











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La población de tapir centroamericano (Tapirus bairdii ) se ha reducido en un 50% en toda su área de distribución en los últimos 30 años (Castellanos et al., 2008). En México, su distribución actual abarca solamente alrededor del 45% de lo que era su distribución histórica (Nolasco, Lira y Ceballos, 2007). Esta reducción en las poblaciones se debe principalmente a la deforestación y fragmentación de su hábitat, a la cacería y a las muertes que se producen en encuentros incidentales con humanos (Naranjo, 2009; Robinson y Redford, 1994).
El tapir centroamericano es el mamífero terrestre más grande del Neotrópico (Tobler, 2002) y tiene un importante papel ecológico en la dinámica de las selvas por su actividad en la herbivoría, dispersión y depredación de semillas de numerosas especies vegetales (Naranjo, 1995; O'Farrill, Calmé y González,2007; Olmos, 1997). Habita preferentemente en áreas boscosas extensas (> 1,000 ha) con escasa perturbación y con cuerpos de agua permanentes (March y Naranjo, 2005). Sin embargo, los tapires también han sido registrados en selvas bajas caducifolias, manglares, acahuales y vegetación secundaria causada por disturbios naturales, e incluso en pastizales y cultivos (Foerster y Vaughan, 2002; Muench, 2001; Naranjo, 2009; Nolasco, 2009; Reyna-Hurtado, 2002).
El tapir centroamericano es considerado a nivel global en peligro de extinción por la Unión Internacional para la Conservación de la Naturaleza (UICN, 2012). En México, la Secretaria de Medio Ambiente y Recursos Naturales lo considera también en peligro de extinción, así como especie prioritaria de conservación (Semarnat, 2002). La conservación y manejo adecuado de este mamífero y su hábitat no será posible si se carece de información sobre sus abundancias, su dinámica y estructura poblacional, así como la disponibilidad y calidad de su hábitat. Factores fundamentales para desarrollar estrategias de manejo y conservación para especies en peligro de extinción (Foerster y Vaughan, 2002; Goulart et al., 2009; Naranjo, 2009).
El sureste de México constituye una región crítica para la conservación del tapir centroamericano en virtud de la extensión de hábitats adecuados como selvas y humedales que aún mantiene y el tamaño de las poblaciones que concentra: según estimaciones, cerca de 50% del total de la población global (Mendoza y Carbajal, 2011; Naranjo, 2009). Si bien existe consenso en cuanto al papel crítico que desempeñan las reservas del sureste del país (Montes Azules, La Sepultura, El Triunfo, Sian Ka'an y Calakmul), la mayoría de los estudios han sido llevados a cabo en las áreas naturales protegidas del estado de Chiapas. Existe muy poca información acerca de la importancia que tienen reservas de menor tamaño como la Reserva Estatal de Balam Kú para mantener las poblaciones de esta especie (Naranjo, 2009).
El objetivo de la presente investigación fue obtener una estimación de la abundancia relativa y la selección de hábitat del tapir centroamericano en 3 sitios dentro del macizo forestal de la Reserva de la Biosfera de Calakmul (RBC) y la Reserva Estatal Balam Kú (REBK). Sitios que se encuentran bajo el efecto del gradiente de humedad presente en la península de Yucatán, donde la precipitación se incrementa del norte al sur (Folan, Gunn, Eaton y Patch, 1983; Gunn y Adams, 1981). Estudios previos mencionan que el tapir centroamericano requiere hábitats con abundantes fuentes de agua y alimento (Foerster y Vaughan, 2002; Fragoso, 1997; Naranjo y Bodmer, 2002). Por lo que en este estudio intentamos responder si existe un efecto del gradiente de humedad en la abundancia relativa y la selección de hábitat de los tapires en esta región.
Materiales y métodos
Área de estudio
La RBC y la REBK se localizan al sureste del estado de Campeche (Fig. 1). Estas reservas se conectan con la Reserva de la Biosfera Maya en el Petén guatemalteco y con el Área de Conservación de Milpas-Río Bravo en Belice, englobando 3,073,998 ha de cobertura boscosa que forman a la segunda área de bosque tropical más grande del continente americano: la Selva Maya (Reyna-Hurtado et al., 2010). El clima predominante en esta zona es cálido subhúmedo (Aw), con una temperatura media anual de 25 °C (Arriaga et al., 2000; García-Gil, 2003). La característica climática más notable de la península de Yucatán es un incremento del gradiente de humedad del noroeste hacia el sureste (Folan et al., 1983; Gunn y Adams, 1981). La precipitación anual varía de 500 a 2,500 mm (García-Gil, 2003). La vegetación se compone principalmente de selvas altas y medianas subperennifolias, selvas medianas subcaducifolias, selvas bajas subcaducifolias, caducifolias y selvas bajas inundables, siendo la selva mediana subperennifolia la vegetación dominante (Carabias, Provencio, De la Masa y Rodríguez, 1999; Martínez y Galindo-Leal, 2002). El agua en Calakmul es escasa y la mayoría del agua de lluvia se infiltra hacia el subsuelo, excepto en las selvas inundables o "bajos" donde se almacena la lluvia temporalmente, poca escurre superficialmente en corrientes temporales o semipermanentes, almacenándose en cuerpos de agua conocidos localmente como "aguadas". Estos reservorios son la única fuente de agua para la fauna silvestre y para varias comunidades humanas durante la época seca (Reyna-Hurtado et al., 2010).
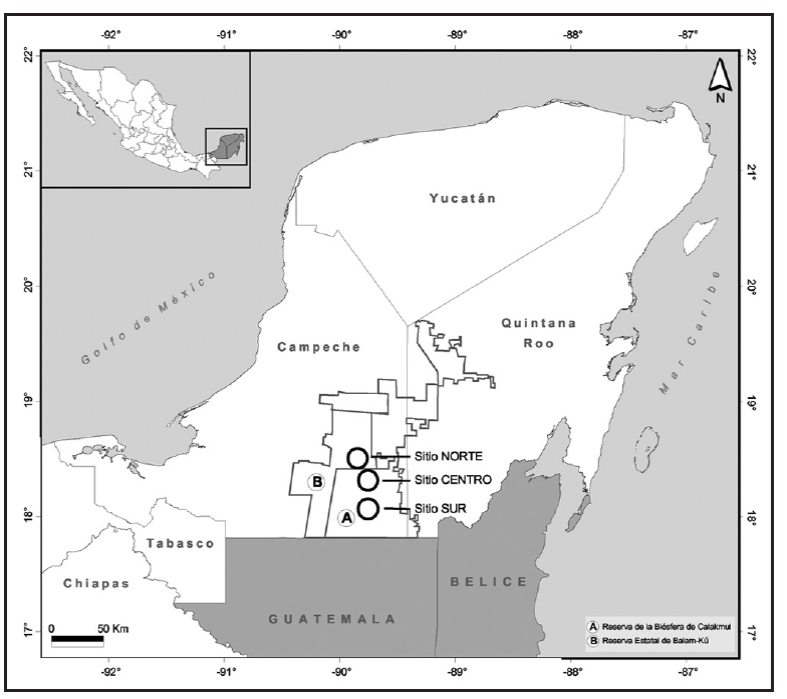
Figura 1 Sitios de estudio en la Reserva de la Biosfera de Calakmul y la Re serva Estatal de Balam Kú, Campeche, México.
El trabajo de campo se llevó a cabo durante los meses de abril a julio del 2012. Se trabajó en 3 sitios, 2 ubicados en la RBC (sitio sur y sitio centro) y 1 en la REBK (sitio norte) para abarcar así el gradiente de humedad que se presenta en la región (Fig. 1).
Abundancia relativa de huellas
En cada sitio se establecieron 5 transectos lineales de amplitud variada de 2 km cada uno, 30 km en total, con una distancia de al menos 3 km entre transecto. Para obtener el índice de abundancia relativa de huellas de los tapires (IAR), se recorrieron los transectos una vez al mes, realizando una búsqueda exhaustiva de huellas que se encontraran hasta 1 m a cada lado de la línea central del transecto. Debido al suelo cárstico de Calakmul y a que la mayoría del agua de lluvia se infiltra al subsuelo (García-Gil, 2003) la impresión de huellas no es muy clara, por lo que no fue posible tomar medidas para comparar entre los diferentes rastros encontrados en un transecto. Por ello, para evitar una sobreestimación de la abundancia, las huellas que se encontraban en conjunto o a una distancia menor a 50 m fueron tomadas como un solo individuo (Naranjo, 1995; Pérez-Cortés y Matus-Pérez, 2010). Los rastros fueron borrados para evitar ser contados de nuevo en el siguiente recorrido.
El índice para estimar la abundancia relativa consiste en dividir el número de rastros encontrados por kilómetro recorrido (Caughley, 1977):

donde NH es el número de huellas observadas y km son los kilómetros recorridos. Se obtuvo un IAR total para toda la zona de estudio y para cada uno de los sitios en estudio.
Abundancia relativa de foto-capturas
En cada sitio se ubicaron 3 aguadas elegidas por su accesibilidad en época de lluvias, 9 aguadas en total, en las que se instalaron 3 modelos de cámaras-trampa (PC800 Hyperfire professional Reconix, Inc. Moultrie Game Spy l-65 y Cuddeback Capture IR digital scouting camera). Se colocaron 4 cámaras por sitio y se mantuvieron funcionando continuamente. El esfuerzo de muestreo se obtuvo al multiplicar el número de cámaras utilizadas, por el número de días que duró el muestreo (días-trampa, periodos de 24 h en que las cámaras estuvieron activas; Dillon y Kelly, 2007)
Se contabilizó el número de fotografías de tapir en cada aguada cada 24 h (capturas por día), todos los registros de tapir en un mismo día se tomaron como un solo individuo; únicamente se cuantificaron como más de un registro cuando se pudieron diferenciar individuos por alguna seña en particular; por ejemplo, sexo, edad, cicatrices. Se obtuvo el índice de abundancia relativa de foto-capturas que se expresó como el número de capturas fotográficas por 1,000 días-trampa (Maffei, Cuéllar y Noss, 2002; Seydack, 1984):
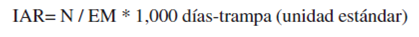
donde N es el número de visitas independientes registradas por aguada y EM es el esfuerzo de muestreo. El IAR se obtuvo para toda la zona de estudio y para cada uno de los sitios.
Se comprobó si los datos se ajustaban a la distribución normal a través de una prueba de bondad de ajuste de Kolmogorov-Smirnov. Para saber si había diferencias significativas en el número de registros de tapir entre los sitios (huellas y foto- capturas) y entre transectos y aguadas dentro de cada sitio, se aplicaron pruebas de Kruskal-Wallis y de Mann-Whitney (Sokal y Rohlf, 1981). Todos los análisis estadísticos se realizaron con el programa SPSS para Windows Versión 8.0.
Disponibilidad de alimento
En cada uno de los transectos se realizó el conteo e identificación de los frutos que son conocidos por formar parte de la dieta del tapir (Cruz, 2001; Naranjo, 1995; O'Farrill, Calmé y González, 2006; Pérez-Cortés y Matus-Pérez, 2010). Para ello, se identificaban los frutos que se encontraban en el transecto. Una vez que se verificaba la presencia del árbol de donde presuntamente provenían, se contabilizaban los frutos encontrados en el suelo en 2 m2 alrededor de dicho árbol.Con estos datos se obtuvo el porcentaje de frutos para cada uno de los sitios.
Debido a que en esta región no se conocen todas las especies vegetales que forman parte de la dieta del tapir, se calculó únicamente el porcentaje de la cobertura vegetal del sotobosque en cada sitio. Para ello, se tomaron fotografías de la vegetación a un lado del transecto colocando la cámara a 1 m de altura con el zoom desactivado (objetivo: 4 mm). La toma de fotografías se hizo una vez al mes, al inicio, a la mitad (1 km) y al final del transecto (2 km). Posteriormente, estas fotografías se colocaron en una plantilla con una cuadrícula de 10×10 y se contaron los cuadros ocupados por plantas. Con este método se obtuvo el porcentaje de la cobertura vegetal del sotobosque para cada sitio. Para determinar si había relación entre el porcentaje de frutos y el porcentaje de cobertura vegetal del sotobosque con el IAR de huellas, se realizó una correlación de Spearman con la ayuda de la plataforma estadística R versión 2.11(c).
Hábitat disponible
Para conocer la proporción del hábitat disponible en los transectos marcados, se registró el tipo de vegetación de acuerdo con la reclasificación del Instituto Nacional de Ecología (INE, 1999) cada 100 m de kilómetro recorrido, además, cada vez que se encontraba un registro confiable de tapir se registraba también el tipo de vegetación. Se contrastó la disponibilidad de hábitat que se obtuvo en los transectos cada 100 m, independientemente de la presencia o ausencia del tapir, con la proporción de cada tipo de vegetación donde hubo registros de tapir a través de un análisis de χ2. Utilizando el número de huellas encontradas, se obtuvo la frecuencia observada, la frecuencia esperada y los intervalos de Bonferroni para cada tipo de cobertura vegetal utilizada por la especie. Estos análisis se realizaron empleando el programa HABUSE 4.0 (Byers, Steinhorst y Krausman, 1984).
Resultados
Abundancia relativa de huellas
En total fueron registradas 40 huellas de tapir entre los 3 sitios. El IAR total fue de 0.33 huellas/km para toda el área de estudio. En el sitio norte el IAR fue de 0.42 huellas/km, en el centro de 0.25 huellas/km y en el sur de 0.32 huellas/km. No se encontraron diferencias significativas al contrastar la abundancia de registros de tapir entre los 3 sitios de estudio (H = 0.783; p = 0.67). El mes de julio fue el mes en que se registró un mayor número de huellas en los 3 sitios con 0.50 huellas/km.
Abundancia relativa de foto-capturas
El esfuerzo de muestreo fue de 1 464 días-trampa para todo el estudio. El índice de abundancia total fue de 17 registros/1,000 días-trampa, el sitio con mayor registro de fotografías de tapir fue el sur con 34 registros/1,000 días-trampa, seguido por el norte con 10 registros/1,000 días-trampa y el centro con 6 registros/1,000 días-trampa. El tapir estuvo presente en 8 de las 9 aguadas en estudio. La aguada que mayor cantidad de registros tuvo fue la aguada Bonfil que se ubica en el sitio sur, con una abundancia de 53 registros/1,000 días-trampa, asimismo, Bonfil fue la única aguada en que se registró presencia de tapir durante los 4 meses de estudio. No se encontraron diferencias significativas en la abundancia de registros fotográficos entre los 3 sitios (H = 3.3; p = 0.187). El mes de julio fue el mes en el que la mayoría de las aguadas se mantuvieron llenas.
Disponibilidad de alimento, frutos
En total fueron registradas 21 especies de frutos en el área de estudio, de las cuales 8 han sido registradas como parte de la dieta del tapir (Cruz, 2001; Foerster y Vaughan, 2002; O'Farrill et al., 2006; Pérez-Cortés y Matus-Pérez, 2010). De estas, Manilkara zapota representó el 40% del total de especies encontradas y su fruto fue el único queestuvo presente en todos los sitios durante los meses que duro el muestreo. El sitio con mayor abundancia de frutos fue el sur con 55.7%, seguido por el centro con 26.5% y el norte con 17.7%. La correlación de Spearman no mostró una relación significativa entre el porcentaje de frutos y el IAR de huellas (r s = .049; p = 0.7).
Cobertura vegetal del sotobosque
En los 4 meses de estudio, la cobertura vegetal del sotobosque fue muy parecida en los sitios centro y sur, con 41.9 y 41.3%, respectivamente, mientras que en el sitio norte se registró una menor cantidad de plantas en el sotobosque con 16.6%. La correlación de Spearman no muestra una relación significativa entre el porcentaje de cobertura vegetal del sotobosque y el índice de abundancia relativa de huellas (r s = -.096; p = 0.4).
Hábitat disponible
En los 3 sitios de estudio la selva mediana fue el tipo de vegetación dominante, representando 89% en el sur, 39% en el centro y 51% en el norte. En los 3 sitios se encontraron selvas bajas subcaducifolias, siendo más frecuente en el sitio norte, el cual fue el único sitio en el que se encontraron selvas caducifolias, así como en el sitio sur no fueron registradas selvas bajas inundables. La selva alta fue el tipo de vegetación con menos representación en el sitio de estudio.
Preferencias de hábitat
La vegetación predominante en cada punto donde se encontró una huella de tapir fue la selva mediana, con un total de 28 huellas, 6 huellas se registraron en la selva baja inundable, 5 en la selva baja subcaducifolia, 1 en la selva baja caducifolia y ninguna en la selva alta. Aunque se registró un mayor número de huellas en la selva mediana, el análisis de χ2 determinó que no hubo diferencias significativas entre el uso que el tapir hace de cada tipo de vegetación y que usa los diferentes tipos de vegetación de acuerdo con su disponibilidad (N = 40; χ2= 3.52; p = 0.47).
Discusión
Los resultados no muestran diferencias significativas en los IAR de huellas entre los 3 sitos de estudio, sugiriendo que los 3 sitios cuentan con las características necesarias para la presencia de tapir y que en las reservas de Calakmul y Balam Kú el tapir puede habitar en zonas más secas, siempre y cuando los recursos necesarios para su supervivencia estén disponibles.
La abundancia relativa de tapir registrada en este estudio (0.33 huellas/km) resultó menor a la documentada por Pérez-Cortés (2011) (1.48 huellas/km). La diferencia entre abundancias se debe a las características espaciales de cada estudio, ya que uno de los objetivos de Pérez-Cortés (2011) era estimar el uso que dan los tapires a las aguadas en diferentes épocas del año, por lo que los transectos para la búsqueda de huellas partían directamente de las aguadas, aumentando así la posibilidad de encontrar un mayor número de rastros de tapir.
En el mes de julio se registraron más huellas. Esto puede deberse a que en julio existe una mayor cantidad de agua en aguadas, lo que podría ocasionar un aumento en el movimiento de los tapires, sin tener que localizar sus movimientos cerca de los cuerpos de agua remanentes o de parches de vegetación con alimento. Igualmente Pérez-Cortés (2011) registra que durante la estación seca los tapires son más abundantes alrededor de los cuerpos de agua perennes, mientras que en la estación de lluvias aumenta su abundancia en las aguadas pequeñas e intermitentes.
En la península de Yucatán sólo 2 estudios más han documentado abundancias fotográficas de tapir, uno de ellos en la RBC donde registran una abundancia relativamente mayor a la que se registró en este estudio (Pérez-Cortés, Enríquez, Sima-Panti, Reyna-Hurtado y Naranjo, 2012). Otra investigación se llevó a cabo en el Área de Protección de Flora y Fauna Balam Ka'ax en Quintana Roo (Pérez-Cortés y Matus-Pérez, 2010). Estos autores documentan una abundancia relativa de tapires alta, lo que puede indicar que a pesar de ser una reserva de menor tamaño al igual que Balam Kú, la ANP Balam Ka'ax tiene características importantes para la subsistencia de esta especie. Igualmente en Oaxaca, México, se realizó un estudio con cámaras-trampa para conocer las abundancias de mamíferos medianos y grandes en la región de los Chimalapas (Lira-Torres y Briones-Salas, 2012). Los resultados indican que a pesar de que el tapir es una de las principales presas de caza por las comunidades rurales, es una de las especies de mamíferos más abundantes en la región. Estos autores apuntan a que las características accidentadas e inaccesibles de la zona han ayudado a que esta especie aún sea abundante en esta región. Las abundancias relativas más altas obtenidas con cámaras-trampa, hasta el momento se registraron en la región de Talamanca en Costa Rica, en una investigación para conocer la abundancia de estos mamíferos a diferentes altitudes (González-Maya, Schipper y Rojas-Jiménez, 2009). Estos autores encontraron las mayores abundancias en sitios con altitudes mayores a 2 500 m snm y con mejor estado de conservación, y lo atribuyen a que estas zonas se encuentran más aisladas de poblaciones humanas (Tabla 1).
Tabla 1 Índices de abundancia relativa de tapir centroamericano (Tapirus bairdii) obtenidos con cámaras-trampa en México y Centroamérica.
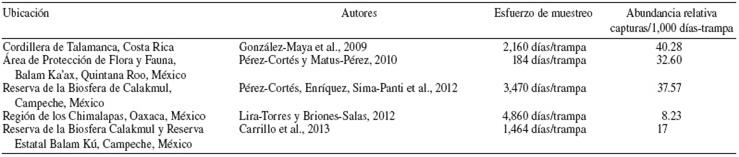
Los resultados del presente estudio indican que el IAR de foto-capturas no fue estadísticamente diferente entre sitios, sin embargo, en el sur, la aguada Bonfil obtuvo la mayor abundancia de fotografías comparada con las demás aguadas. Bonfil se caracteriza por ser una aguada perenne desde el año 2008 y con una gran cantidad de vegetación hidrófila de la que se puede llegar a alimentar el tapir. Igualmente, Pérez-Cortés et al. (2012) documentan mayor cantidad de registros fotográficos para esta aguada, la cual se encuentra a 20 m de la carretera que lleva hacia las ruinas mayas de la RBC y es visitada constantemente por turistas. A pesar de que es una aguada con cierto grado de perturbación, los tapires siguen accediendo a ella, quizá por ser un sitio seguro donde encontrar una fuente de agua permanente y a que los hábitos nocturnos del tapir impiden el encuentro con turistas. Sin embargo, no sabemos en un futuro qué tanto pueda afectar la presencia humana constante en las visitas de los tapires a esta aguada.
Existen diferencias en la cantidad de recursos alimenticios entre ambas reservas. La RBC fue el sitio con mayor cantidad de frutos encontrados en el suelo y mayor cobertura vegetal del sotobosque, en especial la parte sur de la zona de estudio. Mientras que la REBK tuvo un menor porcentaje de frutos, menor cobertura vegetal del sotobosque, mayor proporción de selvas bajas subcaducifolias y de selvas caducifolias. Estas diferencias pueden deberse a la influencia del gradiente de humedad norte-sur presente en la península delimitando a la REBK como una zona más seca que la RBC.
El fruto de M. zapota fue el único fruto que estuvo presente en los 3 sitios durante los 4 meses de estudio. Al ser un recurso presente en toda el área de estudio, el zapote podría tener un papel importante en la sobrevivencia del tapir, sobre todo, en las zonas secas donde los demás recursos pueden ser escasos. Reyna-Hurtado, Rojas-Flores y Tanner (2009) documentan resultados similares para los pecaríes labios blancos (Tayassu pecari ) en la RBC, donde observaron que los pecaríes son capaces de sobrevivir en el bosque semiseco, tomando ventaja, entre otros recursos, de los frutos como el zapote y de algunas herbáceas.
En los sitios bajo estudio, el tapir no mostró preferencias por algún tipo de hábitat en particular, ya que está usando todos los tipos de vegetación de acuerdo con su disponibilidad. Estudios previos en la RBC y en la selva Lacandona en Chiapas documentan resultados similares (Naranjo, 2001; Reyna-Hurtado y Tanner, 2005; Sánchez-Núñez, Ortiz y Arellano, 2011), lo que sugiere que los tapires pueden llegar a ser generalistas en el uso de hábitat. Sin embargo, el pequeño tamaño de muestra obtenido en este estudio no permite afirmar este resultado, ya que además, existen otros estudios que sí muestran una preferencia de hábitat por parte de los tapires (Naranjo y Cruz, 1998; Tejeda-Cruz, Naranjo, Cuarón, Perales y Cruz-Burguete, 2009). Aunque los datos del presente estudio no permiten determinar si la presencia de tapir en los diferentes tipos de hábitat es para alimentarse o para desplazarse, muestran la importancia de los diferentes tipos de vegetación para la especie.
Diversos estudios indican que el tapir hace uso de varios tipos de vegetación donde pueda encontrar alimento, siempre y cuando no sean zonas con mucha presencia humana (Lira-Torres, Naranjo, Güiris y Cruz, 2004; Muench, 2001; Naranjo y Cruz, 1998). Para la RBC, Reyna-Hurtado y Tanner (2007) concluyen que cuando al tapir no se le caza se le encuentra incluso en zonas perturbadas.
Las reservas de Calakmul y Balam Kú tienen vegetación en buen estado de conservación y de forma continua, además, este estudio demuestra que hay disponibilidad de alimentos y agua, y al ser áreas naturales protegidas, la cacería es casi nula o inexistente (Reyna-Hurtado et al., 2010). Estos factores pueden ser una razón de por qué el tapir se desplaza por toda el área en busca de zonas de alimentación o de cuerpos de agua.

