suaeda monoica Forssk. ex J.F.Gmel (Chenopodiaceae; Lev-Yadun & Aloni, 1991). Other Iresine species should be examined to ascertain whether multiseriate rays with a meristematic center are characteristic of the genus or unique to I. latifolia, or whether they are associated with the shrub or scandent habit.
We conclude that I. latifolia wood is similar to that of other Amaranthaceae, with stem growth occurring due to successive cambia. Vessel element and fiber lengths increase from pith to periphery in I. latifolia with successive cambia as in other species with a single vascular ring, when growth conditions are favorable. Thus, cell size in species with numerous successive cambia reaches the optimal size of the fusiform initial to perform the proper water movement (vessel elements) and contribute to efficient support (fibers).

