










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.43 no.7 Texcoco oct./nov. 2009
Recursos naturales renovables
Regeneration of Abies religiosa in canopy gaps versus understory, Cofre de Perote National Park, México
Regeneración de Abies religiosa en claros del dosel versus sotobosque, Parque Nacional Cofre de Perote, México
Rogelio Lara–González1,2, Lázaro R. Sánchez–Velásquez1,*, Juan Corral–Aguirre2
1 Instituto de Biotecnología y Ecología Aplicada. Instituto de Genética Forestal. Dirección General de Investigaciones, Universidad Veracruzana, Campus para la Cultura, las Artes y el Deporte, Avenida de las Culturas Veracruzanas No. 101, Colonia Emiliano Zapata, Apartado Postal 250, 91090. Xalapa, Veracruz. México. *Autor responsable: (rolago30@hotmail.com).
2 Facultad de Biología, Zona Universitaria, Xalapa, Veracruz, Universidad Veracruzana. Email: (lasanchez@uv.mx).
Received: May, 2008.
Approved: January, 2009.
ABSTRACT
Some studies have shown that fir (Abies spp.) regeneration can occur either in canopy gaps or in understory. The purpose of this study was to know how canopy gaps contribute to fir regeneration. Points were randomly chosen in three fir forests, and starting from there the nearest gap was recorded. Likewise, other random samples were placed in understory with the same diameter as that of the gap registered before. Gap age, gap size, and tree seedling for each sample were recorded. The average gap age was 7.61 ±1.86 yr. The average gap size was 66±42.8 m2. The most common gaps were of intermediate size (44–88 m2). Only seedlings of A. religiosa (H.B.K.) Schlecht. & Cham and Pinus montezumae Lamb, were observed either in gaps or in understory. The number of seedlings was higher in gaps than in understory (p<0.001). The average number of seedlings in gaps was 4 430 ±596 per ha (A. religiosa 95.7 % and R montezumae 4.3 %), whereas in understory it was 993±316 pef ha (A. religiosa 99 % and P. montezumae 1 %). There was a significant relationship between gap age and number of seedlings of A. religiosa (R=0.62, p<0.001). The gap size and the interaction gap size–age were not significant (p>0.23) and did not explain the regeneration density.
Key words: Abies religiosa, canopy gap, disturbances, fir forest, México, regeneration.
RESUMEN
Algunos estudios han mostrado que la regeneración del abeto (Abies spp.) puede ocurrir tanto en claros del dosel y en el sotobosque. El propósito de este estudio fue saber cómo contribuyen los claros del dosel a la regeneración del abeto. Se eligieron puntos aleatorios en tres bosques de abetos y se registró el claro más cercano a cada punto. Asimismo, bajo el dosel se realizó otro muestreo aleatorio con el diámetro igual al claro antes registrado. Se registró la edad y el tamaño de los claros, antes de plántulas para cada muestra. La edad promedio de los claros fue 7.61 ± 1.86 año. El tamaño promedio de los claros fue 66±42.8 m2. Los claros más comunes fueron de tamaño intermedio (44–88 m2). Las plántulas de A. religiosa (H. B. K.) Schlecht & Cham y de Pinus montezumae fueron las únicas observadas en los claros o en el sotobosque. El número de plántulas fue mayor en claros que en sotobosque (p<0.001). El número promedio de plántulas en los claros fue 4,430 ±596 por ha (A. religiosa, 95.7% y R Montezumae, 4.3%), mientras que en el sotobosque fue 993±316 por ha (A. religiosa, 99% y R Montezumae, 1 %). Hubo una relación significativa entre la edad de los claros y el número de plántulas de A. religiosa (R=0.62, p<0.001). El tamaño de los claros y la interacción tamaño–edad del claro no fueron significativos (p>0.23) y no explicaron la densidad de regeneración.
Palabras clave: Abies religiosa, claro del dosel, disturbios, bosque de abetos, México, regeneración.
INTRODUCTION
Forests constantly change in structure due to disturbances on a large scale (e.g. fire and clear cutting) as well as on a small scale (e.g. trees brought down by storms or by the fall of branches) (Spies and Franklin 1989; Pham et al, 2004; Worrall etal., 2005). Disturbance in the canopy can produce small gaps which maintain the regeneration of some tree species. In gaps, the regeneration process can include seed rain, seed bank or seedling bank, which activate their development by improving the light available (Coomes and Grubb 2003; Martinez–Ramos, 1994; Arrieta and Suárez 2005).
In México, Abies religiosa (H.B.K.) Schlecht. & Cham, known as oyamel is a dominant specie in woodlands (fir) forests. These forests are considered relicts and their altitudinal range is from 2000 to 3600 m. It is estimated that fir forest areas in México represent only 0.16 %. There are eight species and five varieties of Abies in México (Manzanilla, 1974). The Mexican fir forests have relatively dense canopies; light that reaches the soil is scarce and the understory is sparse (Challenger and Caballero, 1998). The fir species that is more widespread in México is A. religiosa. However, some Abies populations are being fragmented and reduced (Sánchez–Velasquez et al, 1991; Aguirre–Planter et al, 2000; Brower et al., 2002). Usually, under the canopy of fir forests, A. religiosa (like other species of Abies) is a shade–tolerant species (Rzedowski, 1978; Spurr and Barnes, 1982; Whitmore, 1989). Nevertheless, some species of Abies can regenerate in canopy gaps as well as in understory (Narakawa and Yamamoto, 2001; Sugita andTani, 2001; Mori andTakeda, 2002). The question posed in this study was: how do the canopy gaps contribute to fir regeneration? Our hypothesis was that fir regeneration is better in canopy gaps than in understory.
MATERIALS AND METHODS
Study area
This research was carried out at the Cofre de Perote, National Park, ejido El Conejo, Veracruz, México, located at 19° 31' 54.5" N and 97° 09' 14.8" W, at an elevation of 3140 m. The climate is temperate cold–sub–humid, with temperatures mean annual between –5 and 22 °C (max. and min.), summer rainfalls and a mean annual precipitation between 1200 and 1500 mm (Medina and Ángulo, 1990). The fir forest has a basal area of 70 m2 ha–1, approximately (A. religiosa trees are larger than 10 cm dbh) (Sánchez–Velásquez et al., 1991), and these are forest belts located on a volcano. The basal area of Pinus montezumae is around 1 m2 per ha. The soil type is andosol derived from volcanic ash which is characterized by a dark surface layer and is rich in humus. The susceptibility of soil to erosion is high and increases in the case of a clear cutting because of its hillside position (Narave, 1985). Abies religiosa is used as Christmas trees, and the adult trees provide wood for housing, furniture, building, roofing, firewood, and fences.
Samples and analysis
Three transects were randomly traced out in three fir forests, choosing randomly 30 points on all transects, and at each point a direction was also randomly taken (right or left from the transect). On this route, a maximum distance of 50 m was covered, and the gap nearest to the point was registered. The minimum size considered for the gap was 5 m in diameter. In each gap, gap size, number of seedlings per species, and gap age were recorded. Gap age was recorded as the time passed since the creation of the gap, and estimated by the number of whorls of the individuals regenerating within the gap; whorls in Abies are produced at the rate of one whorl per year (Silvertown and Doss, 1999; Rasmussen etal., 2003; Earle, 2007). Furthermore, acore sample was taken with a Swedish increment borer from two trees located around each gap in order to find the age of the gap. In these samples, the growth ring liberation could be observed, which coincided with the age (number of branch whorls) of advanced regeneration; each ring is equivalent to one year.
In order to estimate the size (m2) of each gap, two diameter measures were taken in the gap, the shortest diameter (SD) and the longest diameter (LD). Afterwards, the average radius (m) was estimated as (SD+LD)/4. The gap area was estimated based on the circle area formula (A=pr2). The boundary gap was defined by looking up to the margin of the opening using a clinometer (Suunto, Forestry Supplier, 205 West Rankin ST, P.O. Box 8397, MS, USA). Likewise, from each boundary gap sample, in understory, a cardinal direction was randomly chosen and at a distance of 20 m another sample was placed with the same diameter as that of the gap registered before. In this sample, the number of seedlings per species was recorded.
Applying multiple regressions and correlations step by step (deviance), the relationships between density (number of A.religiosa seedlings between >0.2 and <2 m tall), gap size and gap age were examined. Furthermore, the same method was used to explore the relationship between two size categories (seedlings with fewer than 3 whorls, and seedlings with three or more whorls) and gap size and gap age. This categorization was made arbitrarily, considering that the number of youngest seedlings (those of the first category) might explain better the relationship between the gap size and the number of seedlings. Also, regression analyses were used between number of seedlings observed in understory and size area. In all cases, the number of seedlings was projected per hectare.
With the Pearson Association Index (Ip), corrected by Yates (Greig–Smith 1983), the association between seedlings of different species was estimated. The comparison between numbers of seedlings in gaps versus number of seedlings in understory (control sample) was estimated by a χ2 test.
RESULTS AND DISCUSSION
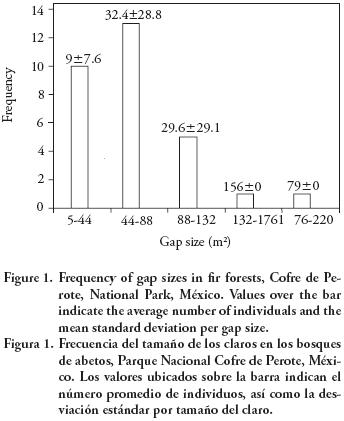
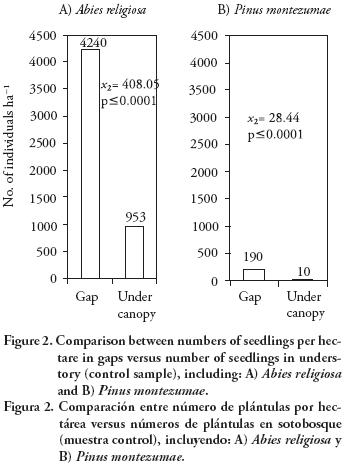
The average gap age was 7–61 ±1.86 yr. The range of gap sizes recorded was 19.6–219 m2 with an average of 66±42.8 m2. The most common gaps were those of intermediate size (44–88 m2) (Figure 1). Only seedlings of A. religiosa and P. montezumae were observed either in gaps or in understory. The number of A. religiosa seedlings as well as of P. montezumae was considerably higher in gaps than in understory (p<0.001) (Figure 2). The average number of seedlings in gaps was 4430 ±596 per ha (A. religiosa 95.7 % and P. montezumae 4.3 %), whereas in understory it was 993±316 per ha (A. religiosa 99 % and 1 % of P. montezumae). There was no significant association between A. religiosa and P. montezumae (Ip), in gaps (Ip=0.00; χ2 <0.01; p>0.05) nor in understory (Ip=0.13; χ2=0–54; p>0.05).
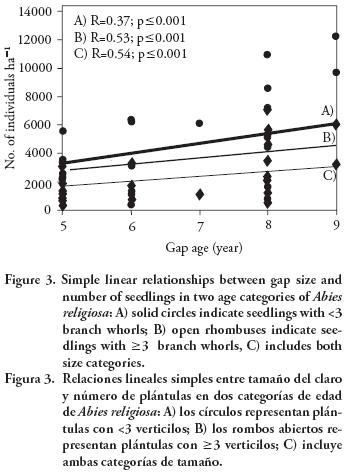
A significant linear relationship between gap age and total number of seedlings per ha of A. religiosa was observed (R=0.54; p<0.001), which is represented by the equation: Y=678.1 X; where: X=gap age (years), and Y=number of seedlings of A. religiosa (Figure 3). The correlations between gap age and the two categories of regeneration size, i.e., seedlings >3 whorls and seedlings >3 whorls (ha), also showed significant relationships (R=0.37, p<0.001 and R=0.53, p<0.001). The equation for: A) seedlings with fewer than three whorls and, B) seedlings with three or more whorls were Y=340.24 X and Y=337.8 X (both p<0.001) (Figure 3). Significant differences were not observed between either regression linear slopes (p=0.75). When the gap size and the interaction gap size–age were added in the regression model, no significant contribution (p>0.1) was observed to explain the total number of seedlings nor the number of seedlings per size class (number of branch whorls). The relationship between size (area) and regeneration density in the understory were not significant in all cases (p>0.05).
The genus Abies has species with high shade tolerance (eg. A. balsamea (L.) Mill., Kneeshaw et al., 2006) as well as low shade tolerance (eg. A. sachalinensis (F. Schmidt) Mast. Kubota and Hara, 1996). Some studies (Rzedowski, 1978; Sánchez–Velásquez et al., 1991; Figueroa and Moreno, 1993; Challenger and Caballero, 1998) suggest that in A. religiosa forests, regeneration occurs in the understory. In contrast, the results of this study advance that A. religiosa regeneration is considerably higher in gaps than in understory. This data agrees with other species of tropical forests (Denslow, 1980, 1987; Whitmore, 1984; Schupp etal., 1989; Schnitzer and Carson 2001), as well as in old–growth mesic forest (Poulson and Platt 1989; Runkle 1989; Ott and Juday 2002), where regeneration in gaps was higher. However, some species of Abies can have seedlings both in canopy gaps and in understory (Narakawa and Yamamoto, 2001; Sugita and Tani, 2001; Mori and Takeda, 2002). Nevertheless, in order to evaluate the specific contributions of regeneration (gaps ps understory) on the population growth rate (λ), a long term study is necessary.
According to González et al. (1991), A. religiosa also grows in exposed sites in smaller proportions than understory. It has been observed that in open areas the regeneration is more successful under the canopy of Baccharis conferta Kunth. This suggests that B. conferta shrub is a nurse species which facilitates fir regeneration (Snook 1993).
Gap size is important in the regeneration process (Brockway and Outcalt, 1998; Schnitzer and Carson, 2001; Ott and Juday, 2002), since light and temperature of the soil are dependents of gap size. The gap sizes in this study were between 19–6 and 219 m2, and they are similar to those registered by Runkle (1981), Narakawa and Yamamoto (2001), Ott and Juday (2002), Clinton (2003), and Pham et al. (2004). This gap size range can be classified as small gaps according to Coates (2002), who used the following categories: small (20–300 m2), intermediate (301–1000 m2), and large gaps (1001–5000 m2), or as gaps of fine scale (<1000 m2) according to Spies and Franklin (1989). However, our results suggest that regeneration in gaps occur in pulses as time goes by, because there is a strong relationship between number of seedlings and gap age, but not between number of seedlings and gap size, not number of seedlings in understory and size area.
Fir forests are characterized by their simple structure (one to tree species); they are seldom found mixed with other tree species, sometimes with late successional species and more frequently with early successional species (Rzedowski, 1978; Spurr and Barnes, 1982; Challenger and Caballero, 1998). In this study, besides A. religiosa, only P. montezumae seedlings were observed, mainly in the gaps, but their densities are low. Pinus montezumae did not show significant association with A. religiosa. Perhaps this is due to the fact that some pine species require gaps larger than 1000 m2 (White, 1979), and under the situation of these large gaps, the fir forest might be replaced by pine forest (Romme and Knight 1981; Keeling et al. 2006). In other ecosystems, with the presence of large–scale disturbances (fire and complete clear cutting), some pine species can replace the cloud forest. However, if there are not disturbances, pines can be replaced by cloud forest species (Sánchez–Velásquez, 1986; Jardel, 1991; Sánchez–Velasquez and García–Moya, 1993).
In sum, the regeneration of A. religiosa is more successful in gaps than in understory. Seedlings with more than three whorls are mostly associated to the gap age. Pinus montezumae regeneration in the gaps was lowest than A. religiosa. For the management of these forests, the method of tree selection could be tested, choosing big trees to create regeneration conditions and guarantee a sustainable production of timber and Christmas trees.
CONCLUSIONS
The regeneration of fir forests in Cofre de Perote National Park, México, is higher in canopy gaps (95.7 %) than in understory. Our results are different from some studies which suggest that fir regeneration only occurs in understory. However, a demographic study is necessary to know the true contributions of regeneration, such in canopy gaps as understory, on population growth rate. Our hypothesis was not rejected, i.e. the fir regeneration is better in canopy gaps than in understory.
ACKNOWLEDGEMENTS
Thanks to Don Remigio Domínguez and his kind family for their hospitality and the facilities given to carry out this work. This research was financed by Sectorial Funds of CONAFOR –CONACYT (code: 2002–C01–6163) and PRODUCE–Ver Foundation (PRODUVER) through the project: Plantations, ecology, and demography of Abies religiosa and A. hickelii: two potential species for Christmas tree production at Cofre de Perote National Park. Thanks for the revision of the manuscript to Ma. del Rosario Pineda-López, Jorge Galindo-González, Francisco Diaz Fleischer, Jerico Sánchez, Lorena Sánchez, Agrociencia’s editor, and two anonymous reviewers who revised and improved the last manuscript.
LITERATURE CITED
Aguirre-Planter, E., G.R. Furnier, and L.E. Eguiarre. 2000. Low level of genetic variation within and high level of genetic differentiation among populations of species of Abies from southern México and Guatemala. Am J. Bot. 87:362-371. [ Links ]
Arrieta, S., and F. Suárez. 2005. Marginal holly (Ilex aquifolium L. populations in Mediterranean central Spain are constrained by low–seedling recruitment. Flora 201:152–160. [ Links ]
Brockway, D., G., and K.W. Outcalt. 1998. Gap–phase regeneration in longleaf pine wiregrass ecosystems. For. Ecol. Manage. 106:125–139. [ Links ]
Brower, L., P., G. Castilleja, and A. Peralta. 2002. Quantitative changes in forest quality in a principal over wintering area of the monarch butterfly in Mexico, 1971–1999. Conserv. Biol. 16:346–359. [ Links ]
Challenger, A., y J. Caballero. 1998. Utilización y Conservación de los Ecosistemas Terrestres de México: Pasado, Presente y Futuro. CONABIO, Instituto de Biología. Sierra Madre, México, D. F. 847 p. [ Links ]
Clinton, B., D. 2003. Light, temperature, and soil moisture responses to elevation, evergreen understory, and small canopy gaps in the southern Appalachians. For. Ecol. Manage. 186:243–255. [ Links ]
Coates, K., D. 2002. Tree recruitment in gaps of various sizes, clearcuts and undisturbed mixed forest of interior British Columbia, Canada. For. Ecol. Manage. 155:387–398. [ Links ]
Coomes, D., A., and P.J. Grubb. 2003. Colonization, tolerance, competition and seed–size variation within functional groups. Trends in Ecol. Evol. 18:283–291. [ Links ]
Denslow, J., S. 1980. Gap partitioning among tropical rainforest trees. Biotropica 12:47–55. [ Links ]
Denslow, J., S. 1987. Tropical rainforest gaps and tree species diversity. Ann. Rev. Ecol. Syst. 18:431–451. [ Links ]
Earle, C., J. 2007. The Gymnosperm Database. http://conifers.org/pi/ab/. Cited 27 Dec 2007. [ Links ]
Figueroa, R., y S. Moreno M. 1993. Estructura y sucesión en poblaciones de Abies religiosa y Abies religiosa var. emarginata en la Sierra de Manantlán, Jalisco. Agrociencia Ser. Rec. Nat. 3:49–63. [ Links ]
González, G., M. Musalem, M. A. Zarate, G. Lara y M.A. Velásquez. 1991. Estudios de germinación en Oyamel (Abies religiosa (H.B.K.) Schl et Cham) bajo ambientes naturales en Zoquiapan, México. Rev. Chapingo 15:59–66. [ Links ]
Greig–Smith, P. 1983. Quantitative Plant Ecology. Blackwell Scientific Publications. Oxford. 288 p. [ Links ]
Jardel, E., J. 1991. Perturbaciones naturales y antrópicas y su impacto en la dinámica sucesional en los bosques de Las Joyas de la Sierra de Manantlán, Jalisco. Tiempos de Ciencia 22:9–26. [ Links ]
Keeling, E., G. A. Sala, and T. H. DeLuca. 2006. Effects of fire exclusion on forest structure and composition in unlogged ponderosa pine/Douglas–fir forests. For. Ecol. Manage. 237:419–428. [ Links ]
Kneeshaw, D., D. R. K. Kobe, and K. D. Coates. 2006. Sapling size influences shade tolerant ranking among southern boreal tree species. J. Ecol. 94:471–480. [ Links ]
Kubota, Y., and T. Hara. 1996. Recruitment processes and species coexistence in a sub–boreal forest in northern Japan. Ann. Bot. 78:741–748. [ Links ]
Manzanilla, H. 1974. Investigación Epidométrica y Silvicultura de Bosques Mexicanos de Abies religiosa. SAG, México. 345 p. [ Links ]
Martínez–Ramos, M. 1994. Regeneración natural y diversidad de especies arbóreas en selvas húmedas. Bol. Soc. Bot. Méx. 54:179–224. [ Links ]
Medina A. M., E., y J. Ángulo M. 1990. Atlas Climático del Municipio del Cofre de Perote (Estado de Veracruz). Serie Estudios Climáticos No. 1. Instituto de Ecología A. C. Xalapa, Ver. México. 48 p. [ Links ]
Mori, A., and H. Takeda. 2002. Light–related competitive effects of overstory trees on the understory conifer saplings in a subalpine forest. J. For. Res. 8:163–168. [ Links ]
Narakawa, Y., and I.S. Yamamoto. 2001. Gap formation, microsite variation and the conifer seedling occurrence in a subalpine old–growth forest, Central Japan. Ecol. Res. 16:617–625. [ Links ]
Narave, H. 1985. La vegetación del Cofre de Perote, Ver., México. Biótica 10:35–63. [ Links ]
Ott, R. A., and G. P. Juday. 2002. Canopy gaps characteristics and their implications for management in the temperate rainforests of southeast Alaska. For. Ecol. Manage. 159:271–291. [ Links ]
Pham, A. T., L. D. E. Grandpré, and S. Gauthier. 2004. Gap dynamics and replacement patterns in gaps of the northeastern boreal forest of Quebec. Can. J. For. Res. 34: 353–364. [ Links ]
Poulson, T. L., and W. J. Platt. 1989. Gap light regimes influence canopy tree diversity. Ecology. 70:553–555. [ Links ]
Rasmussen, H. N., S. Soerensen, and L. Andersen. 2003. Bud set in Abies nordmanniana Spach., influenced by bud and branch manipulations. Tree 17:510–514. [ Links ]
Romme, W. H., and D. H. Knight. 1981. Fire frequency and subalpine forest succession along a topographic gradient in Wyoming. Ecology 62:319–326. [ Links ]
Runkle, J. R. 1981. Gap regeneration in some old–growth forests of the eastern United States. Ecology. 62:1041–1051. [ Links ]
Runkle, J.R. 1989. Synchrony of regeneration, gaps and latitudinal differences in tree species diversity. Ecology. 70:546–547. [ Links ]
Rzedowski, J. 1978. La Vegetación de México. Edit. LIMUSA, México. 432 p. [ Links ]
Sánchez–Velásquez, L. R. 1986. Estudio de la sucesión forestal en la Sierra de Juárez, Oaxaca, México, después de un incendio forestal superficial. Biotica 11:219–232. [ Links ]
Sánchez–Velásquez, L. R., y E. García–Moya. 1993. Sucesión forestal en la Sierra de Manantlán Jal., México: bosque mesófilo de montaña y bosque de Pinus. Agrociencia 3:7–26. [ Links ]
Schnitzer, S. A., and W. P. Carson. 2001. Tree fall gaps and the maintenance of species diversity in a tropical forest. Ecology 82:913–919. [ Links ]
Schupp, E. W., H. F. Howe, and C. K. Augspurger. 1989. Arrival and survival in tropical treefall gaps. Ecology 70:562–564. [ Links ]
Silvertown, J., and M. Doss. 1999. Evolution of life history in balsam (Abies balsamea) in subalpine forests. Proc. R. Soc. Lond. B. 266:729–733. [ Links ]
Spies, T. A., and J. F. Franklin. 1989. Gap characteristics and vegetation response in coniferous forest of the Pacific Northwest. Ecology 70:543–545. [ Links ]
Snook, L. 1993. Conservation of the Monarch butterfly reserves in México: Focus on the forest. In: Malcolm, S., and M. Zalucki (eds). Biology and Conservation of the Monarch Butterfly. Science Series No. 38, Natural History Museum of Los Angeles County, pp: 362–375. [ Links ]
Spurr, H., y B.V. Barnes. 1982. Ecología Forestal. 3ª Ed. AGT Editor, SA. México. 690 p. [ Links ]
Sugita, H., and M. Tani. 2001. Differences in microhabitat related regeneration patterns between two subalpine conifers, Tsuga diversifolia ana Abies mariesii, on Mount Hayachine, northern Honshu, Japan. Ecol. Res. 16:423–433. [ Links ]
White, P. S. 1979. Pattern process, and natural disturbance in vegetation. Bot. Rev. 45:229–299. [ Links ]
Whitmore, T. C. 1984. Gap size and species richness in tropical rain forest. Biotropica 16:239. [ Links ]
Whitmore, T. C. 1989. Canopy gaps and the two major groups of forest trees. Ecology 70: 536–538. [ Links ]
Worrall, J. J., T. D. Lee, and T. C. Harrington. 2005. Forest dynamics and agents that initiate and expand canopy gaps in Picea –Abies forest of Crawford Notch, New Hampshire, USA. J. Ecol. 93:178–190. [ Links ]

