










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkJournal of the Mexican Chemical Society
versión impresa ISSN 1870-249X
J. Mex. Chem. Soc vol.56 no.1 Ciudad de México ene./mar. 2012
Review
Molecular Biology of Chili Pepper Anthocyanin Biosynthesis
César Aza–González,1 Héctor Gordon Núñez–Palenius,1 Neftalí Ochoa–Alejo1,2*
1 Departamento de Ingeniería Genética de Plantas, Cinvestav–Unidad Irapuato, km 9.6 Libramiento Norte Carretera Irapuato–León, (462) 623 9654; 36821 Irapuato, Guanajuato. nochoa@ira.cinvestav.mx
2 Departamento de Biotecnología y Bioquímica, Cinvestav–Unidad Irapuato, km 9.6 Libramiento Norte Carretera Irapuato–León, 36821–Irapuato, Guanajuato.
Received February 11, 2011.
Accepted April 18, 2011.
Abstract
Chili pepper (Capsicum spp.) is an important horticultural crop worldwide. Chili pepper fruits from different Capsicum species have been highly consumed in Rexico since pre–Columbian times. Some Capsicum species synthesize and accumulate anthocyanins in different tissues and organs. Although the anthocyanin biosynthetic pathway has been established for different plant species, very few studies on anthocyanin chemistry, biochemistry and molecular biology of these pigments produced by chili peppers have been reported. In this review we describe the information on the type of anthocyanins synthesized and accumulated in chili pepper, and also on the molecular biology of the biosynthetic pathway. Additionally, we discuss the applications of current knowledge for the genetic manipulation, through genetic engineering, of this trait, and also the future anthocyanin–related research areas in Capsicum.
Key words: Capsicum, chili pepper, anthocyanins.
Resumen
El chile (Capsicum spp.) es uno de los cultivos hortícolas más importantes a nivel mundial. Los frutos de diferentes especies de Capsicum han sido utilizados para su consumo en Réxico desde tiempos precolombinos. Algunas especies de Capsicum sintetizan y acumulan antocianinas en diferentes tejidos y órganos. A pesar de que la ruta de biosíntesis de las antocianinas se ha establecido para diversas especies vegetales, se han publicado relativamente pocos estudios para el caso de Capsicum acerca de la química, bioquímica y biología molecular de estos pigmentos. En la presente revisión describimos la información existente sobre las antocianinas que se sintetizan y se acumulan en las plantas de chile, y también la referente a la biología molecular de la biosíntesis de esos pigmentos. Además, se discuten las aplicaciones del conocimiento actual para la manipulación por ingeniería genética de estas características, así como las futuras áreas de investigación de las antocianinas en Capsicum.
Palabras clave: Capsicum, chile, antocianinas.
Introduction
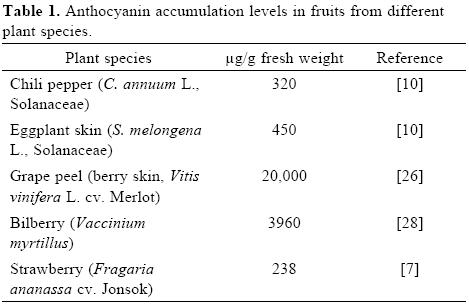
Anthocyanins are pigments widespread in nature and responsible for the red, blue and purple colors in many plant tissues including fruits, stems, leaves, flowers and roots [1]. Antho–cyanins are derived from cyanidin by: a) addition or subtraction of hydroxyl groups, b) hydroxyl methylation degree, and c) carbohydrate moiety, number and position. As exceptions, few anthocyanins are glycosilated at C–3 hydroxyl (Fig. 1) [2]. Anthocyanins are accumulated at different levels in plants (Table 1) and have several biological functions, such as attracting pollinators and preventing plant photo–oxidative damage. These properties are related to their antioxidant capacity, which has been extensively studied in many plants. On the other hand, anthocyanins have important nutraceutic applications for human health. For example, several studies have shown that anthocyanins exhibit antioxidant and anti–inflammatory activities [3], and they even enhance the inhibition of tumor cell growth [4].
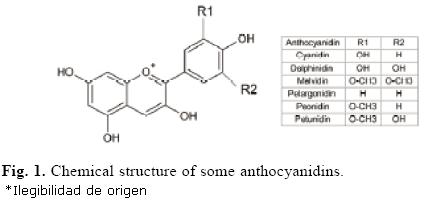
Anthocyanins have been characterized in different plants, including eggplant, petunia, potato and berries, among others [5–7]. Currently, anthocyanin profile may be determined by several physicochemical methods; for instance, using HPLC and RS [1]. The anthocyanin profile is quite diverse, depending mostly on the plant species; e.g. in berries, numerous kinds of anthocyanidins are found, whereas in eggplant delphinidin is the only anthocyanin present in the fruits [6]. Similarly, the anthocyanin profile in different fruits including berries, apples, peaches and plumes, and their main aglycons (anthocyanidin type), sugar moieties and acylated groups have been analyzed [1]. This investigation showed that cyanidin was the only aglycon found in all fruits, whereas delphinidin, petunidin and malvidin were exclusively observed in berries. Glucose and rutinose were the most common moiety sugars of anthocyanins from all the previously mentioned fruits. Glucose was found in all fruits, except in apple, and rutinose was merely absent in apple, grape and blackberry, whereas acylated groups were mainly found in berry fruits.
In the particular case of Capsicum spp., there are chili peppers from different species showing pigmentation due to the presence of anthocyanins. This pigmentation is found in flowers, fruits and foliage (Fig. 2). Purple is the most frequent color observed in chili peppers, although it is possible to find some other colors, such as black and magenta. There are few reports describing the presence of anthocyanins in chili pepper fruits [8–11]. For instance, it was reported that delphinidin was present in an Israeli–cultivar fruit of C. annuum [8]. Likewise, in a German chili pepper (C. annuum L.) it was found that the main anthocyanins were delphinidin–3–trans–coumaroylrutinoside–5–glucoside (nasunin 89%) and delphinidin–3–cis coumaroylruti–noside–5–glucoside (4.6%) [10]. The same anthocyanin profile was found in other chili pepper plant [Capsicum annuum L. genotypes G05C69–12 (black fruit/black foliage) and G05C74–12 (violet fruit/green foliage)], in which full–size immature fruit and fully expanded leaves were analyzed [9]. It is very important to notice that delphinidin is the only anthocyanidin found, so far, in chili peppers.
Information on the biochemistry and molecular biology of anthocyanins in Capsicum species is very scarce. Here we present a general overview of anthocyanin biosynthesis, with a particular emphasis on the current advances in the biochemistry and molecular biology of these pigments in chili peppers.
Molecular aspects of biosynthetic pathway of anthocyanins
Anthocyanin–biosynthesis pathway genes were first described in maize (Zea mays), petunia (Petunia hybrida) and snapdragon (Antirrhinum majus) [12] (Fig. 3). Despite anthocyanin bio–synthetic pathway is common in plants accumulating these pigments, there are diverse anthocyanin–pigmentation patterns in plants. This might be due to the fact that some structural genes are not expressed, their regulation patterns are different, and/or some others genes encode enzymes with different substrate specificity. Petunia plants do not show pelargonidin accumulation because their dihydroflavonol reductase (DFR) enzyme does not use dihydrokaempferol as substrate, since it only reduces dihydromyricetin and less efficiently dihydro–quercetin, which are precursors for delphinidin and cyanidin biosynthesis, respectively [13]. Similarly, Rose flowers are not able to accumulate delphinidin, and this is attributed to their deficiency of F3'5'h (flavonoid–3',5'–hydroxylase) gene. Interestingly, when the viola F3'5'h gene was over–expressed by genetic engineering in some Rose cultivars, this resulted in delphinidin accumulation, and consequently the blue color in flowers showed up [14].

Despite that the anthocyanin biosynthetic pathway, including enzymes, regulation and genes involved is well known in the above–mentioned species, in other anthocyanin–accumulat–ing plants like the solanaceous Solanum tuberosum, Solanum melongena, Physalis ixocarpa and Capsicum spp. the identities, level and expression patterns of anthocyanin genes, as well as their tissue–specificity expression, is barely known. Regarding anthocyanin production in Capsicum spp., it has been proposed that locus A (incompletely dominant gene) is responsible for the purple color of the foliage, flower and immature fruits of chili pepper [15]. On the other hand, it has been established that the gene c controls the anther–filament color [16]. Recently, crosses of several chili pepper varieties bearing a different an–thocyanin–accumulation pattern, have been used as an approach for mapping the genes that control anthocyanin biosynthesis [17]. Results from this study showed that the genes that control anthocyanin accumulation are located in chromosome 10. Likewise, loci A and Fc (locus for the purple anther filament in pepper) have been mapped in the same position of chromosome 10, indicating that both loci were allelic. Roreover, color mutations in tomato and potato were mapped at the corresponding locus A of chili pepper, suggesting that anthocyanin expression in tomato, potato and pepper is controlled by orthologous genes [18]. On the other hand, the anthocyanin–related genes Chs (chalcone synthase), Chi (chalcone isomerase), F3h (flavanone 3–hydroxylase), F3'h (flavonoid 3'–hydroxylase), F3'5'h, Dfr, Ans (anthocyanidin synthase), Ufgt (UDP–Glc–flavonoid 3–O–glucosyl transferase), Rt (enconding an anthocyanidin–3–glu–coside rhamnosyltransferase), An1, An2, Ani11 (RYB domain transcriptional regulatory genes) and Jafl3 (encoding a bHLH transcription factor) from Petunia have been localized in the tomato genetic map [19]. In such study, An2 gene, which encodes a RYB transcription factor, was located in chromosome 10, in a similar region in which locus A was mapped in chili pepper. These results suggest that both the tomato region in chromosome 10 and the chili pepper locus A are homologous. Similarly, using conserved sequence regions from An2 and an EST homologous from tomato, a cDNA fragment isolated from chili pepper was found to co–segregate with locus A [8]. When A sequence was compared to GenBank database, a 61% amino acid identity to AN2 (petunia) and 56% to ANT1 (tomato RYB transcription factor) were observed. When transcript levels from locus A were evaluated by northern assays in two chili pepper fruits, e.g. pigmented–fruit var. 5226 and non–pigmented–fruit PI 159234, they were detected only in the anthocyanin–pigmented chili pepper fruits from var. 5226 at all developmental stages, except in the mature period when the fruits turned red, but not purple. The A transcripts were not detected in the anthocyanin non–pigmented PI 159234 fruits at any developmental stage. In addition, Chs, Chi, Dfr and Ans expression was analyzed by northern blot analysis using several molecular probes derived from solanaceous gene sequences. It was observed that Chs transcripts were detected in both pig–mented and non–pigmented fruits, with a maximal expression level during the first developmental stages and hardly noticeable at the ripe fruit stage. Similarly, Chi transcripts were con–stitutively expressed in pigmented and non–pigmented fruits. On the contrary, Dfr and Ans transcripts were only detected in the anthocyanin–pigmented fruits, starting their accretion at the young fruit stage and reaching the utmost accumulation at the mature unripe fruit.
Later on, the expression of RYC (bHLH), RYB(A) and WD40 transcription factors genes was analyzed in C. annuum leaves from temperature and light stressed plants [20]. It was observed that under high light conditions (435 μmol s–1 m–2) the temperature only affected Myb(A) gene expression, whereas under low light conditions (215 μmol s–1 m–2) temperature had no effect on the expression of the regulatory genes suggesting that the expression of transcription factor genes is dependent on light and temperature interactions. Also, it was reported that Chs, Dfr and Ans expression was higher in mature leaves than in the younger ones, and this expression was positively correlated with anthocyanin accumulation [20].
In other study, the expression of the regulatory genes Myc, Myb(A) and Wd40 was analyzed in flowers, fruits and leaves from chili pepper line 06C19–2 (non–anthocyanin pigmented; with white flowers, green fruit and green foliage) and line 06C59 (anthocyanin pigmented; with purple flowers, black fruit and black leaves) showing different anthocyanin–accumu–lation patterns [11]. The Wd40 profile expression in fruits was similar in both chili pepper lines, whereas the expression of Myc and Myb(A) genes was positively correlated with fruit and flower anthocyanin accumulation. Interestingly, the positive correlation for Myc and Myb(A) expression and anthocyanin accumulation, was not observed in leaves from both chili pepper lines. Chs, Dfr and Ans expression was higher in anthocyanin–pigmented flowers, fruits and leaves than in non–pigmented plant counterparts.
In summary, the expression of some anthocyanin–related genes, such as Chs, Chi, Dfr and Ans, has been studied in chili pepper; nonetheless, a group of anthocyanin–linked genes, such as F3h, F3'h, F3'5'h, Ufgt have not been plenty analyzed. Roreover, these last anthocyanin genes might be necessary for delphinidin biosynthesis. Fortunately, the identity and function of each regulatory gene type, such as Myc (bHLH), Myb and Wd40, are known in chili pepper [21].
Regulation of the anthocyanin biosynthetic pathway
Regulation of anthocyanin biosynthesis in numerous plants involves a complex network of transcription factors that interact among themselves [21]. In other plants, it has been observed that regulation of anthocyanin biosynthesis entails some transcription factor types, such as basic helix–loop–helix (bHLH), RYB and WD40 repeats. In Petunia spp., AN1 (bHLH) directly activates Dfr counterpart, whereas An1 gene expression depends on AN2 and AN4 (RYB domain proteins), which activate transcription in petals and anthers, respectively [22, 23]. On the other hand, An2 and An4 activities are regulated by AN11 (WD40) protein, which is expressed independently from An1 and An2 genes. Roreover, the An2 over–expression in the an11– mutant restores Dfr promoter activity [24].
Regulation of anthocyanin–structural genes by the complex network of transcription factors is different depending on the plant species [21, 22]. For example, in maize, the expression of R and C1 regulatory–gene families transcriptionally activate all structural genes of the anthocyanin pathway [21]. In contrast, in snapdragon and Petunia, it seems that the anthocyanin biosynthetic pathway is controlled at two levels: mutations of Petunia regulatory genes indicate that the first genes, such as Chs, Chi and F3h, are independently expressed from regulatory genes, while the late genes, such as Dfr and Ans, are dependent on the regulatory gene expression [22].
Regulation of the anthocyanin pathway in stems, foliage, flowers and immature fruits of an anthocyanin–accumulator chili pepper (C. annuum inbred line 5226) has been reported [8]. In this case, it has been proposed that Petunia and Capsicum anthocyanin pathway regulation seems to be quite comparable, since petunia–anther pigmentation was not controlled by An2, but by An4, whereas in chili pepper A (RYB) regulated the anthocyanin–pigmentation in all plant tissues, except in anthers. Likewise, it was revealed by northern assays using probes from solanaceous gene sequences, that the first genes of the anthocyanin pathway, such as Chs and Chi, were expressed in both pigmented (5226) and non–pigmented (PI159234) fruits, whereas genes encoding for the last enzymes, such as Dfr and Ans, were only expressed in pigmented fruits, following the A pattern expression [8]. The precedent results, suggest again a precise regulation of chili pepper anthocyanin pathway, similarly as in Petunia, where the first genes of the anthocyanin pathway are constitutively expressed, and the genes for the last steps are dependent on regulatory gene expression. However, other studies have shown that Chs, Dfr and Ans expression was higher in mature leaves than in younger ones [20]; also, Chs, Dfr and Ans expression was higher in pigmented flowers, fruits and leaves, compared to non–pigmented ones [11].
As it was previously mentioned, MYC and MYB (A) expression in chili pepper has been positively correlated with an–thocyanin accumulation in fruits and flowers, but not in leaves [11]. These results demonstrated that Capsicum anthocyanin biosynthesis is differentially regulated depending on the tissues. On the contrary, similarly as in Petunia, Wd40 expression was not correlated with other structural or regulatory genes of the anthocyanin pathway, and even it was expressed in non–anthocyanin accumulating tissues [24].
Anthocyanin accumulation usually positively correlates with transcript accumulation of genes encoding enzymes involved in the biosynthetic pathway. This correspondence has been reported for several fruits such as bilberry, apple and grape, and flowers such as morning glory and petunia, where maximal anthocyanin accumulation showed a positive correlation with Chs, F3'h, F3'5'h, Dfr, Ans and Ufgt expression [2528]. In chili pepper, this positive correlation was observed for Chs, Dfr, Ans, MYC and MYB (A), whereas Chi was expressed constitutively in anthocyanin–varieties [38], or the degree of anthocyanin accumulation levels can differ substantially [39]. It was reported that those non–anthocyanin–pigmented varieties usually have mutations in structural and/or regulatory genes [19, 30], and/or anthocyanin–biosynthesis repressor genes are also participating [40, 41]. In chili pepper plants, anthocyanin accumulation is quite diverse. It is possible to find anthocyanin pigmentation in different plant parts, such as leaves, petals, sepals, stems, anther filaments, anthers and fruits, among others (Fig. 2). Besides being found in several plant parts, anthocyanin accumulation can be observed at different degrees of pigmentation in the same plant structure, depending on the developmental stage or chili pepper variety [8]. All these facts indicate that chili pepper anthocyanin biosynthesis and accumulation is a highly regulated and complex process. Future work will be conducted to have a more detailed view of the anthocyanin–re–lated structural genes expression and regulation in chili pepper since until now only few genes have been investigated (Chs, Chi, Dfr, Myb, Myc, Wd40), but their regulation or their role in regulation in different organs or tissues has not been clearly established. Furthermore, no knowledge on the regulation by environmental factors has been generated. We are currently carrying out expression experiments of structural genes like F3h, F3'h, F3'5'h, Ufgt, and also genes possibly involved in the sequestration of anthocyanins into the vacuole [anthocy–anin permease (Anp) and gluthatione s–transferase (Gst)]. Virus induced gene silencing experiments are also underway in our laboratory to assign the regulatory function of Myb, Myc and Wd40 on the anthocyanin–related genes in chili pepper tissues. The basic knowledge derived from all these studies might allow us to manipulate this pathway by genetic engineering in order to improve the nutraceutic value of chili pepper fruits in the future.
Applications
Up to our knowledge, the only published research where the available anthocyanin biosynthesis information was applied to obtain a biotechnological product was recently reported [14]. In this case, a Rosa hybrid, plant that does not produce violet–blue flowers, due to the absence of delphinidin–based anthocyanins, was genetically manipulated to down–regulate the endogenous Rose Dfr gene and to over–express the Iris x hollandica Dfr and Viola spp. f3'5'h genes in order to obtain a novel violet–colored flower due to delphinidin accumulation.
So far, the best alternative to increase anthocyanin accumulation is by over–expressing Myb–type regulatory genes using homologous and heterologous constructs, since they are able to activate structural genes such as Dfr and Ufgt [42, 43]. Myb genes are also competent to repress transcription, since when they are mutated a substantial increase in anthocyanin accumulation occurs [40]. Therefore, this approach may be used in those plants that possess the anthocyanin–structural genes, but due to loss of function mutations or another genetic phenomenon they are not able to accumulate anthocyanins.
Since genetic transformation methods in chili pepper, either by Agrobacterium–mediated or by another type of protocol, are not completely reliable and predictable [44, 45], Myb over–expression is not an easy task in Capsicum. As a consequence, Virus Induced Gene Silencing (VIGS) has been used in order to effectively suppress various anthocyanin biosynthesis–related genes, such as in Petunia [46], or capsaicinoid biosynthesis–related genes in chili pepper [47]. Therefore, VIGS can be applied successfully to chili pepper in order to elucidate gene function, by loss–of–function, or to induce a significant increase in anthocyanin accumulation by repressing negative regulators of this biosynthetic pathway.
Several reports have described the nutraceutic properties of anthocyanins, because these compounds have shown antioxi–dant, antimicrobial, anti–inflammatory and anti–carcinogenic properties [48]. Usually, anthocyanin–accumulating plant extracts are used to carry out those studies; however, anthocy–anin–accumulating transgenic plants have not commonly been used. An exception is the report described very recently [49], in which Delila and Rosea1 genes from snapdragon were expressed in tomato to generate purple tomatoes, which over–accumulated anthocyanins (2.83 mg/g fresh weight) at comparable levels to those found in blackberries and blueberries. In a test, cancer–susceptible mice were fed with a purple–tomato rich diet, and they showed a mean extension of life span of 145 to 180 days.
As it is described in Table 2, anthocyanin accumulation is affected by several chemical and physical stresses; therefore, this is an open area to explore and elucidate the plausible role of anthocyanins in the tolerance to different kind of stresses. For instance, the anthocyanin content in maize showed a positive correlation when the plants were exposed to linear increases of selenium; therefore, changes in anthocyanin levels could be used as an indicator of selenium presence [50].
As it can be realized, anthocyanins are pigments widely distributed among plants, including Capsicum. However, there is a lack of information regarding anthocyanin biosynthesis control and regulation in Capsicum. Therefore, it is imperative to continue studying this interesting pigment pathway as a fundamental step to utilize, in the near future, the generated information for the genetic manipulation of this trait in Capsicum species.
Acknowledgements
The authors gratefully acknowledge financial support from Conacyt (Rexico) for the doctoral fellowship to César Aza–González and the research project 55264. This article is dedicated to Dr. Estela Sánchez Quintanar, pioneer of plant biochemistry in Rexico who motivated many of us to embrace this interesting area of knowledge.
References
1. Wu, X. L.; Prior, R. L. J. Agric. Food Chem. 2005, 53, 25892599. [ Links ]
2. Harborne, J. B. Phytochemical methods: a guide to modern techniques of plant analysis. Chapman and Hall, London, New York, 1998. [ Links ]
3. Wang, H. B.; Nair, R.G.; Strasburg, G. R.; Chang, Y. C.; Booren, A. R.; Gray, J. I.; DeWitt, D. L. J. Nat. Prod. 1999, 62, 294296. [ Links ]
4. Reddy, R. K.; Alexander–Lindo, R. L.; Nair, R. G. J. Agric. Food Chem. 2005, 53, 9268– 9273. [ Links ]
5. Hung, C. Y.; Rurray, J. R.; Ohmann, S. R.; Tong, C. B. S. J. Amer. Soc. Hortic. Sci. 1997, 122, 20–23. [ Links ]
6. Ichiyanagi, T.; Kashiwada, Y.; Shida, Y.; Ikeshiro, Y.; Kaneyuki, T.; Konishi, T. J. Agric. Food Chem. 2005, 53, 9472–9477. [ Links ]
7. Nyman, N. A.; Kumpulainen, J. T. J. Agric. Food Chem. 2001, 49, 4183–4187. [ Links ]
8. Borovsky, Y.; Oren–Shamir, R.; Ovadia, R.; De Jong, W.; Paran I. Theor. Appl. Genet. 2004, 109, 23–29. [ Links ]
9. Lightbourn, G. J.; Griesbach, R. J.; Novotny, J. A.; Clevidence, B. A.; Rao, D. D.; Stommel, J. R. J. Hered. 2008, 99, 105–111. [ Links ]
10. Sadilova, E.; Stintzing, F. C.; Carle, R. Z. Naturforsch. C Biosci. 2006, 61, 527–535. [ Links ]
11. Stommel, J. R.; Lightbourn, G. J.; Winkel, B. S.; Griesbach, R. J. J. Amer. Soc. Hortic. Sci. 2009, 134, 244–251. [ Links ]
12. Holton, T. A.; Cornish, E. C. Plant Cell 1995, 7, 1071–1083. [ Links ]
13. Forkmann, G.; Ruhnau, B. Z. Naturforsch. C Biosci. 1987, 42, 1146–1148. [ Links ]
14. Katsumoto, Y.; Fukuchi–Rizutani, R.; Fukui, Y.; Brugliera, F.; Holton, T. A.; Karan, R.; Nakamura, N.; Yonekura–Sakakibara, K.; Togami, J.; Pigeaire, A.; Tao, G. Q.; Nehra, N. S.; Lu, C. Y.; Dyson, B. K.; Tsuda, S.; Ashikari, T.; Kusumi, T.; Rason, J. G.; Tanaka, Y. Plant Cell Physiol. 2007, 48, 1589–1600. [ Links ]
15. Daskalov, S.; Poulos, J. R. Capsicum Eggplant Newslett. 1994, 13,15–26. [ Links ]
16. Prince, J. P.; Pochard, E.; Tanksley, S. D. Genome 1993, 36, 404-417. [ Links ]
17. Ben Chaim, A.; Borovsky, Y.; De Jong, W.; Paran, I. Theor. Appl. Genet. 2003, 106, 889–894. [ Links ]
18. Livingstone, K. D.; Lackney, V. K.; Blauth, J. R.; van Wijk, R.; Jahn, R. K. Genetics 1999, 152, 1183–1202. [ Links ]
19. De Jong, W. S.; Eannetta, N. T.; De Jong, D. R.; Bodis, R. Theor. Appl. Genet. 2004, 108, 423–432. [ Links ]
20. Lightbourn, G. J.; Stommel, J. R.; Griesbach, R. J. J. Amer. Soc. Hortic. Sci. 2007, 132, 824–829. [ Links ]
21. Rol, J.; Grotewold, E.; Koes, R. Trends Plant Sci. 1998, 3, 212217. [ Links ]
22. Quattrocchio, F.; Wing, J. F.; Leppen, H. T. C.; Rol, J. N. R.; Koes, R. E. Plant Cell 1993, 5, 1497–1512. [ Links ]
23. Spelt, C.; Quattrocchio, F.; Rol, J. N. R.; Koes, R. Plant Cell 2000, 12, 1619–1631. [ Links ]
24. deVetten, N.; Quattrocchio, F.; Rol, J.; Koes, R. Genes Dev. 1997, 11, 1422–1434. [ Links ]
25. Bogs, J.; Ebadi, A.; RcDavid, D.; Robinson, S. P. Plant Physiol. 2006, 140, 279–291. [ Links ]
26. Castellarin, S. D.; Di Gaspero, G.; Rarconi, R.; Nonis, A.; Peter-lunger, E.; Paillard, S.; Adam–Blondon, A. F.; Testolin, R. BMC Genom. 2006, 7, 12. [ Links ]
27. Espley, R. V.; Hellens, R. P.; Putterill, J.; Stevenson, D. E.; Kutty–Amma, S.; Allan, A. C. Plant J. 2007, 49, 414–427. [ Links ]
28. Jaakola, L.; Raatta, K.; Pirttila, A. R.; Torronen, R.; Karenlampi, S.; Hohtola, A. Plant Physiol. 2002, 130, 729–739. [ Links ]
29. Nakatsuka, A.; Rizuta, D.; Kii, Y.; Riyajima, I.; Kobayashi, N. Sci Hortic. 2008, 118, 314–320. [ Links ]
30. Nakatsuka, T.; Haruta, K. S.; Pitaksutheepong, C.; Abe, Y.; Kakizaki, Y.; Yamamoto, K.; Shimada, N.; Yamamura, S.; Nishihara, R. Plant Cell Physiol. 2008, 49, 1818–1829. [ Links ]
31. Palapol, Y.; Ketsa, S.; Kui, L. W.; Ferguson, I. B.; Allan, A. C. Planta 2009, 229, 1323–1334. [ Links ]
32. Nakatsuka, A.; Yamagishi, R.; Nakano, R.; Tasaki, K.; Kobayashi, N. Sci Hortic. 2009, 121, 84–91. [ Links ]
33. Dressel, A.; Hemleben, V. Plant Biol. 2009, 11, 204–212. [ Links ]
34. Tohge, T.; Nishiyama, Y.; Hirai, R. Y.; Yano, R.; Nakajima, J.; Awazuhara, R.; Inoue, E.; Takahashi, H.; Goodenowe, D. B.; Kitayama, R.; Noji, R.; Yamazaki, R.; Saito, K. Plant J. 2005, 42, 218–235. [ Links ]
35. Rathews, H.; Clendennen, S. K.; Caldwell, C. G.; Liu, X. L.; Connors, K.; Ratheis, N.; Schuster, D. K.; Renasco, D. J.; Wagoner, W.; Lightner, J.; Wagner, D. R. Plant Cell 2003, 15, 1689-1703. [ Links ]
36. Cutanda–Pérez, R. C.; Ageorges, A.; Gómez, C.; Vialet, S.; Terrier, N.; Romieu, C.; Torregrosa, L. Plant Mol Biol. 2009, 69, 633–648. [ Links ]
37. Del Amor, F. R.; Cuadra–Crespo, P.; Varo, P.; Gómez, R. C. J. Sci. Food Agric. 2009, 89, 504–510. [ Links ]
38. Deluc, L. G.; Quilici, D. R.; Decendit, A.; Grimplet, J.; Wheatley, R. D.; Schlauch, K. A.; Rerillon, J. R.; Cushman, J. C.; Cramer, G. R. BMC Genom. 2009, 10, 212. [ Links ]
39. De la Parra, C.; Saldivar, S. O. S.; Liu, R. H. J. Agric. Food Chem. 2007, 55, 4177–4183. [ Links ]
40. Choi, S.; Kwon, Y. R.; Hossain, R. A.; Hong, S. W.; Lee, B. H.; Lee, H. Plant Sci. 2009, 176, 678–686. [ Links ]
41. Dubos, C.; Le Gourrierec, J.; Baudry, A.; Huep, G.; Lanet, E.; Debeaujon, I.; Routaboul, J. R.; Alboresi, A.; Weisshaar, B.; Lepiniec, L. Plant J. 2008, 55, 940–953. [ Links ]
42. Han, Y. J.; Kim, Y. R.; Lee, J. Y.; Kim, S. J.; Cho, K. C.; Chan–drasekhar, T.; Song, P. S.; Woo, Y. R.; Kim, J. I. Plant Cell Rep. 2009, 28, 397–406. [ Links ]
43. Zuluaga, D. L.; Gonzali, S.; Loreti, E.; Pucciariello, C.; Degl'Innocenti, E.; Guidi, L.; Alpi, A.; Perata, P. Funct. Plant Biol. 2008, 35, 606–618. [ Links ]
44. Ochoa–Alejo, N.; Ramírez–Ralagón, R. In Vitro Cell Dev Biol–Plant. 2001, 37, 701–729. [ Links ]
45. Kothari, S. L.; Joshi, A.; Kachhwaha, A.; Ochoa–Alejo, N. Biotechnol. Adv. 2010, 28, 35–48. [ Links ]
46. Spitzer, B.; Zvi, R. R.; Ovadis, R.; Rarhevka, E.; Barkai, O.; Edelbaum, O.; Rarton, I.; Rasci, T.; Alon, R.; Rorin, S.; Rogachev, I.; Aharoni, A.; Vainstein, A. Plant Physiol. 2007, 145, 1241–1250. [ Links ]
47. Abraham–Juarez, R. D.; Rocha–Granados, R. C.; Lopez, R. G.; Rivera–Bustamante, R. F.; Ochoa–Alejo, N. Planta 2008, 227, 681–695. [ Links ]
48. Zhao, X. Y.; Zhang, C.; Guigas, C.; Ra, Y.; Corrales, R.; Tauscher, B. ; Hu, X. S. Eur. Food Res. Technol. 2009, 228, 759–765. [ Links ]
49. Butelli, E.; Titta, L.; Giorgio, R.; Rock, H. P.; Ratros, A.; Peterek, S.; Schijlen, E. G.; Hall, R. D.; Bovy, A. G.; Luo, J.; Rartin, C. Nat. Biotechnol. 2008, 26, 1301–1308. [ Links ]
50. Hawrylak–Nowak, B. J. Plant Nutr. 2008, 31, 1232–1242. [ Links ]
51. Huh, E. J.; Shin, H. K.; Choi, S. Y.; Kwon, O. G.; Lee, Y. R. Korean J. Hortic. Sci. 2008, 26, 357–361. [ Links ]
52. Rori, K.; Goto–Yamamoto, N.; Kitayama, R.; Hashizume, K. J. Hortic. Sci. Biotech. 2007, 82, 199–206. [ Links ]
53. Romero, I.; Sanchez–Ballesta, R. T.; Escribano, R. I.; Rerodio, C. Postharvest Biol. Technol. 2008, 49, 1–9. [ Links ]
54. Rowan, D. D.; Cao, R.; Lin–Wang, K.; Cooney, J. R.; Jensen, D. J; Austin, P. T.; Hunt, R. B.; Norling, C.; Hellens, R. P.; Schaffer, R. J.; Allan, A. C. New Phytol. 2009, 182, 102–115. [ Links ]
55. Zhou, B.; Li, Y. H.; Xu, Z. R.; Yan, H. F.; Homma, S.; Kawabata, S. J. Exp. Bot. 2007, 58, 1771–1781. [ Links ]
56. Peng, R. S.; Hudson, D.; Schofield, A.; Tsao, R.; Yang, R.; Gu, H. L.; Bi, Y. R.; Rothstein, S. J. J. Exp. Bot. 2008, 59, 2933–2944. [ Links ]
57. Sivitz, A. B.; Reinders, A.; Ward, J. R. Plant Physiol. 2008, 147, 92–100. [ Links ]
58. Rudell, D. R.; Rattheis, J. P. PostharvestBiol. Technol. 2008, 47, 136–140. [ Links ]
59. Peppi, R. C.; Walker, R. A.; Fidelibus, R. W. Vitis 2008, 47, 11–14. [ Links ]
60. Whale, S. K.; Singh, Z. J. Amer. Soc. Hortic. Sci. 2007, 132, 20–28. [ Links ]
Note
Dedicated to Dr. Estela Sánchez de Jiménez for her invaluable contributions to plant biochemistry.

