










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkTropical and subtropical agroecosystems
versión On-line ISSN 1870-0462
Trop. subtrop. agroecosyt vol.14 no.3 Mérida sep./dic. 2011
Revisión
Estacionalidad reproductiva de la oveja en México
Reproductive seasonality of sheep in Mexico
J. Arroyo*
Universidad del Mar, Campus Puerto Escondido, km 1.5 Carretera Puerto Escondido - Vía Sola de Vega, Puerto Escondido 71980, Oaxaca, México. *E-mail: arroyo@zicatela.umar.mx
* Corresponding author
Submitted February 21, 2011
Accepted April 15, 2011
Revised received June 07, 2011
Resumen
Con el propósito de discutir y analizar la información disponible, referente al comportamiento reproductivo estacional de los ovinos en la República Mexicana, se realizó la presente revisión. Se analizaron las bases neuroendocrinas que regulan el ciclo reproductivo anual en la oveja y posteriormente se discutió el grado de estacionalidad reproductiva en ovejas criollas de lana, en razas originarias de latitudes altas y en ovejas de pelo, principalmente, en ovejas Pelibuey. Las ovejas criollas de lana, muestran un anestro estacional corto con tendencia a ovular todo el año. Las hembras de origen septentrional, expresan actividad reproductiva estacional, similar a la observada en los individuos ubicados geográficamente en latitudes mayores a 35°. Las ovejas Pelibuey, muestran comportamiento reproductivo anual variable, con reducido anestro estacional o ausencia del mismo. Se sugiere que los mecanismos neuroendocrinos que regulan el anestro estacional en los ovinos, se encuentran activos en las ovejas de origen septentrional que habitan en México; de manera contraria, no se activan en ovejas criollas y de pelo.
Palabras Clave: Estacionalidad reproductiva; anestro estacional; ovejas; neuroendocrinología; México.
Abstract
In order to discuss and analyze the available information concerning the seasonal breeding behavior of sheep in Mexico, this review was conducted. We analyzed the neuroendocrine basis that modulate the annual reproductive cycle in sheep and then discussed the degree of reproductive seasonality in Creole sheep wool, breeds originating in high latitudes and hair sheep, mainly in Pelibuey ewes. The Creole sheep wool show continuous annual reproductive activity and short seasonal anestrous. The females of northern origin, express seasonal reproductive activity, similar to that observed in individuals geographically located at latitudes above 35°. Pelibuey sheep show variable annual reproductive behavior with reduced anestrus or lack thereof. It is suggested that the neuroendocrine mechanisms regulating seasonal anestrus in ewes, are active in the sheep of northern origin that live in Mexico, in a manner contrary is not activated in Creole and hair sheep.
Key Words: Reproductive seasonality; seasonal anestrous; sheep; neuroendocrinology; México.
INTRODUCCIÓN
La estacionalidad de la reproducción, como parte del proceso de selección natural, es un mecanismo de adaptación desarrollado por algunos mamíferos silvestres como estrategia para minimizar el impacto negativo del ambiente (temperatura, humedad y disponibilidad de alimento) en la supervivencia de las crías (Lincoln y Short, 1980; Karsch et al, 1984; Malpaux et al., 1996), de manera que los nacimientos ocurren en la época más favorable del año, con abundancia de pastos y temperatura ambiental confortable. Los ovinos presentan anualmente dos etapas fisiológicas bien definidas (Barrell et al., 1992). Una fase de anestro estacional (días largos), con ausencia de ciclos estrales regulares, receptividad sexual y ovulación; en el macho, cesa la espermatogénesis y la libido. La otra etapa fisiológica, conocida como época reproductiva (días cortos), se caracteriza por la ocurrencia de ciclicidad estral, conducta de estro y ovulación en la hembra; en el macho, se restablece la espermatogénesis y el deseo sexual (Hafez et al, 1952; Legan y Karsch, 1979; Karsch et al, 1984; Malpaux et al, 1997). El fotoperiodo es el factor ambiental primario que regula estos eventos. La oveja posee un sistema neurofisiológico capaz de transformar la señal luminosa en una señal hormonal a través de la síntesis de melatonina y de esta manera detecta las variaciones anuales en la duración del fotoperiodo (Williams y Helliwell, 1993; McMillen et al, 1995; Arendt, 1998; Malpaux et al, 2002). El origen de la raza determina el comportamiento reproductivo estacional; por lo tanto, las razas originarias de latitudes altas (>35°) presentan una marcada estacionalidad reproductiva (Hafez, 1952; Legan y Karsch,1979; Karsch et al, 1984; Robinson y Karsch, 1984; Robinson et al., 1985; Malpaux et al., 1987) y los ovinos de origen mediterráneo o ecuatorial, expresan estacionalidad reproductiva reducida y en ocasiones inexistente (Porras, 1999; Cerna et al., 2000; Valencia et al., 2006; Arroyo et al, 2007).
En México, los primeros estudios referentes a la estacionalidad reproductiva se realizaron en la década de los 70 (Castillo et al, 1972; Valencia et al, 1975; Valencia et al, 1978). El núcleo ovino nacional en la región central y norte de la República Mexicana lo integró inicialmente la oveja criolla de lana (De Lucas-Tron, 2000). La introducción de ovinos de pelo en la región sur del país y el establecimiento de sistemas tecnificados de crianza con razas puras en varias regiones de México, generó variaciones importantes en la genética del ovino nacional (De Lucas-Tron, 2000); lo cual, posiblemente provocó diferencias en su ciclo reproductivo anual. La ausencia de anestro estacional es una ventaja productiva que permite desarrollar programas reproductivos todo el año, sin utilizar fármacos hormonales u otras estrategias, como la bioestimulación sexual. El propósito de la presente revisión es analizar las bases neuroendocrinas que regulan la estacionalidad reproductiva en los ovinos, caracterizar el ciclo reproductivo anual de la oveja en la República Mexicana, establecer con base en lo anterior, su grado de estacionalidad y explicar las posibles causas de esta característica biológica.
CONTROL NEUROENDOCRINO DEL CICLO REPRODUCTIVO ANUAL
La estacionalidad reproductiva en la oveja, condujo al desarrollo de mecanismos especializados en la detección de señales ambientales que permiten determinar el momento óptimo para la reproducción. De todos los factores ambientales, el fotoperiodo es el más repetible y con variabilidad nula entre años. Por lo tanto, la duración de las horas luz, sincroniza el ciclo reproductivo anual de la oveja. Los ovinos detectan las variaciones anuales en la duración del fotoperiodo, utilizan una compleja red neural a nivel central y transforman la señal luminosa en una señal hormonal a través de la síntesis y secreción de melatonina (Bittman y Karsch, 1984; Malpaux et al, 1996; Malpaux et al, 1997; Mapaux et al, 1999; Barrell et al, 2000). En este mecanismo, la luz es captada en el ojo, a través de la retina, la señal luminosa se transforma en una señal eléctrica que es conducida de la retina al hipotálamo por medio del tracto retinohipotalámico; en el hipotálamo, el núcleo supraquiasmático capta la señal y posteriormente se transfiere al núcleo paraventricular; finalmente al cerebro posterior, específicamente al ganglio cervical superior (Arent, 1998). En este punto, la señal eléctrica se transforma en una señal química; el ganglio cervical superior libera noradrenalina, la cual es captada por receptores alfa y beta adrenérgicos en la membrana celular de los pinealocitos, se induce la síntesis de la N-acetil-transferasa, enzima fundamental en la síntesis de melatonina (Arent, 1998); de esta manera, la hormona se sintetiza en los pinealocitos de la glándula pineal durante las horas de oscuridad a partir del aminoácido triptofano (McMillen et al, 1995; Malpaux et al, 2002; Rosa y Bryant, 2003). La menor duración en la secreción de melatonina durante los días largos, permite la síntesis de dopamina e induce el anestro estacional. Durante los días cortos, la mayor duración en la síntesis y secreción de melatonina inhibe la producción de dopamina, con el subsecuente restablecimiento de la actividad estral y la ovulación (Viguié et al., 1997; Malpaux et al, 1999).
Influencia de la Kisspeptina y la Hormona Inhibidora de Gonadotropinas (GnIH) en el Ciclo Reproductivo Anual de la Oveja
Recientemente se descubrieron dos péptidos RF-amida (péptidos con una terminal-C similar, arg-fen-NH2), la kisspeptina (ó metastina) y la GnIH (ó RFRP) (Oakley et al, 2009; Hameed et al, 2011). Las kispeptinas son una familia de neuropéptidos codificados por el gen Kissl, son ligandos endógenos del receptor asociado a la proteína G, Kiss Ir, también conocido como receptor GPR54 (Hameed et al, 2011). Los conocimientos actuales demuestran que la kisspeptina participa de manera determinante en la activación del eje reproductivo y es fundamental para el inicio de la pubertad (Meza-Herrera et al, 2010; Hameed et al, 2011). La kisspeptina estimula la secreción de gonadotropinas en roedores y ovejas, con un efecto directo en las neuronas GnRH, las cuales, expresan el receptor Kiss Ir. En la oveja, las neuronas kisspeptina se localizan en el núcleo arcuato (ARC; también conocido como núcleo A12) y en el área preóptica (Pompólo et al, 2006; Smith et al, 2007) y parece ser determinante en la ocurrencia del pico preovulatorio de LH (Estrada et al, 2006; Clarkson et al, 2008). Por otro lado, se sugiere que la expresión hipotalámica del gen Kissl se regula de acuerdo con el ciclo reproductivo anual en hámsteres y ovejas (Greives et al, 2007; Smith etal, 2007).
Por otro lado, la GnIH se identificó inicialmente en el sistema hipotálamo-hipofisiario de la codorniz, se determinó que inhibe la síntesis y secreción de gonadotropinas (Ubuka et al, 2008). Se sugiere que el efecto inhibitorio de este neuropéptido se facilita porque las neuronas GnIH tienen contacto con las neuronas GnRH en aves y mamíferos (Greives et al, 2008); de manera adicional, se identificaron receptores para GnIH en las neuronas GnRH (Bentley et al, 2003; Oakley et ai, 2009). El péptido GnIH y su RNAm se localizan en el núcleo dorsomedial del hipotálamo (DMH) en hámsteres y ovejas (Himuna et al., 2000; Smith et al., 2008).
En este contexto, Smith et al. (2008) evaluaron la expresión de la Kisspeptina y la GnIH (la proteína y su RNAm) en el cerebro de ovejas Blackface ovariectomizadas tratadas con un implante subcutáneo de estradiol y sin tratamiento esteroide, durante las épocas reproductiva y de anestro estacional. Las ovejas ovariectomizadas con implante de estradiol tuvieron una mayor (P<0.05) expresión del RNAm-Kissl y Kisspeptina en el núcleo arcuato y menor (P<0.05) expresión de la GnIH en el núcleo dorsomedial del hipotálamo durante la época reproductiva en comparación con el anestro; en adición, se observó un mayor (P<0.05) número de terminales neuronales kisspeptina hacia las neuronas GnRH y menos contacto entre las neuronas GnIH-GnRH en la época reproductiva, en relación con la época de anestro estacional. Hubo efecto del estradiol en la inhibición de la expresión de RNAm para Kissl y la proteína Kisspeptina durante el anestro estacional. Se sugiere que los dos peptidos RF-amida (Kisspeptina y GnIH) actúan en sincronía, con efectos opuestos para regular la actividad de las neuronas GnRH entre épocas, de manera que modulan los cambios anuales en la fertilidad y participan en la transición cíclica estacional de la época reproductiva al anestro estacional. Los autores proponen tres hipótesis (Smith et al., 2008) para explicar los mecanismos a través de los cuales, las variaciones anuales en la expresión de Kisspeptina y GnIH, el sistema dopaminérgico, el estradiol y la melatonina, actúan en conjunto y regulan la conducta reproductiva anual de la oveja. La primera teoría sugiere que las neuronas del sistema dopaminérgico, inhiben la GnRH al estimular las neuronas GnIH y el incremento en la concentración de este péptido, bloquea las neuronas Kisspeptina y al disminuir su síntesis, se reduce la actividad de las neuronas GnRH, lo cual conduce al anestro estacional. La segunda teoría propone que el estradiol actúa de manera directa en las neuronas Kisspeptina del ARC, reduce la síntesis de la neurohormona e inhibe la síntesis y secreción de GnRH. La tercera posibilidad sugiere que la melatonina actúa directamente en las neuronas Kisspeptina y GnIH, regula su actividad a través del año y durante los días cortos, estimula la síntesis y secreción de Kisspeptina e inhibe la GnIH; de manera contraria, durante los días largos, reduce la síntesis y secreción de Kisspeptina e induce la síntesis y liberación de GnIH, modulando de esta manera el ciclo reproductivo anual de la oveja.
Factores nutricionales implicados en la reproducción
La influencia de la nutrición en la reproducción se ha investigado extensamente. De manera general se concluyó que la secreción de GnRH se reduce en animales desnutridos (Tatman et al., 1990; Schillo 1992; Keisler et al., 1996; Snyder et al., 1999; Wade y Jones, 2004). Sin embargo, el mecanismo a través del cual, las señales metabólicas generadas por una nutrición deficiente son captadas a nivel central para regular la secreción de GnRH, es complejo y no se ha establecido de manera precisa. Se han estudiado distintos indicadores metabólicos que participan en este proceso, tales como la glucosa, ácidos grasos volátiles, algunos aminoácidos y ácidos grasos no esterificados (Keisler et al., 1996). También se investigaron mediadores endocrinos entre el estado nutricional y los procesos reproductivos; entre ellos, el factor de crecimiento asociado a la insulina (IGF-I), la hormona del crecimiento, la colesistoquinina, el neuropéptido Y (NPY), los peptidos opioides endógenos y su relación con la insulina (Keisler et al., 1996). La glucosa regula la liberación de GnRH y, al parecer, los peptidos asociados a la insulina participan en el control del metabolismo de energía en el cerebro (Werner et al., 1989). Snyder et al. (1999), demostraron que en ovejas con una condición corporal baja, ovariectomizadas y tratadas con implantes subcutáneos de estradiol, se reduce el IGF-I, lo cual inhibe el incremento de secreción de LH asociado con el inicio de la época reproductiva. Por lo tanto, una nutrición inadecuada puede prolongar el anestro estacional.
Otro péptido que puede integrar las señales metabólicas generadas por el estado nutricional con el eje reproductivo es la leptina; hormona que, se sugiere, juega un papel clave en la nutrición, el metabolismo y la endocrinología reproductiva (Blache et al., 2000). La proteína, el gen que la codifica y los receptores para la hormona se identificaron en ovinos (Dyer et al., 1997). En esta especie, como en otras, la producción de leptina se asocia con la masa de tejido adiposo; esta hormona pasa de la circulación sistémica al fluido cerebroespinal y posteriormente a los núcleos hipotalámicos, donde puede afectar el apetito y de alguna manera la secreción de GnRH (Blache et al., 2000).
Por otro lado y como se discutió previamente, es claro que el sistema kisspeptina estimula el eje reproductivo en la oveja (Smith, 2006); se sabe que las neuronas kisspeptina en el cerebro responden a señales metabólicas y pueden transmitir información relevante a las células GnRH, por ejemplo el ARNm para Kissl se reduce en ratas adultas con restricción alimenticia (Brown et al., 2008). Backholer et al. (2010), realizaron un estudio en ovejas Corriedale, subalimentadas con reducido peso corporal y con alimentación y peso corporal adecuados, con el propósito de establecer si las células kisspeptina expresan el receptor Ob-Rb para leptina y reponden por lo tanto a esta hormona, de manera adicional se determinó si existen conexiones recíprocas entre las células kisspeptina, proopiomelanocortina (POMC) y NPY, para modular las funciones metabólica y reproductiva. Los resultados mostraron que la leptina regula las células kisspeptina en el núcleo ARC y el APO, regiones donde se expresa el receptor Ob-Rb. Se observaron conexiones recíprocas complejas entre las células kisspeptina del ARC y las células POMC, formando una red neural a través de la cual las señales metabólicas y las señales de los esteroides sexuales pueden afectar ambos tipos de células; de esta manera, se puede regular la reproducción y la función metabólica. Al parecer, las conexiones recíprocas entre el NPY y las células kisspeptina pueden ser de menor importancia.
Lo anterior es relevante, porque previamente se determinó que el NPY estimula el consumo de alimento e inhibe la reproducción en la oveja (Clarke et al, 2005) y las melanocortinas (como la POMC) reducen el consumo de alimento y estimulan la reproducción en esta especie (Watanobe et al., 1999); entonces, el conocimiento generado en el estudio de Backholer et al. (2010) puede explicar uno de los mecanismos neuroendocrinos y fisiológicos que integran la nutrición con la reproducción.
Factores Sociales Implicados en la Reproducción
La comunicación social entre machos y hembras puede modificar su estado reproductivo (O'Callaghan et al., 2004). Se ha documentado ampliamente que la exposición repentina de hembras anéstricas (anestro estacional) a un macho sexualmente activo, incrementa rápidamente la frecuencia de pulsos de LH (Álvarez y Zarco, 2001; Delgadillo et al., 2008) y la ovulación ocurre entre 40 y 50 h después de la primera exposición; ambos eventos, en la mayoría de los casos, se acompañan por conducta estral (Hulet, 1966; Flores et al, 2000; Álvarez y Zarco, 2001; Delgadillo et al, 2008). El efecto ejercido por los carneros en el sistema reproductivo de las ovejas, es mediado por feromonas presentes en la lana y la cera de lana de los carneros (Knight y Lynch, 1980; Okamura y Mori, 2005).
Wayne et al. (1989), realizaron un estudio con ovejas Suffolk pinelectomizadas, ovariectomizadas con implante de estradiol, alojadas con ovejas y machos intactos, y demostraron que las señales sociales emitidas por estos animales pueden sincronizar la actividad reproductiva estacional en ovejas sin la capacidad de responder a señales fotoperiodicas. O'Callaghan et al. (2004) demostraron un efecto crónico de la presencia del carnero en el ciclo reproductivo anual de la oveja. Observaron que el final de la época reproductiva se retrasa 43 días por la exposición continua del macho, durante 2 meses a partir del solsticio de invierno. Estos resultados demostraron que el momento de transición de la época reproductiva al anestro, en condiciones naturales, no sólo se regula por el fotoperiodo, existen otras señales involucradas en la sincronización de la época reproductiva en ovejas y la importancia relativa de estas señales puede cambiar a través del año (O'Callaghan et al, 2004). En razas de origen ecuatorial, las señales sociales parecen ser también importantes. Por ejemplo, la detección diaria de celos durante la época de días largos (dic-jun; 19° Lat. N), con carneros adultos provistos con mandil, parece estimular la actividad estral en ovejas Pelibuey (Valencia et al, 2006). Las evidencias anteriores sugieren que, si bien el fotoperiodo es la señal ambiental primaria que sincroniza el ciclo reproductivo anual de la oveja, las señales sociales en épocas específicas del año (transición fisiológica), pueden regular la actividad sexual de la oveja.
Época Reproductiva
La estación reproductiva en la oveja, ocurre durante la época de días cortos y se caracteriza por la presencia de ciclos estrales regulares, conducta de estro y ovulación (Rawlings et al, 1977; Legan y Karsch, 1979), en el hemisferio norte, se presenta entre los meses de agosto a enero, pero varía de acuerdo con la raza y ubicación geográfica (Hafez et al, 1952; Legan y Karsch, 1979; Karsch et al, 1984; Malpaux et al, 1997). El ciclo estral de los ovinos tiene una duración aproximada de 17 días. En la fase lútea, que comprende el metaestro y diestro, la concentración de progesterona alcanza valores de 1 ng ml-1 o más, esta hormona se sintetiza y libera a partir de un cuerpo lúteo maduro y funcional. La progesterona ejerce un efecto de retroalimentación negativa a nivel hipotalámico e inhibe la secreción pulsátil de GnRH y por lo tanto, de LH (Figura 1). De manera específica, la progesterona actúa a nivel del área preóptica (APO), en donde activa las neuronas GABA e induce la síntesis de este neurotransmisor, el cual actúa en las neuronas productoras de GnRH e inhibe la síntesis de esta hormona (Robinson y Kendrick, 1992; Robinson, 1995; Evans et al, 2002; Jackson y Kuehl, 2002). En este evento, se sugiere también la participación de los péptidos opioides endógenos, neurotransmisores que se sintetizan principalmente en el núcleo hipotalámico A12 y en condiciones fisiológicas específicas (principalmente durante el anestro posparto) inhiben la frecuencia de pulsos de GnRH/LH (Arroyo et al, 2009); sin embargo, su intervención durante la fase lútea del ciclo estral, aún debe confirmarse (Goodman et al, 2002).
Durante la fase folicular (proestro y estro), la concentración de P4 es basal, como consecuencia de la lisis del cuerpo lúteo, inducida por la PGF2α; los folículos ováricos crecen y maduran hasta alcanzar un estado preovulatorio (Padilla et al, 1988; Barrell et al., 1992). La síntesis de estradiol en las células de la granulosa aumenta progresivamente, lo cual induce un incremento de esta hormona esteroide en la circulación periférica y actúa de manera directa en las neuronas GnRH a nivel del núcleo ventromedial, el cual se localiza en el área hipotalámica mediobasal e induce el pico preovulatorio de GnRH/LH y 24 horas después, la ovulación (Figura 2). En esta etapa fisiológica, el estradiol ejerce un efecto de retroalimentación positiva (Legan y Karsch, 1979; McNatty et al, 1984; Noel et al., 1993; Ravindra y Rawlings, 1997; Souza et al., 1997; Bartlewski et al., 2000; Evans et al, 2000).
Anestro Estacional
El anestro estacional en la oveja se caracteriza por la ausencia de ciclos estrales regulares, conducta de estro y ovulación; ocurre durante los días largos, entre los meses de febrero y agosto, en el hemisferio norte, cuando la duración en la secreción de melatonina es menor; su amplitud varía de acuerdo con la ubicación geográfica (latitud) y la raza (Yeates, 1949; Legan y Karsch, 1979; Lincoln and Short, 1980; Karsch et al, 1984; Lehman et al, 2002; Thiéry et al, 2002). En esta etapa fisiológica, el estradiol, cuya concentración es basal, ejerce un efecto de retroalimentación negativa a nivel hipotalámico, actúa específicamente en el núcleo dopaminérgico Al5, donde induce la síntesis y secreción de dopamina, la cual actúa en las neuronas productoras de GnRH e inhibe la frecuencia de síntesis y liberación de esta hormona (Thiéry et al, 1989; Gayrard et al, 1994; Havern et al, 1994; Thiéry et al, 1995; Gallegos-Sánchez et al, 1997; Viguié et al, 1997). El evento fisiológico anterior ocurre a pesar de que en el núcleo Al5 no se identificaron receptores para estradiol (Lehman y Karsch, 1993; Goubillon et al, 1999); por lo tanto, el mecanismo de acción del estradiol en este proceso no es claro (Thiéry et al, 1995).
De manera reciente (Bogusz et al, 2008) determinaron que GABA inhibe la secreción de dopamina y se identificaron procesos neuronales GABA aferentes al núcleo A15, provenientes del área preóptica y se demostró que durante el anestro estacional, el estradiol suprime la liberación de GABA, este efecto inhibitorio ocurre en el núcleo Al5; específicamente, en los procesos neurales mencionados. La supresión en la liberación de GABA, activa las neuronas dopaminérgicas e incrementa la síntesis y secreción de dopamina, la cual ejerce su efecto biológico en las neuronas GnRH y reduce la frecuencia de pulsos de esta hormona y por lo tanto de LH (Figura 3).
ESTACIONALIDAD REPRODUCTIVA EN OVEJAS DE LANA
Los estudios desarrollados con el propósito de conocer el comportamiento reproductivo estacional en ovinos de lana en la República Mexicana, son limitados. Existen dos metodologías principales para estudiar la estacionalidad reproductiva en ovejas. La primera, relativamente sencilla, consiste en la detección de celos utilizando machos con mandil, vasectomizados, con el pene desviado, orquiectomizados con implante de testosterona o bien, hembras androgenizadas. La segunda metodología radica en colectar muestras sanguíneas una o dos veces por semana y determinar la concentración de progesterona plasmática. Concentraciones mayores o iguales a 1 ng/ml indican actividad ovulatoria; las ovejas en anestro presentan concentraciones de progesterona menores a 1 ng/ml (Porras, 1999; Cerna et al, 2000; Arroyo et al, 2007).
Actividad Estral
Valencia et al. (1978) utilizaron 19 ovejas Dorset y 6 criollas adultas con el propósito de estudiar la variación anual en la presentación de estros en la región central de la República Mexicana (19° Lat. N), la detección de celos se realizó con machos vasectomizados. En las ovejas Dorset, las observaciones se realizaron de octubre de 1976 a septiembre de 1977; en las ovejas criollas, la proporción de hembras en calor sólo se evaluó de abril a septiembre de 1977 (6 meses). Para la detección de estros, los machos se introdujeron diariamente al corral de las hembras por 30 minutos. Cuando una hembra se detectó en celo, se retiró del corral con el propósito de permitir que el macho localizara a otras hembras en calor. Se observó una alta proporción de ovejas Dorset ciclando en los meses de octubre a febrero y de junio a septiembre, con una disminución importante (P<0.01) en los meses de marzo, abril y mayo. Las ovejas criollas mostraron una tendencia cíclica continua durante el periodo de observación, con una ligera disminución en el mes de abril, la cual no fue diferente (P>0.05) de la observada en las ovejas Dorset durante el mismo mes.
Posteriormente, Valencia et al. (1980), desarrollaron un estudio en la región central de la República Mexicana (19° Lat. N), utilizaron 6 ovejas criollas adultas en condiciones de estabulación y determinaron la actividad estral a partir de abril de 1977, hasta marzo de 1978. La detección de celos se realizó con machos vasectomizados de acuerdo con la metodología descrita por Valencia et al. (1978). A pesar del limitado número de observaciones, una alta proporción de ovejas cicló los 12 meses de estudio. Dos ovejas no presentaron estro en el mes abril; sin embargo, el 100% mostró celos en mayo. De mayo a julio y de octubre a febrero, todas las ovejas presentaron estro. En agosto y septiembre, una oveja no manifestó celo y en marzo, una hembra distinta no mostró estro. Las ovejas criollas en confinamiento, expresaron una tendencia cíclica anual continua, lo cual representa un importante potencial productivo para estos ovinos, que posiblemente, aun en la actualidad, no se ha considerado.
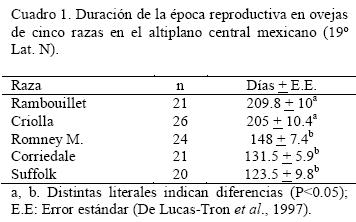
Por su parte, De Lucas-Tron et al. (1997), determinaron la actividad estral anual en cinco razas ovinas en el altiplano central de la República Mexicana (19° Lat. N). Las razas estudiadas fueron la Rambouillet, criolla, Romney Marsh, Corriedale y Suffolk. El autor mencionó que las razas Romney Marsh, Corriedale y Suffolk mostraron estacionalidad reproductiva definida, con un periodo de actividad estral de cuatro a cinco meses y anestro en los meses restantes. De manera contraria, las ovejas Rambouillet y criolla presentaron actividad estral prácticamente todo el año. La duración de la época reproductiva fue similar entre las razas Rambouillet y criollas, pero diferente (P<0.05) de las razas restantes (Cuadro 1).
Actividad Ovulatoria
Existen pocos estudios que evalúan la actividad ovulatoria anual a través de la concentración hormonal (progesterona) en ovejas de lana, ya sea criollas o de raza en la República Mexicana. Un tipo de ovejas poco estudiado por su ubicación, netamente regional es la llamada "Oveja Chiapas", su crianza se desarrolla en la región montañosa de Los Altos de Chiapas (16° Lat. N), principalmente por grupos indígenas, tales como los Tzotziles (Perezgrovas y Castro, 2000). Esta raza, puede considerarse criolla, ya que tiene su origen a partir de las razas españolas Churra, Manchega, Lacha, Castellana y Canaria, de las cuales aún conserva características fenotípicas (Perezgrovas y Castro, 2000). Se han desarrollado algunos ensayos con el propósito de estudiar su ciclo reproductivo anual. Pedraza et al. (1992) evaluaron la actividad ovulatoria (muestreo sanguíneo para determinar concentración de progesterona) en estas ovejas por un periodo de 12 meses y mencionaron que se comportan de manera estacional, mostrando actividad reproductiva de junio a octubre y anestro estacional de noviembre a mayo. Posteriormente, Perezgrovas y Castro (2000), observaron el mismo comportamiento estacional en estas ovejas; sin embargo, reportaron una concentración de progesterona en hembras supuestamente en anestro de 3.92±0.37 y de 5.03±0.31 ng/ml. Ambos valores, de acuerdo con varios autores (Porras, 1999; Cerna et al., 2000; Valencia et al., 2006; Arroyo et al., 2007), corresponden a ovejas con actividad ovulatona, en las cuales existen cuerpos lúteos funcionales; por otro lado, concentraciones menores a 1 ng/ml se han detectado en ovejas sin actividad ovulatona (anestro estacional), en las cuales no hay cuerpos lúteos; estas observaciones generan la necesidad de revisar en forma detallada la metodología desanollada y los criterios específicos utilizados para afirmar que una concentración de progesterona sérica o plasmática de 3.92 ng/ml conesponde a hembras ovinas en anestro. La "oveja Chiapas" puede ser un recurso genético importante en México; sin embargo, es necesario desarrollar estudios con metodologías que permitan determinar aspectos fisiológicos y productivos de la raza.
Por otro lado, Anoyo et al. (2007), en el altiplano central de la República Mexicana (19° Lat. N), determinaron la actividad ovulatoria anual en ovejas Suffolk estabuladas y con alimentación controlada, a través de colecciones sanguíneas realizadas dos veces por semana durante dos años, con la posterior determinación de la concentración plasmática de progesterona. Las ovejas Suffolk expresaron anestro estacional claro y definido (Figura 4), similar al observado en ovejas de la misma raza que habitan en latitudes altas (≥35° Lat. N; Legan y Karsch, 1979; Karsch et al., 1984); lo anterior, a pesar de las ligeras variaciones en la amplitud del fotoperiodo características de esta latitud (2h, 17 min, entre el día más largo y el más corto del año). La temperatura ambiental no se asoció con el anestro estacional o la actividad ovulatoria anual de las ovejas (Arroyo et al., 2007). Es importante mencionar que las observaciones se realizaron por dos años consecutivos, lo cual permitió determinar la repetibilidad del comportamiento reproductiva anual en esta raza.
La información analizada indica dos tendencias claras con respecto a las ovejas de lana en México. Primero, se observa una época reproductiva anual extensa en las ovejas criollas, con tendencia a ciclar de manera continua. Aproximadamente el 80% de las hembras presentan estros durante la época de días largos (anestro estacional). Con base en lo anterior, se sugiere que por su origen, no son sensibles a los cambios anuales en la duración del fotoperiodo y este factor ambiental, posiblemente no regula su ciclo reproductivo anual. Por lo tanto, el mecanismo neuroendocrino que en las razas europeas induce el anestro estacional, no es funcional en las ovejas criollas de lana. Sin embargo, aun no se ha investigado el origen fisiológico de este fenómeno. La segunda conclusión relevante se refiere a la clara estacionalidad reproductiva observada en ovejas de razas europeas que habitan en la República Mexicana; esto ocurre a pesar de que las hembras se encuentran en una región cercana al ecuador. De acuerdo con estas observaciones se puede inferir que en estos animales, el mecanismo neuroendocrino que regula el inicio y término del anestro estacional, es funcional y son sensibles a las ligeras variaciones anuales en la duración del fotoperiodo observadas en México (2 h, 17 min, entre el día más largo y el más corto del año).
Es necesario señalar que la mayoría de los estudios referentes a la estacionalidad reproductiva en ovinos de lana en México, se desarrollaron en la región central del país (19° Lat. N). Entonces, resulta de interés realizar estudios similares, en latitudes distintas, con el propósito de establecer las posibles variaciones en el comportamiento reproductivo anual de estas razas.
ESTACIONALIDAD REPRODUCTIVA EN OVEJAS PELIBUEY
Actividad estral
En la década de los 50, Hafez (1952) mencionó que las ovejas de origen ecuatorial presentan una reducida estacionalidad reproductiva e incluso, ausencia de la misma, siendo capaces de reproducirse todo el año. Esta afirmación se aplicó a los primeros rebaños de ovejas Pelibuey en México (Castillo et al., 1972; Castillo et al., 1974). Sin embargo, en estudios más recientes, se reportó disminución de la actividad estral en los meses de febrero a mayo, atribuida a deficiencias nutricionales y factores ambientales como temperatura y humedad (Heredia et al., 1991; González-Reyna et al., 1991; González-Reyna et al., 1992). Por lo tanto, la variación entre resultados generó controversia.
En Yucatán (21° Lat. N), México, Valencia et al. (1981), estudiaron la actividad estral durante tres años en ovejas Pelibuey con alimentación controlada, y observaron que de enero a abril, 17% de las ovejas mostraron estro; en contraste, 95 % de las ovejas presentaron actividad estral de mayo a agosto y 100 % de septiembre a diciembre. Se concluye que las ovejas Pelibuey, en esas condiciones ambientales, presentaron anestro estacional, independientemente del estado nutricional.
Por su parte, Heredia et al. (1991) evaluaron la actividad estral en ovejas Pelibuey con alimentación controlada durante tres años y observaron que de enero a mayo, sólo 15 % de las ovejas mostraron estro; en contraste, 90% de las hembras estuvieron en celo entre los meses de agosto a diciembre. Esta reducción en la actividad estral durante la primavera, ocurrió independientemente del estado nutricional, la condición o el peso corporal de las ovejas.
González-Reyna et al. (1992), utilizaron dos machos adultos vasectomizados para estudiar, durante un año, la actividad estral en ovejas Pelibuey estabuladas, con alimentación controlada y constante en Tamaulipas, México (22° Lat. N), con un clima tropical semiárido. Los meses con el menor porcentaje de actividad estral fueron marzo (34 %) y abril (24 %); el mayor porcentaje de actividad estral fue en agosto (97 %) y septiembre (88 %). No encontraron diferencias (P>0.05) en la proporción de estros en julio, agosto, septiembre y diciembre. Se observaron diferencias (P<0.05) en agosto y septiembre al compararlos con marzo y abril, pero no hubo diferencias (P>0.05) con respecto a los meses restantes. Se concluyó que la variación anual observada en la actividad estral de las ovejas Pelibuey no implica la existencia de anestro estacional y que este comportamiento puede tener su origen en la variación de factores ambientales, como la temperatura y humedad. Sin embargo, estas conclusiones posiblemente no tengan un fundamento sólido, pues la reducción en la actividad estral en los meses de marzo y abril, indica un elevado porcentaje de ovejas no cíclicas, lo que podría indicar un periodo de anestro corto, de aproximadamente 2 meses.
Cruz et al. (1994), en Veracruz (20° Lat. N), México (clima tropical subhúmedo), determinaron la actividad estral en ovejas Pelibuey, utilizando machos vasectomizados. Las ovejas estuvieron en pastoreo y suplementadas con sales minerales ad libitum. La menor proporción de hembras en estro se observó en abril (81.25 %) y la mayor en agosto (100 %); sin embargo, estas variaciones no fueron diferentes (P>0.05). Los autores concluyeron que bajo las condiciones del estudio, no hubo diferencia en la presentación de estros a través del año en la oveja Pelibuey y que esta raza es capaz de mostrar actividad estral anual continua.
Valencia et al. (2006), evaluaron la actividad estral en ovejas Pelibuey (19° Lat. N) durante 8 meses (diciembre a julio). Se utilizaron ovejas adultas y nulíparas; la actividad estral en ambos grupos se monitoreó utilizando machos adultos provistos con mandil, los cuales se introdujeron diariamente a los corrales de las hembras por un periodo de 15 minutos. Se emplearon cinco machos Pelibuey y el semental se cambió cada dos semanas. Los resultados mostraron que entre el 84.6% y 100% de las ovejas adultas mostraron actividad estral regular entre enero y mayo; en las nulíparas, la actividad estral fue menor en todos los meses. Hubo diferencias entre grupos (P<0.05) de enero a abril (Cuadro 2). Diez ovejas adultas (38%) y cuatro nulíparas (19%) presentaron ciclos estrales sin interrupción durante el periodo de observación. La duración del anestro fue diferente (P<0.05) entre adultas (36.1±8.1 días) y primalas (52.8±23.4 días).
En los estudios anteriores se observan resultados variables. Es evidente la reducción en la actividad estral en la oveja Pelibuey entre enero y junio (época de anestro estacional en regiones septentrionales); sin embargo, en todos los reportes existe una proporción variable (17-96%) de ovejas que continúan ciclando durante esos meses, independientemente de las variaciones ambientales y de alimentación.
Actividad Ovulatoria
Por otro lado, se evaluó la actividad ovulatoria anual en la oveja pelibuey, utilizando diferentes modelos experimentales. Los estudios fueron realizados con el propósito de establecer la duración y expresión del anestro estacional en esta raza y determinar los posibles factores que pudieran influir en su presentación. En primer lugar, Martínez et al. (1995) determinaron la actividad ovulatoria, los cambios de peso y condición corporal durante un año en ovejas Pelibuey en pastoreo, bajo clima tropical subhúmedo (20° Lat. N). La actividad ovulatoria se determinó por medio de la concentración plasmática de progesterona (radioinmunoanálisis en fase sólida) a partir de muestras sanguíneas obtenidas dos veces por semana. Los resultados mostraron reducción (P<0.05) en la actividad ovulatoria de las ovejas en abril y mayo (75 y 50 %, respectivamente), a pesar de que durante esa época, los pesos y condición corporal fueron los más altos. Se concluyó que la raza Pelibuey disminuye su actividad ovulatoria durante la primavera, independientemente de las variaciones anuales en peso y condición corporal. El anestro estacional no se correlacionó con deficiencias nutricionales; por lo tanto, se sugiere que la actividad ovulatoria anual puede estar regulada por las variaciones anuales en la duración del fotoperiodo. En el estudio no se realizó detección de estros; sin embargo, las ovejas permanecieron constantemente con carneros vasectomizados.
En el trabajo de Martínez et al. (1995), es clara la disminución en la actividad ovulatoria de las hembras en abril y mayo, pero de manera similar a los estudios de Valencia et al. (1981), Heredia et al. (1991), González-Reyna et al. (1992) y Cruz et al. (1994), algunas ovejas Pelibuey muestran actividad reproductiva durante la época de días largos. Esta observación ha sido constante en los estudios revisados.
Por su parte, Porras (1999), determinó la actividad ovulatoria en ovejas Pelibuey en condiciones de fotoperiodo natural (19° Lat. N) y alimentación controlada, por dos años. Observó que las hembras tuvieron un periodo de anestro, de duración variable, en invierno y primavera. Durante el primer año, el anestro ocurrió entre enero y mayo (duración de 63.7±18.8 días); y entre febrero y julio (duración 109±20.5 días) en el segundo año. Dos ovejas presentaron actividad ovulatoria continua. En este estudio se registró por primera vez la actividad ovulatoria individual de las ovejas y fue el punto de partida para iniciar estudios similares e identificar ovejas con esta característica fisiológica.
Cerna et al. (2000), en un estudio desarrollado en condiciones ambientales similares al de Porras (1999; 19° Lat. N) y sin ser su objetivo principal, determinó durante dos años, la actividad ovulatoria en ovejas Pelibuey y encontró periodos de anestro también variables, que se extienden de enero a julio, en ovejas mantenidas en fotoperiodo natural (Figura 5).
Los objetivos principales de los estudios de Porras (1999) y Cerna et al. (2000) fueron evaluar la actividad ovulatoria de la oveja Pelibuey al aplicar fotoperiodo artificial, característico de latitudes altas (>35°) en el primer caso y fotoperiodo inverso característico de una latitud de 19° N en el segundo caso. Ambos autores demostraron que las ovejas Pelibuey responden a los cambios artificiales en la duración del fotoperiodo, lo cual indica que este factor ambiental puede regular el ciclo reproductivo anual de la raza. Otro resultado relevante fue que en esas condiciones de fotoperiodo artificial, dos (Porras, 1999) y tres (Cerna et al., 2000) ovejas ovularon continuamente sin importar la duración de horas luz aplicada.
Valencia et al. (2006) determinaron la actividad ovulatoria de ovejas Pelibuey adultas y nulíparas durante un periodo de 8 meses (diciembre a julio). Los resultados mostraron una proporción cercana al 100% de ovejas adultas ovulando en los meses de diciembre y enero, con un descenso en marzo, abril y mayo (52, 37 y 33 %, respectivamente). El 29.6% de las hembras ovularon sin interrupción durante el periodo de observación. La duración promedio del anestro fue de 39.7±31.7 (media±D.E.) días. Una observación interesante es que las ovejas nulíparas mostraron actividad ovulatoria menor (P<0.05) en los meses de abril a julio (5.6, 5.6, 5.9 y 29.4 %, respectivamente) que las hembras adultas; es importante resaltar que ninguna oveja nulípara ovuló continuamente.
Con los estudios anteriores se identificaron algunas ovejas Pelibuey que ovulan durante la época de días largos, sin embargo, esta observación se realizó por periodos relativamente cortos y no se registraron los eventos en el resto del año. Tampoco se evaluó si el comportamiento ovulatono de estas ovejas puede ser repetible entre años. Posteriormente, Arroyo et al. (2007), caracterizaron la actividad ovulatoria anual en ovejas Pelibuey a 19° Lat. N, en ausencia de machos y con alimentación controlada, a través de la concentración plasmática de progesterona, obtenida de muestras sanguíneas colectadas dos veces por semana durante dos años. Los pesos de las ovejas y la temperatura ambiental se registraron durante todo el estudio. Los resultados mostraron que el 60 % de las ovejas ovularon continuamente durante los dos años de observación, independientemente de la amplitud del fotoperiodo y las variaciones en la temperatura ambiental (Figura 6). El peso corporal de las ovejas fue constante (P>0.05) durante todo el estudio. Una oveja ovuló sin interrupción durante un año y en el segundo se eliminó del estudio al desarrollar un quiste folicular. Tres ovejas mostraron periodos similares (P>0.05) de anestro con una duración de 29 a 132 días (65±46.6; Media ± DE) de julio a diciembre en el primer año y de 29 a 92 días entre enero y abril en el segundo año (70.6±36; Media ± DE). Esta variación en la duración del anestro en la oveja Pelibuey también la observó Porras (1999); sin embargo, en el estudio de Arroyo et al. (2007), ocurrió un desplazamiento del anestro entre años, en el primer año, sucedió entre julio y diciembre; después del solsticio de verano, periodo en el cual las otras hembras se encontraban en plena época reproductiva. Esto podría indicar que no existe una sincronía entre la interrupción de la actividad ovulatoria de estas ovejas y la señal clásica del fotoperiodo que pone a tiempo el inicio y termino del anestro estacional (Karsch et al., 1984). De manera contraria, en el segundo año, las tres ovejas mostraron anestro entre enero y abril, cuando las horas luz se incrementaron. No se tiene una explicación de esta variación entre años, la temperatura ambiental y los cambios en el peso corporal no se correlacionaron con este comportamiento. Sin embargo, se sugiere que el ritmo reproductivo endógeno de las ovejas que mostraron anestro durante los días cortos, moduló el inicio del anestro, sin que el fotoperiodo regulara este evento.
En el estudio de Arroyo et al. (2007), se demostró que a 19° Lat. N, en un clima semifrío subhúmedo, existen ovejas Pelibuey con la capacidad de ovular todo el año, independientemente de las variaciones en la amplitud del fotoperiodo. La explicación fisiológica de esta característica en dicha raza se desconoce. Dos hipótesis se plantearon para explicar este comportamiento en ovinos originarios de latitudes ecuatoriales, la primera indica que los animales son insensibles al fotoperiodo y la segunda propone que las variaciones en la amplitud del fotoperiodo natural son tan ligeras que no son detectadas por los animales (Chemineau et al. 2004). Las observaciones de Arroyo et al. (2007) prueban la primera hipótesis, pues con los cambios ligeros en la duración del fotoperiodo a 19° Lat. N (2 h, 17 min entre el día más largo y el más corto del año), el 40% de las ovejas Pelibuey interpretaron la señal luminosa y presentaron anestro estacional. Por lo tanto, el estímulo luminoso en algunas ovejas puede inducir un cambio en la sensibilidad del hipotálamo a la concentración circulante de estradiol, generando un efecto de retroalimentación negativa a nivel central, lo cual activa las neuronas dopaminérgicas en el núcleo Al5 del área retroquiasmática lateral y suprime la secreción pulsátil de GnRH/LH, mecanismo neuroendocrino clásico que provoca el anestro estacional y se ha descrito en ovejas originarias de latitudes altas (>35°) (Thiéry et al., 1995; Gallegos-Sánchez et al., 1997; Lehman et al., 2002; Thiéry et al., 2002).
Recientemente se propuso (Arroyo et ai, 2007) que en la oveja Pelibuey existen tres modelos de respuesta al fotoperiodo: en el primero, se incluyen aproximadamente el 40% de las hembras y se plantea que las ovejas son sensibles a los cambios ligeros en el fotoperiodo, característicos de regiones ecuatoriales y muestran anestro estacional; un segundo grupo, que en los rebaños podría alcanzar el 60% de las hembras, son insensibles a las variaciones en la amplitud del fotoperiodo ecuatorial y ovulan todo el año; y un tercer grupo, con un elevado umbral de respuesta a las variaciones en el fotoperiodo, que ovula aun al ser expuesto a fotoperiodos característicos de latitudes altas (>35°).
CONCLUSIONES
El fotoperiodo es el factor ambiental primario que regula el ciclo reproductivo anual de la oveja. Las variaciones anuales en la síntesis y secreción de melatonina, en conjunto con neurotransmisores, tales como, Kisspeptina, Dopamina, GABA e indicadores metabólicos como Leptina, pueden modular el inicio o termino de la época reproductiva. En México, las ovejas criollas de lana y las ovejas Pelibuey, muestran un corto anestro estacional ó actividad reproductiva anual continua. Cuando se presenta el anestro estacional, este ocurre entre marzo y junio. La oveja Pelibuey responde a los cambios anuales en la duración del fotoperiodo; sin embargo, la respuesta a este estimulo es variable. Se han identificado ovejas que responden a los cambios en la amplitud del fotoperiodo ecuatorial, mostrando anestro estacional; otras ovejas, que no responden a las variaciones en las horas luz, ovulan todo el año y un último grupo de ovejas, extremadamente insensibles al fotoperiodo, mantienen su actividad reproductiva a pesar de ser expuestas a fotoperiodos propios de latitudes altas (>35°). Por otro lado, las ovejas de razas europeas lanares nacidas en México, expresan estacionalidad reproductiva similar a la de individuos de la misma raza que habitan o nacen en latitudes altas. Lo cual indica que los mecanismos neuroendocrinos clásicos que regulan el ciclo reproductivo anual en estas razas, se encuentran activos a pesar de encontrarse en una región cercana al ecuador; situación que no ocurre con las ovejas criollas y de pelo nacidas en la república mexicana, en las cuales, se sugiere que este mecanismo no es funcional.
REFERENCIAS
Álvarez, R.L., Zarco, L. 2001. Los fenómenos de bioestimulación sexual en ovejas y cabras. Veterinaria México. 32: 117-129. [ Links ]
Arendt, J. 1998. Melatonin and the pineal gland: influence on mammalian seasonal and circadian physiology. Reviews of Reproduction. 3: 13-22. [ Links ]
Arroyo, J., Magaña-Sevilla, H., Camacho-Escobar, M.A. 2009. Regulación neuroendocrina del anestro posparto en la oveja. Tropical and Subtropical Agroecosystems. 10: 301-312. [ Links ]
Arroyo, L.J., Gallegos-Sánchez, J., Villa-Godoy, A., Berruecos, J.M., Perera, G., Valencia, J. 2007. Reproductive activity of Pelibuey and Suffolk ewes at 19° north latitude. Animal Reproduction Science. 102: 24-30. [ Links ]
Backholer, K., Smith, J.T., Rao, A., Pereira, A., Iqbal, J., Ogawa, S., Li, Q., Clarke, L.J. 2010. Kisspeptin cells in the ewe brain respond to leptin and communicate with neuropeptide Y and proopiomelanocortin cells. Endocrinology. 151: 2233-2243. [ Links ]
Barrell, G.K., Moenter, M.S., Caraty, A., Karsch, J.F. 1992. Seasonal changes of gonadotropin -releasing hormone secretion in the ewe. Biology of Reproduction. 46: 1130-1135. [ Links ]
Barrell, G.K., Thrun, L.A., Brown, M.E., Viguié, C, Karsch, F.J. 2000. Importance of photoperiodic signal quality to entrainment of the circannual reproductive rhythm of the ewe. Biology of Reproduction. 63: 769-774. [ Links ]
Bartlewski, M.P., Vanderpol, J., Beard, P.A., Cook, J.S., Rawlings, C.N. 2000. Ovarian antral follicular dynamics and their associations with peripheral concentrations of gonadotropins and ovarian steroids in anoestrous Finnish Landrace ewes. Animal Reproduction Sciencie. 58: 273-291. [ Links ]
Bentley, G.E., Perfito, N., Ukena, K., Tsutsui, K., Winfield, J.C. 2003. Gonadotropin-inhibitory peptide in song sparrows (Melospiza melodid) in different reproductive conditions, and in house sparrows {Passer domesticus) relative to chicken-gonadotropin-releasing hormone. JournalNeuroendocrinology. 15: 794-802. [ Links ]
Bittman, E.L., Karsch, F.J. 1984. Nightly duration of pineal melatonin secretion determines the reproductive response to inhibitory day length in the ewe. Biology of Reproduction. 30: 585-593. [ Links ]
Blache, D., Tellam, R.L., Chagas, L.M., Blackberry, M.A., Vercoe, P.E., Martin, G.B. 2000. Level of nutrition affects leptin concentrations in plasma and cerebrospinal fluid in sheep. Journal of Endocrinology. 165: 625-637. [ Links ]
Bogusz, A.L., Hardy, L.S., Lehman, M.N., Connors, J.M., Hileman, S.M., Sliwowska, H., Billings, H.J., McManus, C.J., Valent, M., Singh, S.R, Nestor, C.C., Coolen, L.M., Goodman, R.L. 2008. Evidence that gamma-aminobutyric acid is part of the neural circuit mediating estradiol negative feedback in anestrous ewes. Endocrinology. 149: 2762-2772. [ Links ]
Brown, R.E., Imran, S.A., Ur, E., Wilkinson, M. 2008. Kiss-lmRNA in adipose tissue is regulated by sex hormones and food intake. Molecular and Cellular Endocrinology. 281: 64-72. [ Links ]
Castillo, R.H., Valencia, Z.M., Berruecos, J.M. 1972. Comportamiento reproductivo del borrego Tabasco mantenido en clima tropical y subtropical. I. índices de fertilidad. Técnica Pecuaria México. 20: 52-56. [ Links ]
Castillo, R.H., Román, P.H., Berruecos, J.M. 1974. Características del crecimiento del borrego Tabasco. I. Efecto de la edad y peso al destete y su influencia sobre la fertilidad de la madre. Técnica Pecuaria México. 27: 28-32. [ Links ]
Cerna, C, Porras, A., Valencia, M.J., Perera, G., Zarco, L. 2000. Effect of an inverse subtropical (19°13' N) photoperiod on ovarian activity, melatonin and prolactin secretion in Pelibuey ewes. Animal Reproduction Science. 60-61: 511-525. [ Links ]
Chemineau, P., Daveau, A., Cognié, Y., Aumont, G., Chesneau, D. 2004. Seasonal ovulatory activity exists in tropical Creole female goats and Black Belly ewes subjected to a temperate photoperiod. BMC Phisiology. 4:12. [ Links ]
Clarke, I.J., Backholer, K., Tilbrook, A.J. 2005. Y2 receptor-selective agonist delays the estrogen-induced luteinizing hormone surge in ovariectomized ewes, but yl-receptor-selectiven agonist stimulates voluntary food intake. Endocrinology. 146: 769-775. [ Links ]
Clarkson, J., d'Anglemont de Tassigny, X., Santos-Moreno, A., Colledge, W.H., Herbison, A.E. 2008. Kisspeptin-GPR54 singaling is essential for preovulatory gonadotropin-releasing hormone neuron activation and the luteinizing hormone surge. The Journal of Neuroscience. 28: 8691-8697. [ Links ]
Cruz, L.C., Fernández-Baca, S., Alvarez, L.J., Pérez, R.H. 1994. Variaciones estacionales en la presentación de la ovulación, fertilización y sobrevivencia embrionaria de ovejas Tabasco en el trópico húmedo. Veterinaria México. 25: 23-27. [ Links ]
De Lucas-Tron, J., González-Padilla, E., Martínez, R.L. 1997. Estacionalidad reproductiva en ovejas de cinco razas en el altiplano central mexicano. Técnica Pecuaria México. 35: 25-31. [ Links ]
De Lucas-Tron, J. 2000. Situación de la producción ovina en México y perspectivas para el nuevo siglo. In: Curso Avances en nutrición ovina I, Especialidad en Producción Ovina, Facultad de Medicina Veterinaria Y Zootecnia, Universidad Autónoma del Estado de México, El Cerrillo Piedras Blancas, Toluca, Edo. de México, pp. 8-20. [ Links ]
Delgadillo, J.A., Vielma, J., Flores, J.A., Veliz, F.G., Duarte, G., Hernández, H. 2008. La calidad del estímulo emitido por el macho determina la respuesta de las cabras sometidas al efecto macho. Tropical and Subtropical Agroecosystems. 9: 39-45. [ Links ]
Dyer, C.J., Simmons, J.M., Matteri, R.L., Keisler, D.H. 1997. cDNA cloning and tissue-specific gene expression of ovine leptin, NPY-Y1 receptor, and NPY-Y2 receptor. Domestic Animal Endocrinology. 14: 295-303. [ Links ]
Estrada, K.M., Clay, CM., Pompólo, S., Smith, J.T., Clarke, I.J. 2006. Elevated Kiss-1 expression in the arcuate nucleus prior to the cyclic preovulatory gonadotropin-releasing hormone/luteinizing hormone surge in the ewe suggests a stimulatory role for kisspeptin in oestrogen-positive feedback. Journal Neuroendocrinology. 18: 806-809. [ Links ]
Evans, A.C.O., Duffy, P., Hynes, N., Boland, M.P. 2000. Waves of follicle development during the estrous cycle in sheep. Theriogenology. 53: 699-715. [ Links ]
Evans, N.P., Richter, T.A., Skinner, D.C., Robinson, J.E. 2002. Neuroendocrine mechanisms underlying the effects of progesterone on the oestradiol-induced GnRH/LH surge. Reproduction Supplement. 59: 57-66. [ Links ]
Flores, J.A., Veliz, F.G., Pérez-Villanueva, J.A., Martínez de la Escalera, G., Chemineau, P., Poindron, P., Malpaux, B., Delgadillo, J.A. 2000. Male reproductive condition is the limiting factor of efficiency in the male effect during seasonal anestrus in female goats. Biology of Reproduction. 62: 1409-1414. [ Links ]
Gallegos-Sánchez, J., Delaleu, B., Caraty, A., Malpaux, B., Thiéry, J.C. 1997 Estradiol acts locally within the retrochiasmatic area to inhibit pulsatile luteinizing-hormone release in the female sheep during anestrus. Biology of Reproduction. 56: 1544-1549. [ Links ]
Gayrard, V., Malpaux, B., Tillet, Y. Thiéry, J.C. 1994. Estradiol increases tyrosine hydroxylase activity of the A15 nucleus dopaminergic neurons during long days in the ewe. Biology of Reproduction. 50: 1168-1177. [ Links ]
Goodman, R.L., Gibson, M., Skinner, D.C., Lehman, M.N. 2002. Neuroendocrine control of pulsatile GnRH secretion during the ovarian cycle: evidence from ewe. Reproduction Supplement. 59:41-56. [ Links ]
González-Reyna, A., Valencia, J., Foot, W.C., Murphy, B.D. 1991. Hair sheep in México: Reproduction in the Pelibuey sheep. Animal Breeding Abstracts. 59: 509-524. [ Links ]
González-Reyna , A., Murphy, B.D., Foot, W.C. 1992. Circannual estrous variations and ovulation rate in Pelibuey ewes. Small Ruminant Research. 8: 225-232. [ Links ]
Goubillon, M.L., Delaleu, B., Tillet, Y., Caraty, A., Herbison, A.E. 1999. Localization of estrogen-receptive neurons projecting to the GnRH neuron-containing rostral preoptic area of the ewe. Neuroendocrinology. 70: 228-236. [ Links ]
Greives, T.J., Mason, A.O., Scotti, M.A.L., Levine, J., Ketterson, E.D., Kriegsfeld, L.J., Demás, G.E. 2007. Enviromental control of kisspeptin: implications for seasonal reproduction. Endocrinology. 148: 1158-1166. [ Links ]
Greives, T.J., Kriegsfeld, L.J., Bentley, G.E., Tsutsui, K., Demás, G.E. 2008. Recent advances in reproductive neuroendocrinology: a role for RFamide peptides in seasonal reproduction? Proceedings of the Royal Society B. 275: 1943-1951. [ Links ]
Hafez, E.S.E. 1952. Studies on the breeding season and reproduction of the ewe. Journal Agricultural Science. 42: 189-265. [ Links ]
Hameed, S., Jayasena, C.N., Dhillo, W.S. 2011. Kisspeptin and fertility. Journal of Endocrinology. 208: 97-105. [ Links ]
Havern, L.R., Whisnant, S.C., Goodman, L.R. 1994. Dopaminergic structures in the ovine hypothalamus mediating estradiol negative feedback in anestrous ewes. Endocrinology. 134: 1905-1914. [ Links ]
Heredia, A., Menéndez, T.M., Velázquez, M.A. 1991. Factores que influyen en la estacionalidad reproductiva de la oveja Pelibuey. Memorias de la Reunión Nacional de Investigación Pecuaria. Cd. Victoria, Tamaulipas, México, p. 115. [ Links ]
Himuna, S., Shintani, Y., Fukusumi, S., Iijima, N, Matsumoto, Y., Hosoya, M., Fujii, R., Watanabe, T., Kikuchi, K., Terao, Y., Yano, T., Yamamoto, T., Kawamata, Y., Habata, Y., Asada, M., Kitada, C, Kurokawa, T., Onda, H., Nishimura, O., Tanaka, M., Ibata, Y., Fujino, M. 2000. New neuropeptides containing carboxyl-terminal RF-amide and their receptor in mammals. Nature Cell Biology. 2: 703-708. [ Links ]
Hulet, C.V. 1966. Behavioral social and psychological factors affecting mating time and breeding efficiency in sheep. Journal of Animal Science. 25:5-16. [ Links ]
Jackson, G.L., Kuehl, D. 2002. Gamma-aminobutyric acid (GABA) regulation of GnRH secretion in sheep. Reproduction, Supplement. 59: 15-24. [ Links ]
Karsch, F.J., Bittman, L.E., Foster, L.D., Goodman, L.R., Legan, J.S., Robinson, E.J. 1984. Neuroendocrine basis of seasonal reproduction. Recent Progress in Hormone Research. 40: 185-231. [ Links ]
Keisler, D.H., Lucy, M.C. 1996. Perception and interpretation of the effects of undernutrition on reproduction. Journal of Animal Science. 74 (Suppl. 3): 1-17. [ Links ]
Knight, T.W., Lynch, P.R. 1980. Source of ram pheromones that stimulate ovulation in the ewe. Animal Reproduction Science. 3: 133-136. [ Links ]
Legan, J.S., Karsch, J.F. 1979. Neuroendocrine regulation of the estrous cycle and seasonal breeding in the ewe. Biology of Reproduction, 20: 74-85. [ Links ]
Lehman, M.N., Karsch, F.J. 1993. Do gonadotropin-releasing hormone, tyrosine hydroxylase-, and J -endorphin-immunoreactive neurons contain estrogen receptors? A double-label immunocytochemical study in the Suffolk ewe. Endocrinology. 133: 887-895. [ Links ]
Lehman, M.N., Coolen, L.M., Goodman, R.L., Viguié, C, Billings, H.J., Karsch, F.J. 2002. Seasonal plasticity in the brain: the use of large animal models for neuroanatomical research. Reproduction Supplement. 59: 149-165. [ Links ]
Lincoln, G.A., Short, R.V. 1980. Seasonal breeding: Nature's contraceptive. Recent Progress in Hormone Research. 36: 1-52. [ Links ]
Malpaux, B., Robinson, J.E., Brown, M.B., Karsch, F.J. 1987. Reproductive refractoriness of the ewe to inductive photoperiod is not caused by inappropriate secretion of melatonin. Biology of Reproduction. 36: 1333-1341. [ Links ]
Malpaux, B., Viguié, C, Skinner, D.C., Thiéry, J.C., Pelletier, J., Chemineau, P. 1996. Seasonal breeding in sheep: Mechanism of action of melatonin. Animal Reproduction Science, 42: 109-117. [ Links ]
Malpaux, B., Viguié, C, Skinner, D.C., Thiéry, J.C., Chemineau, P. 1997. Control of the circannual rhythm of reproduction by melatonin in the ewe. Brain Research Bulletin. 44: 431-438. [ Links ]
Malpaux, B., Thiéry, J.C., Chemineau, P. 1999. Melatonin and the seasonal control of reproduction. Reproduction Nutrition Development. 39: 355-366. [ Links ]
Malpaux, B., Tricoire, H., Mailliet, F., Daveau, A., Migaud, M., Skinner, D.C., Pelletier, J., Chemineau, P. 2002. Melatonin and seasonal reproduction: understanding the neuroendocrine mechanisms using the sheep as a model. Reproduction Supplement. 59: 167-179. [ Links ]
Martínez, R.R.D., Zarco, Q.L., Cruz, L.C., Rubio, G.I. 1995. La estacionalidad de la actividad ovárica en la oveja Pelibuey es independiente de variaciones en el peso o condición corporal de los animales. Memorias del VIII Congreso Nacional de Producción Ovina, Asociación Mexicana de Técnicos Especialistas en Ovinocultura, A.C. Chapingo, México, p. 131-134. [ Links ]
McMillen, J.C., Houghton, D.C., Young, I.R. 1995. Melatonin and the development of circadian and seasonal rhythmicity. Journal of Reproduction and Fertility, Suppl. 49: 137-146. [ Links ]
McNatty, K.P., Hudson, N.L., Henderson, K.M., Lun, S., Heath, D.A., Gibb, M., Ball, K., McDiarmid, J.M., Thurley, D.C. 1984. Changes in gonadotropin secretion and ovarian antral follicular activity in seasonally breeding sheep throughout the year. Journal of Reproduction and Fertility. 70:309-321. [ Links ]
Meza-Herrera, C.A., Veliz-Deras, F.G., Wurzinger, M., López-Ariza, B., Arellano-Rodriguez G., Rodríguez-Martínez, R. 2010. The kiss-1-kisspeptin-gpr54 complex: a critical modulator of GnRH neurons during pubertal activation. Journal Applied Biomedicine. 8: 1-9. [ Links ]
Noel, B., Bister, J.L., Paquay, R. 1993. Ovarian follicular dynamics in Suffolk ewes at different periods of the year. Journal of Reproduction and Fertility. 99: 695-700. [ Links ]
Oakley, A.E., Clifton, D.K., Steiner, R.A. 2009. Kisspeptin signaling in the brain. Endocrine Reviews. 30: 713-743. [ Links ]
O'Callaghan, D., Donovan, A., Sunderland, S.J., Boland, M.P., Roche, J.F. 1994. Effect of the presence of male and female flockmates on reproductive activity in ewes. Journal of Reproduction and Fertility. 100: 497-503. [ Links ]
Okamura, H., Mori, Y. 2005. Characterization of the primer pheromone molecules responsible for the "male effect" in ruminant species. Chemical Senses. 30 (Suppl. 1): i140-i141. [ Links ]
Padilla, R.F.J., Mapes, S.G.E., Jimenez, K.F. 1988. Perfiles hormonales durante el ciclo estral de la oveja. Técnica Pecuaria México. 28: 96-108. [ Links ]
Pedraza, P., Peralta, M., Pérez-Grovas, R. 1992. El borrego Chiapas: una raza local mexicana de origen español. Archivos de Zootecnia. 41: 355-362. [ Links ]
Perezgrove, G.R., Castro, G.H. 2000. El borrego Chiapas y el sistema tradicional de manejo de ovinos entre las pastoras Tzotziles. Archivos de Zootecnia. 49: 391-403. [ Links ]
Pompolo, S., Pereira, A., Estrada, K.M., Clarke, I.J. 2006. Colocalization of kisspeptin and gonadotropin-releasing hormone in the ovine brain. Endocrinology. 147: 804-810. [ Links ]
Porras, A.A.I. 1999. Efectos del fotoperiodo artificial sobre la actividad reproductiva de la oveja Pelibuey. Tesis de Doctorado en Ciencias Veterinarias. Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México. México, D.F. [ Links ]
Ravindra J.P., Rawlings N.C. 1997. Ovarian follicular dynamics in ewes during the transition from anoestrus to the breeding season. Journal of Reproduction and Fertility. 110: 279-289. [ Links ]
Rawlings, N.C., Kennedy, S.W., Chang, C.H., Hill, J.R., Henricks, D.M. 1977. Onset of seasonal anestrus in the ewe. Journal of Animal Science. 44: 791-797. [ Links ]
Robinson, J.E., Karsch, F.J., 1984. Refractoriness to inductive day lengths terminates the breeding season of the Suffolk ewe. Biology of Reproduction. 31: 656-663. [ Links ]
Robinson, J.E., Wayne, N.L., Karsch, F.J. 1985. Refractoriness to inhibitory day lengths initiates the breeding season of the Suffolk ewe. Biology of Reproduction. 32: 1024-1030. [ Links ]
Robinson J.E., Kendrick K.M. 1992. Inhibition of luteinizing hormone secretion in the ewe by progesterone: associated changes in the release of gamma-aminobutyric acid and noradrenaline in the preoptic area as measured by intracranial microdialysis. Journal Neuroendocrinology. 4: 231-236. [ Links ]
Robinson, J.E. 1995. Gamma amino-butyric acid and the control of GnRH secretion in sheep. Journal of Reproduction and Fertility. Supplement 49: 221-230. [ Links ]
Rosa, H.J.D., Bryant, M.J. 2003. Seasonality of reproduction in sheep. Small Ruminant Research. 48: 155-171. [ Links ]
Schillo, K.K. 1992. Effects of dietary energy of control of luteinizing hormone secretion in cattle and sheep. Journal of Animal Science. 70: 1271-1282. [ Links ]
Smith, J.T., Clifton, D.K., Steiner, R.A., 2006. Regulation of the neuroendocrine reproductive axis by kisspeptin-GPR54 signaling. Reproduction. 131: 623-630. [ Links ]
Smith, J.T., Clay, CM., Caraty, A., Clarke, I.J. 2007. Kiss-1 messenger ribonucleic acid expression in the hypothalamus of the ewe is regulated by sex steroids and season. Endocrinology. 148: 1150-1157. [ Links ]
Smith, J.T., Coolen, L.M., Kriegsfeld, L.J., Sari, IP., Jaafarzadehshirazi, M.R., Maltby, M., Bateman, K., Goodman, R.L., Tilbrook, A.J., Ubuka, T., Bentley, G.E., Clarke, I.J., Lehman, M.N. 2008. Variation in kisspeptin and RFamide-related peptide (RFRP) expression and terminal connections to gonadotropin-releasing hormone neurons in the brain: a novel medium for seasonal breeding in the sheep. Endocrinology. 149: 5770-5782. [ Links ]
Snyder, J.L., Clapper, J.A., Roberts, A.J., Sansón, D.W., Hamernik, D.L., Moss, G.E. 1999. Insuline-like growth factor-I, insulin-like growth factor-binding proteins, and gonadotropins in the hypothalamic-pituitary axis and serum of nutrient-restricted ewes. Biology of Reproduction. 61: 219-224. [ Links ]
Souza, J.H.C., Campbell, K.B., Baird, T.D. 1997. Follicular dynamics and ovarian steroid secretion in sheep during the follicular and early luteal phases of the estrous cycle. Biology of Reproduction. 56: 483-488. [ Links ]
Tatman, W.R., Judkins, M.B., Dunn, T.G., Moss, G.E. 1990. Luteinizing hormone in nutrient-restricted ovariectomized ewes. Journal of Animal Science. 68: 1097-1102. [ Links ]
Thiéry, J.C., Martin, G.B., Tillet, Y., Caldam, M., Quentin, M., Jamain, C, Ravault, J.P. 1989. Role of Hypothalamic catecholamines in the regulation of luteinizing hormone and prolactin secretion in the ewe during seasonal anestrus. Neuroendocrinology. 49: 80-87. [ Links ]
Thiéry, J.C., Gayrard, V., Le Corre, S., Viguié, C, Martin, G.B., Chemineau, P., Malpaux, B. 1995. Dopaminergic control of LH secretion by the A15 nucleus in anoestrus ewes. Journal of Reproduction and Fertility, Supplement 49: 285-296. [ Links ]
Thiéry, J.C., Chemineau, P., Hernández, X., Migaud, M., Malpaux, B. 2002. Neuroendocrine interactions and seasonality. Domestic Animal Endocrinology. 23: 87-100. [ Links ]
Ubuka, T., Kim, S., Huang, Y.C., Reid, J., Jiang, J., Osugi, T., Chowdhury, V.S., Tsutsui, K., Bentley, G.E. 2008. Gonadotropin-inhibitory hormone neurons interact directly with gonadotropin-releasing hormone-I and -II neurons in European starling brain. Endocrinology. 149: 268-278. [ Links ]
Valencia, Z.M., Castillo, R.H., Berruecos, V.J. 1975. Reproducción y manejo del borrego Tabasco o Peligüey. Técnica Pecuaria México. 29:66-72. [ Links ]
Valencia, J., Barrón, C, Fernández-Baca, S. 1978. Variaciones estacionales de la presentación de estros en ovejas Dorset y Criollas en México. Veterinaria México. 9: 45-50. [ Links ]
Valencia, J., Barrón, C, Fernández-Baca, S., Huerta, N., Ortiz, A. 1980. Presentación de estros en ovejas criolla a lo largo del año. Veterinaria México. 11:71-74. [ Links ]
Valencia, M., Heredia, M., González, E. 1981. Estacionalidad reproductiva en hembras Pelibuey. Memorias de la VIII Reunión de la Asociación Latinoamericana de Producción Animal. Sto. Domingo, República Dominicana, p. 137. [ Links ]
Valencia, J., Porras, A., Mejía, O., Berruecos, J.M., Trujillo, J., Zarco, L. 2006. Actividad reproductiva de la oveja Pelibuey durante la época del anestro: influencia de la presencia del macho. Revista Científica, FCV-LUZ. 16: 136-141. [ Links ]
Viguié, C, Thibault, J., Thiéry, J.C., Tillet, Y., Malpaux, B. 1997. Characterization of the short day-induced decrease in median eminence tyrosine hydroxylase activity in the ewe: temporal relationship to the changes in luteinizing hormone and prolactin secretion and short day-like effect of melatonin. Endocrinology. 138: 499-506. [ Links ]
Wade, G.N., Jones, J.E. 2004. Neuroendocrinology of nutritional infertility. American Journal of Physiology, Regulatory, Integrative and Comparative Physiology. 287: R1277-R1296. [ Links ]
Watanobe, H., Schiöth, H.B., Wikberg, J.E., Suda, T. 1999. The melanocortin 4 receptor mediates leptin stimulation of luteining hormone and prolactin surges in steroid-primed ovariectomized rats. Biochemical and Biophysical Research Communications. 257. 860-864. [ Links ]
Wayne, N.L., Malpaux, B., Karsch, F.J., 1989. Social cues can play a role in timing onset of the breeding season of the ewe. Journal of Reproduction and Fertility. 87: 707-713. [ Links ]
Werner, H., Raizada, M.K., Mudd, L.M., Foyt, H.L., Simpson, LA, Roberts, C.T., LeRoith, D. 1989. Regulation of rat brain/Hep G2 glucose transporter gene expression by insulin and insulin-like growth factor I in primary cultures of neuronal and glial cells. Endocrinology. 125: 314-320. [ Links ]
Williams, L.M., Helliwell, R.A. 1993. Melatonin and seasonality in the sheep. Animal Reproduction Science. 33: 159-182. [ Links ]
Yeates, N.T.M. 1949. The breeding season of the sheep with particular reference to its modification by artificial means using light. Journal Agriculture Science, Cambridge. 39: 1-43. [ Links ]

