










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencia forestal en México
versión impresa ISSN 1405-3586
Rev. Cien. For. Mex vol.34 no.106 México jul./dic. 2009
Artículo científico
Seeds stored in the forest floor in a natural stand of Pinus montezumae Lamb.
Fernando Carrillo–Anzures1, Gil Vera–Castillo2, Octavio Salvador Magaña–Torres1, Jim M. Guldin3 and Raymond P. Guries4
1 Campo Experimental Valle de México, C.I.R. Centro, INIFAP. Correo–e: magana.octavio@inifap.gob.mx.
2 Universidad Autónoma Chapingo.
3 USDA FS Southern Research Station.
4 University of Wisconsin–Madison.
Reception date: November 13th, 2007.
Acceptance date: August 18th, 2009.
ABSTRACT
Seed banks have been studied to a greater extent in agriculture than in forestry due to long–standing concerns about the threat to agricultural crops posed by weed species. However, in forest areas seed banks have an important influence on plant succession since the vegetation that colonizes a space after a major disturbance will arise at least partly from them. Knowledge about this condition can help land managers to prescribe site treatments that produce desired vegetation from the perspectives of wildlife habitat, reduced plant competition with crop tree species, and related management concerns. The variability of the soil seed bank of a Pinus montezumae forest was assessed using seedling–emergence method for soil samples. The number of species and seedling abundance were evaluated by sampling four plots in a natural regeneration area. A total of 43 species were recorded in the seed bank (2 trees, 17 shrubs, 1 grass and 23 herbs). Viable seeds of most species were contained in similar abundance in the humus and mineral soil layers. Dominant species in the stand (P. montezumae) and codominant species (P. ayacahuite, Abies religiosa, and Alnus firmifolia) were poorly represented in the soil seed bank which was dominated by seeds of an array of annual and perennial herbs. Regeneration of commercial species under any silvicultural method must come from current seed production, or seed produced off site, but not from the soil seed bank.
Key words: Abies religiosa, Alnus firmifolia, natural regeneration, Pinus montezumae Lamb., seedling, soil seed bank.
RESUMEN
Los bancos de semillas han sido estudiados más en la agricultura que en el área forestal debido a la preocupación sobre el peligro de que los cultivos sean dominados por malas hierbas. Tienen una gran influencia en la sucesión vegetal que coloniza un área después de un disturbio. El conocimiento sobre bancos de semillas en el suelo puede ayudar a prescribir los tratamientos que propicien condiciones de vegetación deseadas desde la perspectiva de hábitat de fauna silvestre, reducción de competencia con especies de importancia y otros problemas relacionados con el manejo forestal. La variación del banco de semillas en el suelo de un bosque de Pinus montezumae fue evaluado por medio del método de emergencia en muestras de suelo; se determinó la abundancia y el número de taxa mediante el muestreo en cuatro parcelas en un área de regeneración natural. Los resultados indican 43 especies (dos de árboles, 17 de arbustos, una de pasto y 23 de hierbas). La abundancia de semillas viables fue similar para la mayoría de ellas en la capa de humus y suelo mineral; en tanto que las dominantes del rodal (P. montezumae) y codominantes (P. ayacahuite, Abies religiosa y Alnus firmifolia), no estuvieron presentes en el banco, en el cual dominaron herbáceas anuales y perennes. La regeneración de las especies comerciales para este tipo de bosque, bajo cualquier método silvícola, debe provenir de semilla producida en el año in situ o de áreas aledañas, pero no del banco de semillas.
Palabras clave: Abies religiosa, Alnus firmifolia, regeneración natural, Pinus montezumae Lamb., plántula, banco de semillas en el suelo.
INTRODUCTION
The soil seed bank is defined as all viable seed present under and on the surface of the soil (Simpson et al., 1989). Seed banks have been studied to a greater extent in agriculture than in forest lands due to long–standing concerns about the threat to agricultural crops posed by weed species. However, seed banks in forests areas have a great influence on plant succession as the vegetation that colonizes an area following a major disturbance will arise at least partly from them. Some plant species emerging in disturbed forest areas are not found in the mature forest, suggesting an origin from migrant or buried seeds (Strickler and Edgerton, 1976). In fact, some studies suggest that germination of the seed bank is the most important process in contributing to the initial composition of plant communities following a disturbance in forested areas (McGee and Feller, 1993).
Unfortunately, long–term storage of seeds in the forest floor is more characteristic of less desirable pioneer trees, annual plants, and shrubs than of commercial tree species. Many of these species have seeds with hard, nearly impermeable coats that allow the embryos to survive for the many decades that may elapse between major fires or similar disturbances. Examples of such species include representatives from the genera Rubus, Ribes and Ceanothus as well as the short–lived pin cherry (Prunus pensylvanica L.f.) (Marks, 1974).
Seeds of a few forests species may remain viable in the humus layers beneath uncut stands for periods longer than one year. One example is Atlantic white–cedar (Chamaecyparis thyoides (L.) Britton, Stern & Poggenb.), whose seeds are stored in large quantities in the poorly aerated peats in which this species grows (Little, 1950). Another example is yellow–poplar (Liriodendron tulipifera L.) (Clark and Boyce, 1964) which seems to require high temperatures or exposure to light to germinate. The seeds of various ash species (Fraxinus spp.) also remain stored in the forest floor for one or two years because it takes them that long after dispersal to mature. With such species it is often possible to expect a seemingly miraculous regeneration after harvesting the entire seed source (Smith, 1986).
Seeds buried in the soil may be exposed to conditions suitable for germination following logging or other site disturbances. To determine species composition of the stand following such a change, it must be considered that composition, depth distribution and density of seeds buried in the litter and soil, all interact with the environmental factors, especially light and temperature. Pre–disturbance and post–disturbance stand composition may differ dramatically. Such differences emphasize the importance of buried viable seeds and seed dispersal from adjacent stands in the successional dynamics on the disturbed site. To understand the contribution of seed propagules to forest stand dynamics, it is important to know the composition and spatial distribution of seed banks and their responses to environmental conditions (Pratt et al., 1984).
Two contrasting techniques are used to estimate soil seed bank composition (Simpson et al., 1989; Brown, 1992). In the first one, physical extraction of the seeds from the soil by a combination of sieving, flotation, or air flow separation, is followed by manual identification of species using seed characteristics. However, this gives no information about viability, which, subsequently, must be established through the tetrazolium or germination tests. It also requires a 'library' of seeds of known identity in order to compare those removed from soil samples. This method probably over–estimates the soil seed bank, as it detects many unviable seeds.
In contrast, seedling emergence techniques provide an estimate of viable seeds in the soil based on germination of seeds maintained under conditions favorable for germination. These requirements are seldom completely met, as germination patterns are very sensitive to fluctuating temperatures, oxygen availability, soil texture and other factors. Thus, these techniques probably under–estimate viable buried seed abundances. Despite this limitation, for community level studies, especially where the potential number of species is high, it is a useful measure because direct counting is extremely tedious and time–consuming, and also requires that the viability be tested.
A combination of seedling emergence and direct counting methods provides a more precise estimate of the seed bank size than either technique alone (Conn et al., 1984), but this is only practical for small samples and where a seed 'library' exists.
In the coniferous forest, the severity of the environment and disturbance events determine plant regeneration strategies (Archibold, 1989). Knowledge of forest soil seed banks and their response to changes can help to understand plant succession. It will also help land managers prescribe site treatments that produce desired vegetation conditions from the perspectives of wildlife habitat, reduced plant competition with crop tree species, and related management concerns (McGee and Feller, 1993).
The viability of naturally dispersed seeds of spruces (Picea) and many pines (Pinus) normally extends into the next growing season and rarely into the second growing season (Stein et al., 1974). The well known failure of conifer seeds to persist beyond one year in seed banks has been reported for a variety of forest communities in North America, as seed bank composition studies from The United States and Canada have all documented very short resident times for major conifers (Table 1).

Mercado and Arriaga (1995) report the soil seed bank composition for a Mexican pine–oak ecosystem (Pinus lagunae (Rob.–Pass.) Passini, Quercus devia Goldman and Arbutus peninsularis Rose & Goldman). Other studies have been developed in lower montane forest (transition zone between neartic and neotropical floristic regions) of Mexico (Williams–Linera, 1993). Many studies on soil seed banks exist for tropical regions in Mexico (Guevara and Gómez–Pompa, 1972; Castro–Acuña and Guevara–Sada, 1976; Álvarez–Buylla and Martínez–Ramos, 1990; Vázquez–Yánes and Orozco–Segovia, 1990; Rico–Gray and García–Franco, 1992; Quintana–Ascencio et al., 1996; Vázquez–Yánes et al., 1996).
Flores and Pérez (1990) sampled the forest floor in stands of P. montezumae at San Juan Tetla, Puebla, Mexico, before the seed dispersal season. They found an average of 60 seeds/m2 of P. montezumae with 0% viable seed, which indicates the short period of time that seeds of this species remain viable.
Several questions about the role of the soil seed bank in stands of this species were identified: What is the abundance and diversity of the soil seed bank? Does one or a few species dominate? Are many species represented? Are species predominantly herbaceous or woody?
In order to provide answers to these questions, the purpose of this study was to characterize the soil seed bank of a Pinus montezumae forest. The specific objectives were:
1. To identify species and estimate seedling abundance that can be expected in a regenerating stand from buried seeds using the seedling–emergence method for soil samples from stands of Pinus montezumae at San Juan Tetla.
2. To determine the depth distribution of buried seeds within the soil profile beneath P. montezumae stands.
3. To assess the degree to which seed bank populations reflect the species composition of current stands.
MATERIALS AND METHODS
San Juan Tetla Experimental Station lied on the east slope of the Iztaccíhuatl volcano (19°10'N, 98° 36'W), in the Municipio of Chiautzingo, State of Puebla, Mexico. The station has 15,80 ha approximately and its altitude range goes from 3,000 to 3,600 m; it includes mainly mixed stands of P. montezumae, P. hartwegii Lindl., Abies religiosa, P. ayacahuite var. veitchii (Roezl) Shaw and various hardwood species (Boyás, 1993).
The average annual precipitation is 1,169 mm, maximum rainfall occurs in summer (June–August), decreases to very low levels in winter (December–February), and remains low until May.
The stands used in this study were located about 1.5 km east of the station headquarters and were dominated by P. montezumae, with minor and varying proportions of three other conifers: white pine (P. ayacahuite var. veitchii), yellow pine (P. teocote Schiede ex Schltdl. & Cham.) and fir (Abies religiosa).In addition, several species of hardwoods are present, including alder (Alnus firmifolia), willows (Salix oxylepis Schn. and Salix sp.) and an oak (Quercus laurina Humb. et Bonpl.). The understory is dense enough to make it difficult to walk through the stand; it was dominated by small Alnus sp., Quercus sp., Salix sp. and a lush growth of perennial and malodorous Senecio sp. and other herbaceous vegetation. The understory is sufficiently dense to make it difficult walking through the stand.
The inventory indicates that this stand averages 156.1 trees/ha with a basal area of 37.61 m2/ha, of which 110.9 trees/ha and 32.75 m2/ha were montezuma pine, which made up 70 per cent of the stems ha–1 and 87 per cent of the basal area ha–1.
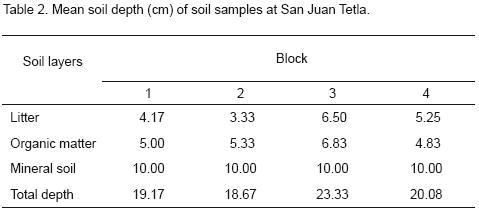
Soil samples were collected from four blocks in a natural regeneration study area. Soil and litter samples were collected on July from rectangular plots of 0.16 m2, after P. montezumae seed dispersion (Acosta and Musálem, 1986). Six systematically distributed samples were taken from each block, and all of them were separated into three layers: litter, organic matter (humus) and mineral soil (horizon A1). The litter layer at San Juan Tetla can be deep, as much as 40–50 cm at some sites, but none of the samples in this study had this depth (Table 2). A fixed mineral soil depth of 10 cm was also assessed. The samples for each litter, organic matter (humus) and mineral soil depth interval from the 6 plots were kept separate within each block, placed in plastic bags and thoroughly mixed. The six samples were divided into three subsamples (litter, organic matter and mineral soil) from each block (3 soil subsamples × 6 plot/block × 4 blocks = 72 subsamples total). The total surface area sampled was 3.84 m2.
The seedling–emergence method was chosen because it has the advantage over physical separation techniques of being less labor intensive, and does not need additional viability tests (Roberts, 1981). As forest soil contains large amounts of organic material, the seedling–emergence method is the only practical method available, and hence it has been used by most researchers (Kjellson, 1992). Finally, no 'library' of seed samples was available for species likely to be contained in the soil seed bank of San Juan Tetla.
All soil and litter samples were placed in 50 × 70 cm wooden flats in a greenhouse. Samples were watered as needed. Number and emergence time of seedlings were recorded weekly for a nine months period. Annual seedlings were removed after species identification to eliminate crowding of new emergents and to prevent seed production from mature plants.
Data analysis
Plant identification was based on the checklists made specifically for San Juan Tetla (May–Nah, 1971; Boyás, 1993). and the flora of the Valle de México, according to the similarity of ecosystems (Sánchez, 1979; Rzedowski and Rzedowski, 1985).
Comparisons between soil samples for different blocks (e.g. 1 vs 2, 1 vs 3, etc.) were tested in order to find differences in species composition among the four blocks. These similarity was calculated through Sorensen's index of similarity (Jonsson, 1993):
Cs = 2j / (a+b)
Where:
j = number of species common to both samples and
a = number of species in the sample a
b = number of species in the sample b
The index varies from 0 when both samples have no species in common, to 1 when all species occur in common in both samples.
RESULTS
The relationship between soil depth and species density is summarized for all plants in Table 3. From the soil samples, a total of 1,797 germinants representing 43 species (2 trees, 17 shrubs, 1 grass and 23 herbs) emerged. Sixteen species occurred in all four blocks (9 herbs and 7 shrubs). The estimated total seed densities varied from 307 m–2 in block 1 to 595 m–2 in block 4; the highest densities were found in blocks 3 and 4 and the lowest in blocks 1 and 2. The observed differences in density among blocks were caused primarily by the variable distribution of Senecio cinerarioides Kunth, with large seed banks of 274 seed m–2 in plot 4. Only one species of grass (Brachypodium mexicanum (Roem. & Schult.) Link) was found in blocks 1 and 2.
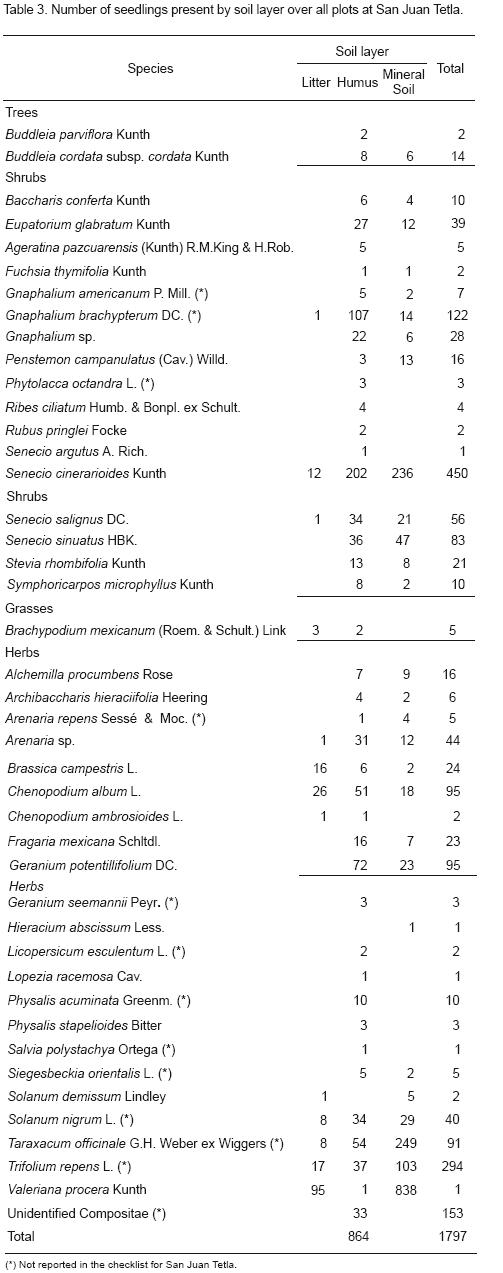
Seeds of woody species occurred in low densities. The most abundant tree was Buddleia cordata subsp. cordata Kunth, which germinated in three of the four blocks. Dicotyledonous seedlings were much more numerous than monocotyledons in all blocks. Nine herbs and three shrubs found in this study were not reported in the checklist for San Juan Tetla (May–Nah, 1971; Boyas, 1993). Seeds of canopy and shrub layer dominants (Pinus montezumae, P. ayacahuite var. veitchii, Salix sp. and Alnus firmifolia) were completely absent in the buried seed bank.
The qualitative difference between humus and mineral soil is high. Almost all species were present in the humus layer, but only 27 of the 43 species were present inthemineralsoil. The most abundant species (Seneciocinerarioides, Trifoliumrepens and an unidentified Compositae) were most abundant in the mineral soil layer. Thesecond mostabundantgroupofspecies (Gnaphaliumbrachypterum, Taraxacum officinale and Chenopodium album) were more abundant in the humus layer.
The number of germinants was significantly higher (p<0.0001) in humus and mineral soil than litter, but no significant differences between humus and mineral soil or between blocks or samples within plots were found when seed densities were comparised (Table 4).

A comparison of the relative abundance of species in seed banks, performed using Sorensen's similarity index, indicated a relatively close correspondence between the four blocks (0.65 to 0.83). The highest similarity was scored for the comparison of blocks 3 and 4, which shared 22 species.
DISCUSSION
Seed bank density.– The densities of buried seeds found in the plots of San Juan Tetla (mean for the four plots = 468 seeds m–2) were intermediately relative to densities reported for conifer forests by other authors (Ahlgren, 1979; Granstrom, 1981; Fyles, 1989). Seed densities reported in other studies have been highly variable, where some conifer forests had less than a hundred (Higo et al., 1995) to several hundreds of buried seeds m–2 (Ingersoll and Wilson, 1990; Wlliams–Linera, 1993; Zobel et al., 2007). Pratt et al. (1984) quantified over 14,000 seeds m–2 in the seed bank in an autumn collection in a coniferous forest in central Washington. Kjellsson (1992) described densities over 15,000 seeds m–2 in Danish deciduous forests. In contrast, Schiffman and Johnson (1992) found an average of 0.43 viable seeds m–2 at Jefferson National Forest, Virginia, while only 0.23 seeds m–2 were viable in the mature oak–dominated forest. Clearly, many factors influence soil seed bank densities and generalities about forest soil seed banks are impossible to make.
Seed distribution in relation to soil depth.– Most studies about seed banks in forests soils have reported a decrease of seed density with depth (Pratt et al., 1984; Archibold, 1989; McGee and Feller, 1993; Moscoso and Diez, 2005). The vertical distribution of seeds in the soil examined in the present case showed no such general trend, but distinctive distribution patterns were observed for some species. Diverse herb species (Licopersicum esculetum, Lopezia racemosa, Physalis acuminata, Physalis stapelioides, Salvia polystachya and Siegesbeckia orientalis), and several shrubs (Phytolacca octandra, Ribes ciliatum, Rubus pringlei and Senecio argutus) were totally confined to the organic matter (humus) layer, while the most abundant (Senecio cinerarioides, Trifolium repens and one unidentified Compositae) were found, mainly, in the mineral soil layer.
Species recorded deep in the soil generally have seeds with capability for long–term survival. Observations on the longevity of seeds of other Senecio species (22–100 years) (Harrington, 1972; Thompson and Grime, 1979), together with the data presented here, suggest that Senecio cinerarioides may possess a persistent seed bank. The abundance of S. cinerarioides in the soil plots reflects the high production of those plants that do become established.
The depth distributions of the seeds of different species suggest that most of the seeds of S. cinerarioides and Trifolium repens may be quite old. The actual age of even the deepest situated seeds is unknown. Continuous accumulation of plant litter, activities of soil animals, and progressive decomposition of the humus layer may all assist in moving newly produced seeds downwards from the soil surface at an unknown rate (Granström, 1981).
Small animals have been considered an important factor in herbaceous plant seed dispersal (Mladenoff, 1990; Kjellsson, 1992). Some authors (Granström, 1981; Turnbull et al., 1983) suggest that species with small seeds may have been moved downwards to the mineral soil by the activities of ants or other animals. McRill and Sagar (1973) have demonstrated that earthworms are able to ingest and relocate seeds of many small–seeded species in the soil. Earthworm activity is very low in these Pinus montezumae forests and is probably unimportant in terms of the vertical distribution of seeds. An alternative explanation for the presence of seeds in the mineral soil is that all the seeds registered in this soil layer could be located in the upper part of the mineral soil layer. No partitioning of the mineral soil samples was done, so possible seed stratification at different depths is unknown.
Seed bank composition.– With 43 species, the P. montezumae seed bank community is more diverse than most conifer forest seed banks (Pratt et al., 1984). Buried seed reserves are not extensive in coniferous forests where seeds quickly lose viability, and many other seeds are lost to predators. Studies elsewhere, reviewed by Archibold (1989) and Hills and Morris (1992) suggest that early successional plant species are better represented in soil seed banks than later successional species, as such representation allows earlier successional species to be maintained in a region that is prone to periodic disturbance.
No germinants of conifer tree species were found in this study. Dominant tree species are rarely characterized by abundant buried seed reserves, but the early successional species are well represented. Pratt et al. (1984) discovered in a seed bank of a P. ponderosa–Symphoricarpos albus (L.) Blake habitat a reversal of dominance between the study–area vegetation and the seed bank. Annual forb species, which account for relatively little cover in the stand, comprised most of the seed bank. In contrast, tree and shrub species, which dominated the vegetation, accounted for less than 1% of the seed bank. They estimated the P. ponderosa seed density in spring as 17 seeds m–2, and zero seeds m–2 in autumn prior to seed fall down. Also, in South Africa, De Villiers et al. (2003) concluded that the seed bank site would be a good source of future annual plants, but not of perennial vegetation.
Several other studies of conifer–dominated ecosystems (Kellman, 1970 and 1974; Johnson, 1975; Strickler and Edgerton, 1976; Whipple, 1978; Hill and Stevens, 1981; Mercado and Arriaga–Cabrera, 1995) have reported uncommon germination of conifer seeds. In a Pinus–Quercus forest in the state of Baja California Sur, México, Mercado and Arriaga (1995) found that the primary species of this forest (Pinus lagunae M.–F. Passini, Quercus devia Goldman and Arbutus peninsularis Rose & Goldman) were not present in the soil seed bank. Zobel et al. (2007) studied a mesophyte mixed spruce forest and found a poor correspondence between the vegetation and the seed bank. P. montezumae, the dominant conifer in the study area, generally produces good seed year crops at 3–5 year intervals (Acosta and Musálem, 1986), and its seeds remain viable for 7–12 months (Flores and Pérez, 1990). P. ayacahuite produces good seed crops annually, and they remain viable for about the same period mentioned above (Serrano, 1986).
Species of Alnus have been reported to retain their viability for only a short period of time (McGee and Feller, 1993), which could explain why no germinants of alder were found in the study.
The presence of Solanum spp., Fuchsia thymofila and Senecio spp. in the soil seed bank suggest a persistency of these species, while others with low density and in few samples comprise the transient ones.
Sampling was carried out a few years after a good seed crop of P. montezumae, and several months after seeds of P. ayacahuite were dispersed. One important feature of some plants is large seeds, which however makes them susceptible to seed predation (Eriksson, 1995). In a comparative study of small and large seeded species, Reader (1993) found that only species with seeds larger than 0.3 mg were negatively affected by seed predators. Seeds of P. montezumae and P ayacahuite are well above this limit, suggesting that they may suffer from heavy predation.
Increasing seed size makes species more likely to succeed in recruitment under litter, but less likely to be preserved in persistent seed banks. Seed predators are more likely to affect large seeded species, so decreasing seed size enhances development of seed banks, but makes species dependent on factors which temporarily remove the limiting effects of litter.
The results of the experiment suggest that for all species examined here, mineral soil was equal to forest floor (litter and humus) as a germination substrate. Removal of the forest litter and organic matter would remove about one–half of the soil seed bank. If exposure of mineral soil provides a generally superiorsubstrate for seed germination, loss of half of the seed bank might not necessarily lead to a reduction in vegetation cover in the early successional plant community. Only a few species were restricted to just litter and organic matter, so species diversity would not be changed dramatically.
Caution must be applied in extrapolating seed bank studies to potential seedling populations in the field, partly because of problems in determining soil seed banks and because species establishment from a seed bank is a function of the depth to which the soil is disturbed. Burning, removal, or redistribution of the forest floor will result in different impacts on the seed bank and different potential plant populations (McGee and Feller, 1993). Burning alone can remove different amounts of forest floor depending on the severity of the fire, as has been found in southwestern British Columbia and elsewhere (Feller, 1989).
CONCLUSIONS
Seed density and species diversity of the seed bank in Pinus montezumae stands sampled for this study compared with literature reports were average, facts that are attributed to the high density of shrubs and herbs.
Dominant species (P. montezumae) and codominant species (P. ayacahuite var. veitchii, Abies religiosa and Alnus firmifolia) in the stand were not comparable with species dominance in the seed bank.
Viable seeds of most species were contained in similar abundance in the humus and mineral soil layers.
Regeneration of desirable (commercial) species under any regeneration cutting method will be expected from seed production in the current or future years, or few seeds produced off site, but none will be expected from the soil seed bank.
REFERENCES
Acosta M., M. y M. A. Musálem. 1986. Dispersión de semillas de Pinus montezumae Lamb., en dos matarrasas del Campo Experimental Forestal San Juan Tetla, Puebla. Revista Chapingo 52: 35–40. [ Links ]
Ahlgren, C. E. 1979. Buried seed in the forest floor of the boundary waters canoe area. Minn. For. Res. Note 271. 4 p. [ Links ]
Álvarez–Buylla, E. R. and M. Martínez–Ramos. 1990. Seed bank versus seed rain in the regeneration of a tropical pioneer tree. Oecologia 84(3):314–325. [ Links ]
Archibold, O. W. 1989. Seed banks and vegetation processes in coniferous forests. In: Leck, M. A., Parker, V. T. and R. L. Simpson (Eds.). Ecology of Soil Seed Banks. Academic Press, San Diego, Calif., pp. 107–122. [ Links ]
Boyás D., J. C. 1993. Flora fanerogámica del Campo Experimental Forestal San Juan Tetla, Puebla. Boletín Técnico No.114. (2a. Ed.) INIFAP. México, D. F. México. 103 p. [ Links ]
Brown, D. 1992. Estimating the composition of a forest seed bank: a comparison of the seed extraction and seedling emergence methods. Can. J. Bot. 70: 1603–1612. [ Links ]
Castro–Acuña, R. y S. Guevara–Sada. 1976. Viabilidad de semillas en muestras de suelo almacenado de 'Los Tuxtlas', Veracruz. In: Gómez–Pompa, A., C. Vázquez–Yanes, S. del Amo–Rodríguez y A. Butanda–Cervera (Eds.). Investigaciones sobre la regeneración de selvas altas en Veracruz, México. INIREB/Compañía Editorial Continental, México, D. F. México. pp. 233–249. [ Links ]
Clark, B. F. and S. G. Boyce. 1964. Yellow–poplar seed remain viable in the forest litter. J. For. 62: 564–567. [ Links ]
Conn, J. S., C. L. Cochrane and J. A. DeLapp. 1984. Soil seed bank changes after forest clearing and agricultural use in Alaska. Weed Sci. 32: 343–347. [ Links ]
De Villiers, A. J., M. W. Van Rooyen and G. K. Theron. 2003. Similarity between the soil seed bank and the standing vegetation in the Strandveld Succulent Karoo, South Africa. Land Degradation & Development 14(6): 527–540. [ Links ]
Eriksson, O. 1995. Seedling recruitment in deciduous forest herbs: the effects of litter, soil chemistry and seed bank. Flora 190: 65–70. [ Links ]
Feller, M. C. 1989. Estimation of nutrient loss to the atmosphere from slashburns in British Columbia. In: Maclver, D. C, H. Auld and R. Whitewood (Eds.). Proceedings of the 10th. Conference on Fire and Forest Meteorology. Environment Canada, Ottawa, Canada, pp. 126–135. [ Links ]
Flores A., E. y J. L. Pérez B. 1990. Evaluación del banco de semillas en un rodal de Pinus montezumae Lamb. In: Memoria de la Reunión de los 10 años de investigación forestal en la Región Central de México. SARH– INIFAP–CIRCE. Metepec, Edo. de Méx. México, pp. 11. [ Links ]
Frank, R. M. and I. O. Safford. 1970. Lack of viable seeds in the forest floor after clearcutting. J. For. 68: 776–778. [ Links ]
Fraser, J. W. 1976. Viability of black spruce seed in or on a boreal forest seedbed. For. Chron. 52:229–231. [ Links ]
Fyles, W. M. 1989. Seed bank populations in upland coniferous forest in central Alberta. Can. J. Bot. 67: 274–278. [ Links ]
Granström, A. 1981. Seed banks in five boreal forest stands originating between 1810 and 1963. Can. J. Bot. 60:1815–1821. [ Links ]
Guevara S., S. and A. Gómez–Pompa. 1972. Seeds from surface soils in a tropical region of Veracruz, Mexico. J.Arnold Arbor. 53: 312–335. [ Links ]
Haig, I. T. 1932. Premature germination of forest tree seed during natural storage in the duff. Ecology 13:311–312. [ Links ]
Haig, I. T., K. P. David and R. H. Weidman. 1941. Natural regeneration in the Western white pine type. U.S. Dept. Agr. Tech. Bui. 767. 99 p. [ Links ]
Harrington, J. F 1972. Seed storage and longevity. In: Kozlowski, T. T. (Ed). Seed Biology, Vol. 3. Academic Press, New York, NY. USA. pp. 145–245. [ Links ]
Higo, M., A. Shinohara and S. Kodama. 1995. The regeneration behavior of major component species in the secondary forest dominated by Pinus densiflora and Quercus serrata in central Japan. Forest Ecology and Management 76:1–10. [ Links ]
Hill, M. O. and P. A. Stevens. 1981. The density of viable seed in soils of forest plantations in upland Britain. J. Ecol. 69: 693–709. [ Links ]
Hills, S. C. and D. M. Morris. 1992. The function of seed banks in northern forest ecosystems: a literature review. Ont. Minist. Nat. Resour. For. Res. Inf. Pap. No. 107. 25 p. [ Links ]
Hofmann, J. V. 1917. Natural reproduction from seed stored in the forest floor. Jour. Agr. Res. 11:1–26. [ Links ]
Ingersoll, C. A. and M. V. Wilson. 1990. Buried propagules in an old–growth forest and their response to experimental disturbances. Can. J. Bot. 68:1156–1162. [ Links ]
Isaac, L. A. 1935. Life of Douglas fir seed in the forest floor. J. For. 33:61–66. [ Links ]
Johnson, A. E. 1975. Buried seed populations in the subartic forest east of Great Slave Lake, Northwest Territories. Can. J. Bot. 48: 2933–2941. [ Links ]
Jonsson, B. G. 1993. The bryophyte diaspore bank and its role after small–scale disturbance in a boreal forest. Journal of Vegetation Science Vol. 4(6): 819–826. [ Links ]
Kellman, M. C. 1970. The viable seed content of some forest soils in coastal British Columbia. Can. J. Bot. 48: 1383–1385. [ Links ]
Kellman, M. C. 1974. Preliminary seed budgets for two plant communities in coastal British Columbia. J. Biogeogr. 1: 123–133. [ Links ]
Kjellson, G. 1992. Seed banks in Danish deciduous forests: species composition, seed influx and distribution pattern in soil. Ecography 15:86–100. [ Links ]
Kramer, N. B. and F. D. Johnson. 1987. Mature forest seed banks of three habitat types in central Idaho. Can. J. Bot. Vol. 65 (9): 1961–1966. [ Links ]
Little, S. Jr. 1950. Ecology and silviculture of white cedar and associated species in southern New Jersey. Yale Univ. Sch. For. Bull. 56. 103 p. [ Links ]
Marks, P. L. 1974. The role of Pin cherry (Prunus pensylvanica L.) in the maintenance of stability in northern hardwood ecosystems. Ecol. Monogr. 44: 73–88. [ Links ]
May–Nah, A. 1971. Estudio fitosociológico del Campo Experimental Forestal San Juan Tetla, Estado de Puebla, México. Tesis Profesional. Escuela Nacional de Ciencias Biológicas. Instituto Politécnico Nacional. México, D. F. México. 130 p. [ Links ]
McGee, A. and M. C. Feller. 1993. Seed banks of forested and disturbed soils in southwestern British Columbia. Can. J. Bot. 71: 1574–1583. [ Links ]
McRill, M. and G. R. Sagar. 1973. The ingestion of weed seed by earthworms. Proceedings of the 12th. British Weed Control Conference, Brighton, UK. Vol II: 519–524. [ Links ]
Mercado, C. and L. Arriaga–Cabrera. 1995. Seed bank and treefall gaps in a Mexican temperate forest. In: DeBano, L. F., P. F. Ffolliott, A. Ortega–Rubio, G. J. Gottfried, R. H. Hamre and C. B. Edminster (Eds.): Conference on the Biodiversity and Management of the Madrean Archipelago: The Sky Islands of the Southwestern United States and Northwestern Mexico. U.S.D.A. Rocky Mountain Forest and Range Experimental Station. General Technical Report RM–GTR–264. Fort Collins, CO. USA. pp. 599–601. [ Links ]
Mladenoff, D. J. 1990. The relationship of the soil seed bank and understory vegetation in old–growth northern hardwood–hemlock treefall gaps. Can. J. Bot. 68:2714–2721. [ Links ]
Moscoso M., L. V. y M. C. Diez G. 2005. Banco de semillas en un bosque de roble de la cordillera central colombiana. Rev Fac. Nal. Agr. 58(2): 2931–2944. [ Links ]
Pratt, D. W, R. A. Black and B. A. Zamora. 1984. Buried viable seed in a ponderosa pine community. Can. J. Bot. 62: 44–52. [ Links ]
Quick, C. R. 1956. Viable seeds from the duff and soil of sugar pine forest. For. Sci. 2:36–42. [ Links ]
Quintana–Ascencio, P. F., M. González–Espinoza, N. Ramírez–Marcial, G. Domínguez–Vázquez and M. Martínez–Ico. 1996. Soil seed banks and regeneration of tropical rain forest from milpa fields at the Selva Lacandona, Chiapas, Mexico. Biotropica 28(2): 192–209. [ Links ]
Reader, R. J. 1993. Control of seedling emergence by ground cover and seed predation in relation to seed size for some old–field species. J. Ecol. 81:169–175. [ Links ]
Rico–Gray, V and J. G. García–Franco. 1992. Vegetation and soil seed bank of successional stages in tropical lowland deciduous forest. J. Veg. Sci. 3:617–624. [ Links ]
Roberts, H. A. 1981. Seed banks in soils. In: Coaker, T. H. (Ed.). Advances in Applied Biology VI. Academic Press, Ltd. London, UK. pp. 1–55. [ Links ]
Rzedowski, J. y G. C. de Rzedowski. 1985. Flora fanerogámica del Valle de México I. Instituto de Ecología. A. C. Pátzcuaro, Mich., México. 1406 p. [ Links ]
Sánchez S., C. 1979. La flora del Valle de México. Editorial Herrero, S. A. de C. V México, D. F. México. 519 p. [ Links ]
Schiffman, P. M. and W. C. Johnson. 1992. Sparse buried seed bank in a Southern Appalachian oak forest: Implications for succession. Am. Midi. Nat. 127:258–267. [ Links ]
Serrano J., M. S. 1986. Efecto de algunos factores ambientales en la germinación de semillas de Pinus ayacahuite var. veitchii, bajo condiciones controladas. Tesis Profesional. Universidad Autónoma Chapingo. Chapingo, Edo. de Méx. México. 72 p. [ Links ]
Simpson, R. L., M. A. Leck and V. T. Parker. 1989. Seed banks: general concepts and methodological issues. In: M. A. Leck, V. T. Parker and R. L. Simpson (Eds.): Ecology of Soil Seed Banks. Academic Press. New York, NY. USA. pp. 3–8. [ Links ]
Smith, D. M. 1986. The practice of silviculture. John Wiley & Sons. Inc. New York, NY. USA. 527 p. [ Links ]
Stein, W. I., P. E. Slabaugh and A. P. Plummer. 1974. Harvesting, processing and storage of fruits and seeds. In: Schopmeyer, C. S. (Technical Coordinator). Seeds of woody plants in the United States., Agriculture Handbook No. 450, U.S. Department of Agriculture. Washington, DC. USA. pp. 98–125. [ Links ]
Strickler, G. S. and P. J. Edgerton. 1976. Emergent seedlings from coniferous litter and soil in Eastern Oregon. Ecology 57(4): 801–807. [ Links ]
Thomas, P. A. and R. W. Wein. 1985. Delayed emergency of four conifer species on postfire seedbeds in eastern Canada. Can. J. For. Res. 15: 727–729. [ Links ]
Thompson, K. and J. P. Grime. 1979. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. The Journal of Ecology. Vol. 67(3): 893–921. [ Links ]
Turnbull, C. L., A. J. Beattie and F. M. Hanzawa. 1983. Seed dispersal by ants in the Rocky Mountains. Southwest. Nat. 28:289–293. [ Links ]
Vázquez–Yánes, C. and A. Orozco–Segovia. 1990. Seed dormancy in the tropical rain forest. In: Bawa, K. S. and M. Hadley. (Eds.) Reproductive ecology of tropical rainforest plants. UNESCO and Parthenon. MAB Vol. 7. pp. 247–259. [ Links ]
Vázquez–Yánes, C. and M. Rojas–Aréchiga. 1996. Ex situ conservation of tropical rain forest seed: problems and perspectives. Interciencia 21: 293–298. [ Links ]
Whipple, S. A. 1978. The relationship of buried germinating seeds to vegetation in an old–growth Colorado subalpine forest. Can. J. Bot. 56: 1505–1509. [ Links ]
Williams–Linera, L. G. 1993. Soil seed banks in four lower montane forest of Mexico. Journal of Trop. Ecol. 9: 321–337. [ Links ]
Zasada, J. C., R. A. Norum, R. M. Van Veldhuisen and C. E. Teutsch. 1983. Artificial regeneration of trees and tall shrubs in experimentally burned upland black spruce/feather moss stands in Alaska. Can. J. For. Res. 13: 903–913. [ Links ]
Zobel, M., R. Kalamees, K. Püssa, E. Roosaluste and M. Moora. 2007. Soil seed bank and vegetation in mixed coniferous forests stands with different disturbance regimes. Forest Ecology and Management 250(1–2): 71–76. [ Links ]

