










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkPolibotánica
versão impressa ISSN 1405-2768
Polibotánica no.31 México Mar. 2011
Plasticity of biomass component allocation patterns in semiarid tamaulipan thornscrub and dry temperate pine species of northeastern Mexico
Componentes de la biomasa en el matorral espinoso tamaulipeco y los bosques templados secos de pino del nordeste de México
José Návar
Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional-IPN
Unidad Durango. Sigma s/n Fracc. 20 de Noviembre II. Durango, Dgo. 34220. México.
& Fax 618-8142091
Correspondencia:
jnavar@ipn.mx
Recibido: 29 abril 2010.
Aceptado: 9 diciembre 2010.
Abstract
The question of how plant communities respond to variation in the availability of resources in the environment was answered by testing the hypotheses of true adjustments and ontogenetic drift in biomass allocation patterns. The slopes of allometric equations that estimate biomass components as a function of diameter at breast height, D, root profiles and the ratio of belowground to aboveground biomass, BGB:AGB, vs. basal diameter, Db, supported these hypotheses in Mexico's northeast Tamaulipan thornscrub and dry temperate pine forests. Covariance analysis showed that plant communities allocate biomass components differently; Tamaulipan thornscrub species have larger root biomass allocation per unit Db than dry temperate pine forests in early stages of development. Tamaulipan thornscrub roots are more numerous, smaller and thinner, and differences in taper were noted for both lateral and tap root systems. This report concludes that environmental factors (more abundant soil nutrients, but less water availability in Tamaulipan thornscrub communities) play an important role in explaining higher BGB: ABG ratios, but differences can be eventually obscured by the developmental stage of these plant communities.
Key words: bole, branch and foliage, coarse roots, root: shoot ratio, root tapering.
Resumen
La pregunta de cómo dos comunidades de plantas responden a la variación en la disponibilidad de recursos en el ambiente fue contestada al probar la hipótesis del ajuste verdadero y aparente en la asignación de componentes de biomasa. Las pendientes de las ecuaciones alométricas que estiman los componentes de biomasa en función del diámetro a la altura del pecho, D, los perfiles radiculares y el índice de la biomasa subterránea a la biomasa aérea, BGB:AGB, vs. diámetro basal, Db, apoyaron estas hipótesis en el matorral espinoso tamaulipeco y los bosques templados secos de pino del noreste de México. Los resultados mostraron a través del análisis de covarianza que las comunidades de plantas asignan a los componentes de biomasa de una forma diferente; el matorral espinoso tamaulipeco distribuye mayores cantidades de biomasa radicular por unidad de Db que los pinos de bosques templados secos pero sólo en las etapas tempranas de la sucesión. Las raíces del matorral son menores en tamaño y en espesor y se observaron diferencias en el ahusamiento de raíces laterales y de anclaje. Este reporte concluye explicando que la diferencia en la partición de biomasa se atribuye a los factores ambientales (mayor fertilidad pero menor disponibilidad de agua en el matorral) pero las diferencias pueden eventualmente ser oscurecidas por el estadio sucesional de estas comunidades.
Palabras clave: fuste, ramas y follaje, raíces gruesas, índice de biomasa radicular a biomasa aérea, ahusamiento de raíces.
INTRODUCTION
Optimal partitioning of biomass among various plant organs helps to effectively capture nutrients, light, water, and carbon dioxide to maximize plant growth rate (Hilbert, 1990; Dewar, 1993). Factors that limit the acquisition of below-ground resources relative to light and CO2, should have increased root biomass in lieu of boles and foliage. The theory behind optimal partitioning is generally accepted and several studies report adjustments in biomass allocation patterns consistent with this theory (Reynolds and D'Antonio, 1996).
The ratio of below (BGB) to above (AGB) ground biomass components; BGB:AGB is recognized as the single most important expression of optimal partitioning (Evans, 1972; Coleman et al., 1994). Several good datasets of BGB:AGB ratios for large trees exist (Barton and Montagu, 2006), although the current debate centers on the mechanisms underlying the observed partitioning responses (Chu et al., 1992; Dewar, 1993; Luo et al., 1994). These ratios are known to vary with a number of environmental factors. However, the ratio also changes with plant size or age, obscuring the true environmental effect on optimal biomass partitioning (Rice and Bazzaz, 1989).
True and apparent biomass allocation patterns are names coined to separate environmental (true) from age or size (apparent) plasticity (Coleman et al., 1994). Plasticity is the amount by which; i.e. the ratio BGB: AGB responds to environmental variation that can or cannot be coupled with size or age. Ontogenetic drift (apparent plasticity) in biomass allocation patterns is generally associated with growth rates. To complicate the understanding on plasticity, in some communities the BGB:AGB ratios can go in the opposite direction when plants are compared at a common age instead of a common size (Evans, 1972); or when ontogenetic drift increases or decreases the degree of phenotypic plasticity observed over time or size (Ackerly et al., 1992; Poorter and Pothmann, 1992; Gedroc et al., 1996; Mc-Connaughay and Coleman, 1998; 1989).
Information on optimal plasticity and whether it changes with the environment is useful for understanding, i.e., the effect of climate cycles or potential climate shifts on biomass compartment allocation patterns and how these episodes may eventually control plant productivity. However, the hypothesis of true or apparent plasticity has not been supported by exploring other relationships; i.e., shifts in root profiles for both lateral and taproot systems. There is a chronic lack of information on root tapering of most tree species. In the past, conventional taper functions have been applied to estimate the volume of available timber, of different sizes, and to examine merchantable timber. These studies help to determine the possible end-uses for a single log; i.e., sawn wood, pulp, and plywood (Cao et al., 1980; Parresol and Thomas, 1996). These techniques describe the root profile, which would be useful to estimate length, volume, and root biomass that aid to understand the hypothesis of true or apparent root plasticity. That is, whether root taper differs between plant communities and if variation is size-dependent as well.
There is also a lack of information on how biomass allocation patterns adjusts in semi arid shrubs compared to dry temperate tree species of Mexico's northeast plant communities. In order to understand whether plasticity is operating in these plant communities, I tested the ontogenetic drift hypothesis that there is no relationship between the BGB:ABG ratio to Db for each of both plant communities. If Ho is rejected, then a second true plasticity set hypothesis would be that the slopes of these relationships (BGB:AGB vs Db) do not interact. Other related single predictions that help to support if true and apparent plasticity patterns are driving productivity changes were: a) the slopes of the biomass component i vs D are similar between both plant communities and b) root tapering is similar between these plant communities. With the aim of understanding better plasticity patterns of both plant communities, I establish the following objectives: a) to measure and allometrically model biomass components (foliage and branches, roots and boles); b) to measure and model root profiles of lateral and taproot systems; c) to carry out statistical analyses to test for differences in treatments and slopes of allometric relationships.
MATERIAL AND METHODS
The study area. Tamaulipan thornscrub forests are well distributed in the plains along the Northern Gulf of Mexico. The species sampled in this plant community were Condalia hookeri M.C. Johnst. and Diospyros texana Scheele. These species represent variations of this plant community. Dry temperate pine forests are well distributed in the Sierra Madre Oriental mountain range of Northern Mexico. Pinus cembroides Zucc, Pinuspinceana Gord. and Pinus pseudostrobus Lindl. are species representative of the environments within this mountain range. Therefore two sites based on differences in environmental variables (i.e, evapoptranspiration and soil fertility) were sampled to test set hypotheses: one in the Tamaulipan thornscrub forest and one in reforestation sites of the Sierra Madre Oriental mountain range of Northeastern Mexico (Fig. 1).
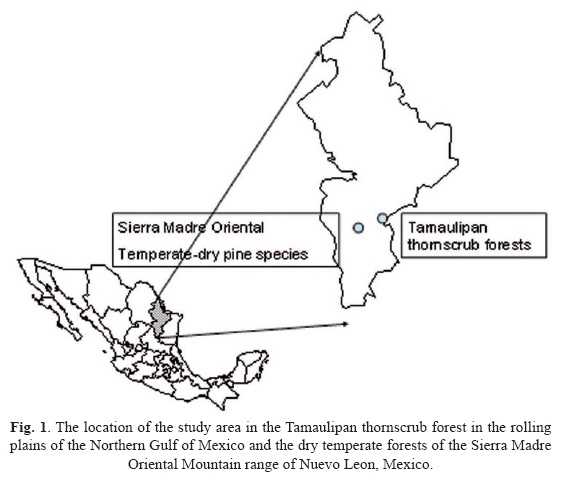
The plains of the Northern Gulf of Mexico occupy an approximate area of 200 000 km2, in Mexico and Southern USA. In Mexico, this ecosystem is in an important transitional state because of constant changes in land use (Treviño et al., 1996; Návar-Cháidez, 2008). This region present semi arid, sub tropical climate with a mean annual rainfall of 750 mm and standard deviation of 250 mm. Rainfall is distributed bimodally, during May-June and September-October, with an intermediate summer drought period. In general, annual rainfall occurs in 80 rainfall events, of which only on the average 4 of these surpass 40 mm in rainfall depth (Návar et al., 1999). The mean annual temperature is 22.3°C and annual pan evaporation is greater than 2 000 mm (Návar et al., 1994). Deep Vertisols dominate the landscapes of the piedmont and plains of Northeastern Mexico, underlined by limestones of Cretacic origin. Vertisols are characteristic of the rolling plains; although there are various soil textures, the main component of the soil is clay (Woerner, 1991).
Dry temperate pine forests were studied within the Universidad Autonoma de Nuevo Leon, UANL, property, which is located near the town of Santa Rosa in the municipality of Iturbide, Nuevo Leon, Mexico, 40 km to the southwest of the city of Linares between the geographical coordinates 24°43' N and 99°52' W. The property is at an altitude that oscillates from 1 200 to 1 900 m.s.n.m., within the main Mountain range of the Eastern Sierra Madre (which extends from Mexico's northeast to the central plateau). The annual average precipitation of the area is 600 mm, with a standard deviation of 240 mm. Rainfall is distributed bimodally, during May-June and September-October, with an intermediate summer drought period. Winters are dry and cold-temperate, with occasional storm waves produced by the "Nortes", cold air masses which originate in the Arctic. The annual average temperature is approximately 17°C, with maximum daily temperatures as high as 30°C in the summer and below 0°C in the winter. Annual pan evaporation is approximately 800 mm (Návar et al., 1994). The geological landscape of the area contains lutitas and limestone of the Upper Cretácic origin, along with recent sedimentary deposits. Outcrops of the Jurassic and the Lower Cretacic period are located along the slopes of the landscape. The topography is rough, with typical slopes ranging from 30-70%, with isolated mesetas that have gentle slope. The soil is thin and the depth variation is affected by slope, vegetation and geology. The limestone areas contain dark clay soils and massive rocks; they are deeper in north facing slopes. The lutitas are more fragile. Several trenches within the landscape show eroded soil, with poorly developed profiles and horizons, but with a high penetrability by plant roots (Woerner, 1991).
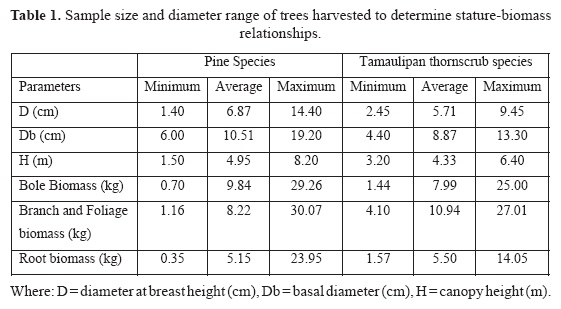
Samples. The experimental design required the selection of 40 trees; 20 in the Tamauli-pan thornscrub forests and 20 in semi arid, upland areas. The species sampled (number of trees) in the Tamaulipan thornscrub forests were Diospyros texana (12) and Condalia hookeri (8); and in the temperate forests, Pinus cembroides Zucc. (4); Pinus pinceana Gordon (5), Pinus psudostrobus Lindl. (7), Pinus brutia Tenn (2) and Cu-pressus arizonica (2). Allometric equations for pine species do not show a great deal of variation as it has been reported earlier by Návar et al. (2004) for other pine species of Durango, Mexico. Therefore the number of pine species that makes the sample is not a limiting factor for contrasting biomass components with other plant communities. Diameters at the base of the stump and at breast height were recorded using diameter tapes on standing trees. Trees were felled and separated into their main components: a) branches and foliage and b) stems. The coarse roots of each of the 40 trees, ranging in D from 1.4 to 14.4 cm, were excavated in their entirety (i.e., > 0.5 cm diameter). All biomass components were freshly weighed and samples were collected for oven-dry analysis (to a constant weight at 70°C).
Dried samples were weighed; a ratio of oven-dry to fresh weight of samples was calculated and multiplied by the total fresh weight. This estimated the total dry weight per biomass component, per tree. Parameters of measured trees are reported in Table 1.
Roots larger than 0.5 cm in basal diameter were separated and their diameter profile was measured with diameter tapes, every 5 cm from the base to the tip. The basal and breast height diameters of the trees were recorded to 0.1 cm. The top height of these trees was recorded to 0.05 m. The top height was measured directly on the main stem after trees were felled down. The total fresh weight of each component was obtained in the field using electronic balances. This weight was recorded to the nearest 1 g for material weighing < 5 kg, or to 10 g for heavier materials. Dry weights were recorded to 0.1 g accuracy.
Data analysis
Supporting allometric relationships. Allometric biomass equations were derived for each biomass component and total tree per plant community. The equations fitted the following mathematical form Bi = αDiβ ± ei where: Bi is the biomass component i; D = diameter at breast height, ei = the error, and α, β are statistical parameters to be determined by regression techniques. This equation can be linearized by taking the logarithms of Bi and D, which gives the most conventional allometric model reported in the scientific literature (Návar, 2010b): Ln(Bi) = α + β*Ln(Di) ± ei. This transformation facilitates covariance analysis to determine the effect of plant communities on the biomass components and changes in the slopes of these relationships account for size. Analysis of covariance, ancova, combines features of analysis of variance, ANOVA, and regression. It examines treatment differences adjusted for the covariate D or Db. Similarly, it determines whether there is a significant relationship between the biomass component and D after adjusting for treatment means. I define treatments as the Tamaulipan thornscrub and dry temperate pine forests. If there are significant changes in the slopes, then each plant community has different biomass components, accounting for differences in diameter. The ancova analysis was conducted with a model that consider the statistical significance of the covariate, the plant communities and the interaction covariate*plant community. The last source of variation, interaction, supports the hypothesis of true biomass allocation. Root profiles were modeled using taper functions. There is a wide range of taper functions described in the literature (Cao et al., 1980; Max-Burkhart, 1976; Parresol and Thomas, 1996; Newnham, 1990). The tapering shifts between trees of the same stand and trees of different stands, in addition to the different mathematical perceptions on the mode of tackling the mathematical description of the stem profile, have led to the abundance of taper functions (Gregoire and Schabenberger, 1996; Tassisa and Burkhart, 1998; Eerikãinen, 2001). In this research, the Newnham (1990) taper model was employed (equation [1]). This model has been consistent in describing the stem profile for seedlings and trees of Northern Mexico (Návar and Domínguez, 1997; Návar, 2010).
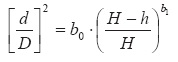
Where: d = diameter i at root length i; h = root length i, D = basal diameter of roots, H = total length of roots, b0 and b1 are statistical parameters.
Changes in the stem profiles were determined by computing statistical parameters b0 and b1 and the confidence intervals, and by graphing the root profiles of the main tree species. Significant changes in the Newnham (1990) model parameters helps to understand the hypothesis of true biomass allocation in roots. Likewise a lack of statistical significance implies that data fails to provide further information on true biomass allocation patterns. Graphs for different root basal diameters explained whether differences of root profiles for each plant community are size-dependent.
Testing the hypothesis of true and apparent biomass allocation patterns. The ratios BGB:AGB related to Db for each of the two plant communities tested the hypothesis of true (environment) and ontogenetic drift (apparent plasticity). The statistical significance of the ancova model (Ln(BGB:AGB) = Ln(Db) + Ln(Plant Communities) + Ln (Db*Plant Communities)) checked the hypothesis that allocation patterns are a function plant size (Db; ontogenetic drift), plant communities and the interaction Db*plant community. Since Db and age are well related for most tree species, apparent plasticity is tested by the statistical significance of this relationship. Statistical differences in slopes (Db*plant community) accept the hypothesis of true biomass adjustments that can be further explained by environmental variation. The plant community treatment effect tests the hypothesis that collected BGB:AGB data is similar in both plant communities.
RESULTS
Empirical supporting predictions. Power functions fit well the allometric data for all biomass components (p = 0.0001) for Tam-aulipan thornscrub species (Fig. 2).
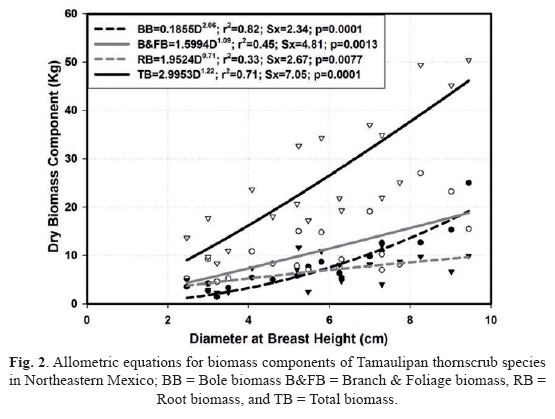
The variance accounted for in the above models is > 33%. All biomass equations, with the exception of root biomass, have an exponent larger than 1. That is, biomass increases exponentially with D, which is consistent with most allometric biomass equations. For roots, allometric equations with exponent values > 1.0 are found when the statistical analysis is conducted on each individual shrub species for this range of measured diameters. The mean (confidence interval) ratio of root biomass to total aboveground biomass is 0.30 (0.04). That is, root biomass is close to 30% of the total aboveground biomass. When considering branch and foliage biomass alone, this value increases to 66%. The ratio of below to aboveground biomass varied notoriously although statistically similar between Diospyros texana and Condalia hookeri, since these species have mean (confidence interval) values of 0.34 (0.05) and 0.23 (0.07), respectively.
The allometric equations are well fit by the power function (p = 0.0001) for all biomass components for dry temperate pine species with coefficients of determination larger than 0.60 (Fig. 3).
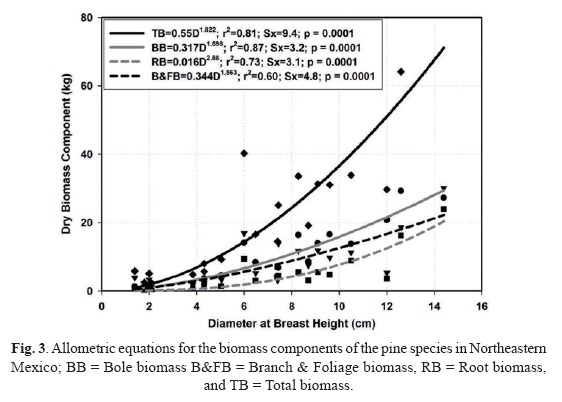
All power coefficients are larger than 1.0, consistent with most allometric biomass equations as well. The root biomass fraction is smaller than either the branch and foliage component or the bole component. Root biomass is better predicted in pine species than in Tamaulipan thornscrub species. Branch and foliage biomass components exhibit a large variation in both plant communities. This component is larger than the root component in both plant communities. The mean (confidence interval) ratio of below to aboveground biomass is 0.26 (0.07).
The ancova analysis showed that D is a significant covariate. This variable explains an important part of the biomass component variance (p = 0.0001). According to the analysis, plant communities exhibit different biomass amounts for branch and foliage (p = 0.0007), for coarse roots larger than 0.5 cm (p = 0.0001) and for total tree biomass (p = 0.0003), unlike for boles (p = 0.062). However, the ancova analysis for testing the slope variability showed that the biomass allocation per unit D value was not significantly different between plant communities for boles (p=0.54), for branches and foliage (p = 0.75) and for total tree biomass (p = 0.30); unlike for roots (p = 0.03). That is, plant communities allocate differential biomass proportions to their root component per unit value of D. Root biomass for this diameter range is larger in Tamaulipan thornscrub shrubs species than in dry temperate pine species.
Root basal diameters are normally distributed for all tree species (Table 2). The mean root basal diameter is larger for the Tamaulipan thornscrub species C. hookeri than for the rest of the species.
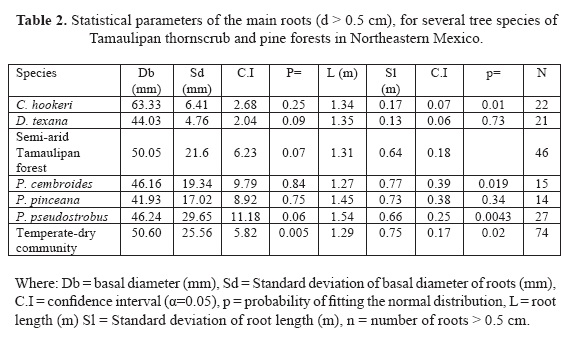
Both plant communities had similar mean root basal diameter values. On the other hand, root length is only normally distributed for D. texana and P. pinceana. However, root length was not significantly different among tree species noted in Table 2. The number of coarse roots per tree was statistically different; with 3.7 (0.25) and 2.3 (0.31) roots per tree for Tamaulipan and temperate-dry forests, respectively.
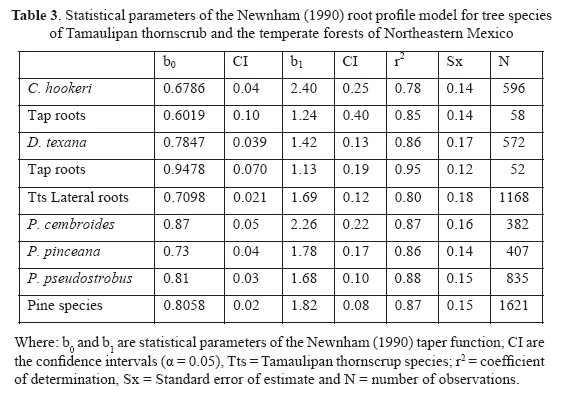
The statistical parameters of the taper models indicate that the intercept (b0) is statistically different between plant communities; however, the slopes (b1) for lateral for both plant communities and for taproot systems of both Tamaulipan thornscrub species are statistically similar (Table 3).
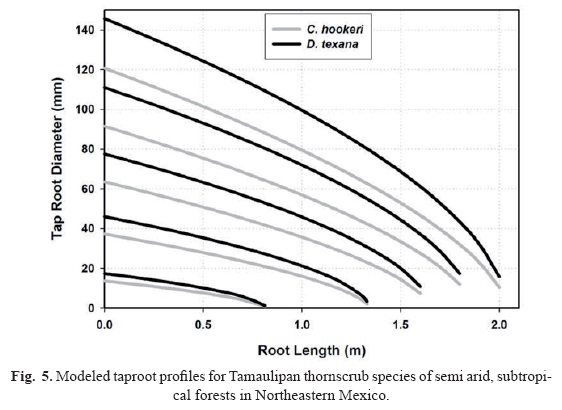
Pines did not present well differentiated tap root systems, thus, there are no root profile equations for these species. Profile equations predict that the roots of Tamaulipan thornscrub forests are less thick and shorter than the roots of the pine species (Fig. 4). The difference in basal diameter, length and tapering between plant communities increases as the root size augments stressing potential adaptations to environment and plant community characteristics; i.e., the differential soil area exploited by each plant community (Fig. 4).
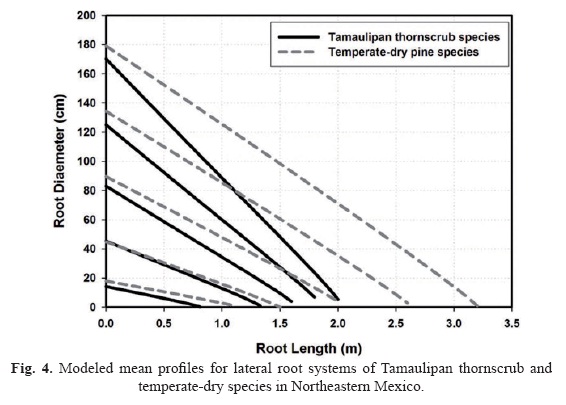
Unlike the roots of the Tamaulipan thornscrub species, the root profiles are quite similar between pine species. Roots of C. hookeri are thicker, shorter, and are less tapered than the roots of D. texana. Tap roots differ in tapering between Tamaulipan thornscrub species (Fig. 5). They are longer and thicker at the base in D. texana than in C. hookeri. In contrast to the roots of C. hookeri, the diameter differences at the root base are on the order of 25% and above 50% at the estimated tip.
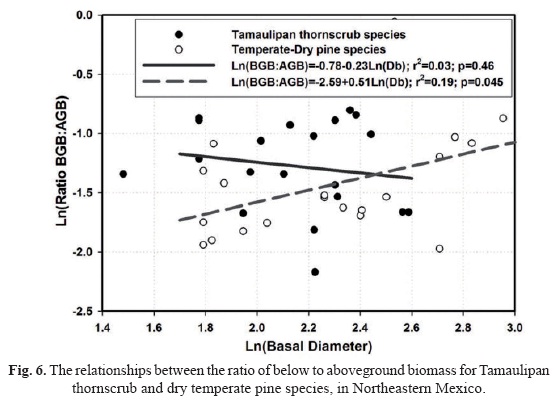
Testing the hypothesis of true and apparent plasticity. The ancova analysis was statistically significant but only at p = 0.10 (F = 2.25; D.F = 39). Shrub size plays an important role in defining the BGB:AGB relationship in pine species (p = 0.042) unlike in Tamaulipan thornscrub forests (p = 0.46) stressing the need for collecting further biomass data in the latter forest community to fully test the ontogenetic drift hypothesis. Plant communities allocate biomass components in a different manner (p = 0.048) supporting the hypothesis of true biomass allocation. Although the ratio is statistically similar in Tamaulipan thornscrub forests (0.30 ± 0.04) than in temperate-dry pine species (0.26 ± 0.07), the interaction (Db*plant community) was significantly different at p = 0.07 indicating that slopes move in the opposite direction. That is, true can overlap apparent biomass allocation patterns at an intermediate developmental stage of these plant communities. In Tamau-lipan thornscrub forests, although the ratio Ln(BGB:AGB) decreases linearly, with a slope of -0.023 per unit of Ln(D), the trend is not statistically significant (p = 0.14) (Fig. 6). On the other hand in dry, temperate pine forests, the log transformation of the ratio increases linearly with diameter, with a slope of 0.51 per unit of ln(D). That is, the ratio is larger in Tamaulipan thornscrub species in the early but it will eventually be smaller in the latter stages of succession in contrast to the ratio of pine species (Fig. 6).
DISCUSSION
The BGB:AGB ratios measured in this study are similar to ratios reported for other plant communities. For worldwide plant communities, Cairns et al. (1997) reported a mean ratio of 0.27. Castellanos et al. (1991) reported a ratio of close to 0.30 for a tropical dry forest in Western Mexico. Kummerow et al. (2004) reported higher values, 0.60, for semiarid shrubs in California. Levy et al. (2004) measured similar mean fresh biomass ratios, of 0.36 (0.008), for coniferous tree species in Great Britain. The ratios for the dry temperate pine species found in this study (0.20) are a bit higher than other measured figures in other plant communities. Návar (2009) recorded estimates of belowground (roots with basal diameter larger than 0.5 cm) to total aboveground ratios and branch and foliage (ABR), to be on the average (± confidence intervals) of 0.10 (± 0.004). The ratio showed a tendency to augment with diameter at breast height. Ratio values of 0.05-0.083 were reported for tropical trees with mean D of 30 cm in Costa Rica (Hertel et al., 2003). The BGB:AGB ratio is dynamic in time and space. Návar (2009) noted that it had a tendency to augment with diameter at breast height for temperate tree species of Durango and Chihuahua, Mexico. Leuschner et al. (2007) observed that the mean root:shoot ratios vary with elevation gradients, from 0.04 at 1050 masl to 0.43 in 3060 masl, in tropical forests of Ecuador.
Allometric analysis of aboveground and belowground biomass, as a function of D, showed that the relationship was conserved across biomass components and plant communities. In contrast, the relationship between D and belowground biomass was significantly different between plant communities. This is commensurate with the large differences in BGB:AGB ratio. Therefore, the true plasticity in root vs shoot allocation between plant communities, as evidenced by unequal slopes in the root allometric plot, is a major finding of this research in native semi arid forests of northeastern Mexico. That is, Tamaulipan thornscrub species allocate less biomass to roots per D unit than pine forests. One plausible explanation is that the Tamaulipan thornscrub species have already attained a sufficiently large root biomass, in contrast to pine species (evident in the ancova analysis). Therefore, they do not require allocating further root biomass to maintain a functioning level of aboveground biomass. Indeed, the ratio of below to aboveground biomass appears to be smaller in pine species (0.26 ± 0.07) than in the Tamaulipan thornscrub community (0.30 ± 0.04). That is, at this intermediate stage of development, D. texana and C. hookeri preferentially allocate shoot biomass. In fact, the ratio is higher in smaller plants and declines with increasing D values.
Competition for belowground resources in the early stages of development is a likely mechanism of root: shoot imbalance. Therefore, Tamaulipan species must allocate biomass preferentially to the root in these early stages of succession as it is shown in this study. Access to soil moisture is of paramount importance to plant survival for D. texana and C. hookeri. In order to exploit higher soil area or volume, plants must allocate biomass preferentially to the root systems to quickly colonize soils. This idea is supported by the information on root dimensions (basal diameter and length root mass, volume tapering and number of roots) for both plant communities. For Tamaulipan thornscrub species, with less tapered, shorter and less thick roots with similar mass, the number of roots must be larger (3.7 ± 0.25) than in temperate-dry pine species (2.3 ± 0.31). In a quite dense plant community, Manzano and Návar (2000) and Návar et al. (2002) reported on average between 5 00020 000 shrubs per ha. De Los Ríos-Carrasco and Návar (2010) reported that root biomass could have a mean biomass and volume of 13.11 Mg ha-1 (1.42) and 15.76 m3 ha-1 (1.70), respectively; twice as much as pine reforestations. In order to have access to water and nutrient, shrubs must exert a tight control on the belowground, soil space. Hence, it is hypothesized that competition for water and nutrients must be stronger than competition for sun light because shrubs exhibit typical small leaf characteristics and conspicuous branch adaptations to this environment. During excavations, I observed little or no root overlap between shrubs. Ma-hall and Callaway (1998) conducted studies to reveal how root competition takes place in semi arid plant species. Ambrosia root systems appear to be capable of detecting and avoiding other Ambrosia root systems, whereas Larrea roots inhibit Larrea and Ambrosia roots in their vicinity.
The ontogenetic drift hypothesis could be adequately tested for temperate-dry pine species but the ratio (BGB:AGB) and Db showed a weak association for Tamaulipan thornscrub species. That is, there was not sufficient data to attain the statistical significance required for this interpretation. A likely explanation is that the BGB:AGB inter-specific variation is large in Tamauli-pan thornscrub forests. However, the most remarkable result of this experiment is the opposite trend in the BGB:AGB ratio with Db, for these plant communities. Therefore, there are indications that the ratios may: a) differ even more with increasing diameter, since they follow different tendencies and may eventually shift patterns in the latter developmental stages and b) overlap at some point in growth, which may obscure at some point the true adjustments in biomass allocation between these plant communities. Other exogenous variables must eventually describe better the ratio since Deans et al. (1996) noted that the ratio is positively and linearly related to the combined variable (D2H), for tropical trees of Cameroon in Africa.
In most pines, competition for the sociological position is a matter of life and death. The species tested in this study are shade intolerant and therefore they should allocate biomass preferentially to the shoot. Once they attain a dominant position, biomass allocation to roots becomes important in maintaining functioning aboveground components. Henceforth, the ratio must increase with Db. This was statistically supported (as a trend) in the present study. Therefore, competition for light shapes the ratio BGB: AGB in temperate-dry pine communities. However, this is a contrast to measurements conducted on white pine (Pinus strobus) forests in Southern Ontario (Peich and Altaf, 2007). These authors noted that the root to shoot biomass ratio decreased from 0.32, in the 2-year-old stand, to 0.24, 0.16, and 0.22 in the 15-, 30-, and 65-year-old stands, respectively. However, notable environmental differences can be described between Ontario and northern Mexico's sites.
Environmental factors such as site potential productivity (reported as soil N content) and soil water balance (reported as a ratio of annual rainfall to pan evaporation) can play an important role in modifying root biomass allocation at these intermediate (secondary) stages of tree development. Comparisons of BGB:AGB ratios, within a species growing across water and nutrient gradients, are partially consistent with predictions based on optimal partitioning theory; allocation to roots decreased under better water balance but not for high nutrient conditions in case studies. These findings have been partially confirmed through ontogenetically controlled experiments by McConnaughay and Coleman (1999). In our plant communities, the BGB: AGB ratios appear to be a function of the water balance (and/or the species composition); higher ratios were measured in drier areas and smaller ratios were measured in wetter landscapes. In general, in the area of Linares, the mean (standard deviation) annual rainfall is 750 mm (250) with an annual pan evaporation of ~2 000 mm. The climatic station of Santa Rosa, Iturbide, N.L., close to the pine forest plantations, records a mean (standard deviation) annual rainfall of 640 mm (240) and annual pan evaporation of 800 mm. The water balance is less negative in areas with dry temperate pine forests than in Tamaulipan thornscrub plant communities, since the annual water balance ratio (rainfall/pan evaporation) is 0.80 and 0.38, respectively.
This research is consistent with findings in other semi arid landscapes. Barton and Montagu (2006) observed that the BGB: AGB ratio was strongly influenced by irrigation (0.68 controls, 0.34 irrigated) but not by spacing, in eucalyptus plantations in NSW, Australia. In contrast to aboveground biomass growth, the root growth of Acacia saligna shrubs, in the USA, was dramatically reduced by irrigation (Zegada-Lizarazu et al., 2007). However, other authors found opposite results or no influence at all of water balance in root:shoot ratio. Woods et al. (2007) measured taproot elongation and noted that it increased with frequency with which plants were watered in seedlings of the Chihuahuan Desert species Prosopis velutina and Acacia greggi. On the other hand, McConnaughay and Coleman (1999) observed that this ratio was not affected by the availability of water, in seedlings of the species studied (A. theophrasti, C. album, P. pensylvanicum).
Nitrogen content in Vertisols of Tamaulipan thornscrub forests is on the order of 11% (Woerner, 1991) while nitrogen content of Cambisols of the Sierra Madre Oriental mountain range is on the order of 4% (Woerner, 1991). That is the BGB:AGB ratio is higher in soils with high nitrogen content and it is in contrast to most studies in the topic. There remains the question whether soil fertility controls the ratio within each plant community, since the scientific literature consistently reports reduced ratios with increased soil fertility indicators. Pavelka and Lindquist (2002); Caolin et al. (1980) and Lee et al. (2007) noted the ratio decreased for corn and vel-vetleaf in Nebraska, in Dactylis glomerata plants in France; and in the fine roots of P. menziesii and T. heterophylla, in the Pacific Northwest, respectively. In a survey which included 77 studies representing 206 cases and 129 species, Reynolds and D'Antonio (1996) found that root weight ratio decreased with increased nitrogen availability (in the majority of cases examined). This response was most consistent when plants were grown individually or under intra-specific competition (versus inter-specific competition). One potential justification for this contrasting trend must be in other differential environmental indicators where the water balance must not be ruled out.
These observations suggest that water could be more limiting than nutrients in plant communities in the Northeast of Mexico. Indeed, the presence of taproots in Tamau-lipan thornscrub species, in contrast to dry pine species, suggests that shrubs employ a rooting strategy attuned to local climatic conditions. In this plant community, the lateral roots tended to distribute close to the soil surface for approximately 80 to 90% of its length and then bend downwards into soil depth. D. texana has thicker and longer taproot systems than C. hookeri. This could be partially attributed to higher soil moisture content at the base of the stem, caused by stemflow, since this species funnels on the average between 3-6% of total rainfall into the stem (Návar and Bryan 1990; Návar, 2010a). On the other hand, C. hookeri funnels less than 2% of the total rainfall as stemflow (Návar et al., 1999; Návar, 2010a). Stemflow is a source of rainwater that can infiltrate deep into the mineral soil after most rainfall events, creating islands of water (Návar and Bryan, 1990; Návar, 1993) and fertility (Návar et al., 2009). Therefore, roots must tap into this source of moisture for coping with drought episodes.
Both Tamaulipan thornscrub species are especially tolerant of droughts; however, they must have other strategies than the tap root system for coping with dry conditions. D. texana has shallow lateral roots and, because it is a deciduous plant species, it can shed leaves in dry spells. C. hookeri is an evergreen plant species, with most roots distributed at intermediate soil depths. Therefore, it exploits different soil compartments and at differential soil water potentials than D. texana. Indeed, soil water potential at 50 cm and at 100 cm is better correlated to predawn plant water potentials, for D. texana and C. hookeri, respectively (Zou et al., 2005). Increasing the surface soil moisture by irrigation produced more leaf biomass in C. hookeri but not in D. texana seedlings (Nelson et al., 2002) stressing the potential difference in biomass allocation patterns between Tamaulipan thornscrub species and the lack of a clear association between the ratio BGB:AGB and Db for these species. Very low plant water potentials (-4.0 MPa) can be observed during heavy dry spells in D. texana, in contrast to other Tamaulipan thornscrub species (González-Rodríguez and Cantú-Silva, 2001), suggesting physiological mechanisms of drought tolerance in this species in addition to its deciduous behavior.
Pine roots tend to spread laterally for less than 50% of their length before bending vertically into the soil. In several trees, roots tend to insert into the soil diagonally. This is advantageous for exploiting the nutrients and moisture in the soil. Therefore, the results show that although all surveyed trees of both plant communities recorded a shallow root system, only the semi-arid, subtropical tree species of Tamaulipan thornscrub forest, in the drier section of the landscape, exhibited taproot systems. These findings are consistent with observations conducted in semi-arid shrubs in Namibia (Hipondoka and Versfeld, 2005).
CONCLUSIONS
In this report, I tested the hypothesis that adjustments in biomass allocation patterns occur as a function of plant size in semi arid and dry temperate forest species in Northeastern Mexico. True plasticity between plant communities was a major observation of this research that was evidenced by unequal slopes in the BGB:AGB vs Db allometric plots. The ontogenetic drift hypothesis could be partially tested because there was a lack of sufficient data in Tamaulipan thornscrub forests. It can be projected that adjustments in the allocation patterns may not be observed when comparing BGB: AGB ratios solely as a function of plant size because the stage of development of these plant communities may obscure true biomass allocation patterns. Commensurate with the optimal partitioning theory, empirical models suggests larger BGB:AGB biomass ratios are expected in Tamaulipan thornscrub than in temperate pine species in the early and vice versa in the later stages of succession. These findings were associated with drier climatic conditions in the plains of the Northern Gulf of Mexico, in contrast to the dry and temperate landscapes of the Sierra Madre Oriental mountain range.
ACKNOWLEDGMENTS
The author of this report wishes to thank M.C. Eduardo de los Ríos-Carrasco for helping in the field data collection. I am also grateful to anonymous reviewers by improving the readability and technical content of the manuscript.
LITERATURE CITED
Ackerly, D.D., Coleman, J.S., Morse, S.R. and Bazzaz, F.A., 1992. "CO2 and temperature effects on leaf area production in two annual plant species". Ecology, 73: 1260-1269. [ Links ]
Barton, C.V.M. and Montagu, K.D., 2006. "Effect of spacing and water availability on root:shoot ratio in Eucaliptus camaldulensis". Forest Ecology and Management, 221: 5262. [ Links ]
Cairns, M.A., Brown, S., Helmer, E.H. and Baumgardner, G.A., 1997. "Root bio-mass allocation in the world's upland forests". Oecologia, 111: 1-11. [ Links ]
Cao, Q., Burkhart, H. and Max, T., 1980. "Evaluations of two methods for cubic volume prediction of loblolly pine to any merchantable limit". Forest Science, 26(1): 71-80. [ Links ]
Caolin, M., Khodre, A.E. and Atry, M., 1980. "Effect of nitrate concentration on the root:shoot ratio in Dactylis glomerata L. and on the kinetics of growth in the vegetative phase". Annals of Botany, 46: 165-173. [ Links ]
Castellanos, J., Maass, M. and Kummerow, J., 1991. "Root biomass of a dry deciduous tropical forest in Mexico". Plant and Soil, 131: 225-228. [ Links ]
Coleman, J.S., McConnaughay, K.D.M. and Ackerly, D.D., 1994. "Interpreting phenotypic variation in plants". Trends in Ecology and Evolution, 9: 187-191. [ Links ]
Chu, C.C., Coleman, J.S., and Mooney, H.A., 1992. "Examining the controls on the partitioning of biomass between roots and shoots: effects of elevated levels of CO2 on growth and resource use of California coastal wild radish". Oecologia, 89: 580-587. [ Links ]
Deans, J.D., Moran, J. and Grace, J., 1996. "Biomass relationships for tree species in regenerating semi-deciduous tropical moist forest in Cameroon". Forest Ecology and Management, 88: 215-225. [ Links ]
De los Ríos-Carrasco and Návar, J., 2010. "Root allometry of two subtropical plant communities of northeastern Mexico". Tropical and Subtropical Agroecosystems, 12: 123-134. [ Links ]
Dewar, R.C., 1993. "A root-shoot partitioning model based on carbon-nitrogen-water interactions and Munch phloem flow". Functional Ecology, 7: 356-368. [ Links ]
Eerikainen, K., 2001. "Stem volume models with random coefficients for Pinus kesya in Tanzania, Zambia and Zimbabwe". Canadian Journal Forest Research, 31: 879-888. [ Links ]
Evans, G.C., 1972. "The quantitative analysis of plant growth". University of California Press, California, USA. [ Links ]
Gedroc, J.J., McConnaughay, K.D.M and Coleman, J.S., 1996. "Plasticity in root/shoot partitioning: optimal, on-togenetic, or both?". Functional Ecology, 10: 44-50. [ Links ]
González-Rodríguez, H. and Cantú-Silva, I., 2001. Adaptación a la sequía de plantas arbustivas de matorral espinoso Tamaulipeco. Ciencia UANL, 4: 454-461. [ Links ]
Gregoire, T.G. and Schabenberger, O., 1996. "Nonlinear mixed-effects modeling of cumulative bole volume with spatially correlated within-tree data". J. Agric. Biol. Environ. Stat., 1:107-119. [ Links ]
Hertel, D., Leuschner, C. and Holscher, D., 2003. "Size and structure of fine root systems in old-growth and secondary tropical montane forests (Costa Rica)". Biotropica, 35: 143-153. [ Links ]
Hilbert, D.W., 1990. "Optimization of plant root:shoot ratios and internal nitrogen concentrations". Annals of Botany, 66: 91-99. [ Links ]
Hipondoka, M.H.T. and Versfeld, W.D., 2005. "Root system of Terminalia sericea shrubs across rainfall gradient in a semi-arid environment of Etosha National Park, Namibia". Ecological Indicators, 6(3): 1-10. [ Links ]
Kummerow, J., Krause, D. and Jow, W., 2004. "Root systems of chaparral shrubs". Oecologia, 29: 163-177. [ Links ]
Lee, H.E., Tingey, D.T., Beedlow, P.A., Johnson, M.G. and Burdick, C.A., 2007. "Relating fine root biomass to soil and climate conditions in the Pacific Northwest". Journal of Ecology and Management, 242: 195-208. [ Links ]
Leuschner, C., Moser, G., Bertsch, G., Roderstein, M. and Hertel, D., 2007. "Large altitudinal increase in tree root/shoot ratio in tropical mountain forests of Ecuador". Basic and Applied Ecology, 8: 219-230. [ Links ]
Levy, P.E., Hale, S.E. and Nicoll, B.C., 2004. "Biomass expansion factors and root:shoot ratios for coniferous tree species in Great Britain". Forestry, 77: 421-430. [ Links ]
Luo, Y., Field, C.B. and Mooney, H.A., 1994. "Predicting responses of photosynthesis and root fraction to elevated [[CO2]a]: interactions among carbon, nitrogen, and growth". Plant, Cell and Environment, 17: 1195-1204. [ Links ]
McConnaughay, K.D.M., and Coleman, J.S., 1998. "Can plants track changes in nutrient availability via changes in biomass partitioning?". Plant and Soil, 202: 201-209. [ Links ]
MaConaghay, K.D.M. and Coleman, J.S., 1999. "Biomass allocation in plants: Ontogeny or optimality? A test along three resource gradients". Ecology, 80: 2581-2593. [ Links ]
Mahal, B.E. and Callaway, R.M., 1998. "Root communication among desert shrubs". Annals of Botany, 81: 213233. [ Links ]
Manzano-Camarillo, M. and Návar, J., 2000. "Desertification processes associated to overgrazing practices in Tamaulipan thornscrub of Linares, N.L., Mexico". Journal of Arid Environments, 44: 1-17. [ Links ]
Max, T. and Burkhart, H., 1976. "Segmented polynomial regression applied to taper equations". Forest Science, 22: 283-289. [ Links ]
Návar, J. and Bryan, R., 1990. "Interception loss and rainfall redistribution by three semi arid growing shrubs in northeastern Mexico". Journal of Hydrology, 115: 51-63. [ Links ]
Návar, J., 1993. "The causes of stemflow variation in three semi-arid growing shrubs in northeastern Mexico". Journal of Hydrology, 145: 175-190. [ Links ]
Návar, J., Cavazos, T. and Domínguez, P.A., 1994. "Los balances hidrológicos mensuales con tres probabilidades del estado de Nuevo León". En C. Pola S., J.A. Ramírez F., M.M. Rangel, & I. Navarro-L. (Eds) Actas Fac. Ciencias de la Tierra UANL Linares, 8: 71-82. [ Links ]
Návar, J. y Domínguez, C.P.A., 1997. "Ajuste de modelos de volumen y funciones que describen el perfil diametrico de cuatro especies de pino plantadas en el nordeste de Mexico". Investigación Agraria: Sistemas Forestales,: 6(1,2): 147-162. [ Links ]
Návar, J., Charles, F. and Jurado, E., 1999. "Spatial variations of interception loss components by Tamaulipan thornscrub in northeastern Mexico". Forest Ecology and Management, 124: 231-239. [ Links ]
Návar, J., Mendez, E. y Dale, V., 2002. "Estimating stand biomass in the Tamaulipan thornscrub of northeastern Mexico". Annals of Forest Sciences, 59: 813-821. [ Links ]
Návar, J., Mendez, E., Graciano, J., Dale, V., and Parresol, B, 2004. "Biomass equations for shrub species of Tamaulipan thornscrub of northeastern Mexico". Journal of Arid Environments, 59(4): 657-674. [ Links ]
Návar-Chaidez, J.J., 2008. "Carbon fluxes resulting from land-use changes in the Tamaulipan thornscrub of northeastern Mexico". Carbon Balance and Management, 2008 3:6 doi:10.1186/1750- 0680-3-6. [ Links ]
Návar, J., 2009. "Allometric equations for tree species and carbon stocks for forests of northwestern Mexico". Forest Ecology and Management, 31: 427-434. doi:10.1016/j.foreco.2008.09.028 [ Links ]
Návar, J., Méndez-González, J., y Gonzalez, H., 2009. "Gross precipitation and throughfall chemistry in legume species planted in Northeastern Mexico". Plant Soil, 318: 15-26 doi:10.1007/ s11104-9812-0. [ Links ]
Návar, J., 2010. "Taper function and merchantable timber for temperate forests in northern Mexico". Revista Chapingo. Serie Ciencias Forestales y del Ambiente. On Review. [ Links ]
––––––––––, 2010a. "Stemflow variation in two forest communities of northeastern Mexico: the contribution to soil moisture content and aquifer recharge". International Hydrology Congress. San Diego California, 7-11 Octubre, 2010. [ Links ]
––––––––––, 2010b. "Measurement and assessment methods of aboveground biomass: A literature Review and the challenges ahead". Biomass. Book Chapter. Sciyo. [ Links ]
Nelson, J.A., Barnes, P.W. and Archer, S., 2002. "Leaf demography and growth responses to altered resources availability in woody plants of contrasting leaf habit in a subtropical savanna". Vegetatio, 160: 193-205. [ Links ]
Newnham, R.M., 1990."Mesure du défilement de forme variable. Fôrets Canada". Institut Forestier National de Petawawa. Rapport d'information PI-X-83-F, 31 pp. [ Links ]
Parresol, B.R. and Thomas, C.C., 1996. "A simultaneous density-integral system for estimating stem profile and biomass: slash pine and willow oak". Can. J. For. Res., 26: 773-781. [ Links ]
Pavelka, K.D. and Lindquist, J.L., 2002. "The effects of nitrogen supply on root:shoot ratio in corn and velvetleaf". North Central Weed Science Society Abstracts, 57: 118. [ Links ]
Peich, M. and Altaf, A.M. 2007. "Allometry and partitioning of above-and below-ground tree biomass in an age-sequence of white pine forests". Forest Ecology and Management, 253: 68-80. [ Links ]
Poorter, H., and Pothmann, P., 1992. "Growth and carbon economy of a fast-growing and a slow-growing grass species as dependent on ontogeny". New Phytologist, 120: 159-166. [ Links ]
Reynolds, H.L. and D'Antonio, C., 1996. "The ecological significance of plasticity in root weight ratio in response to nitrogen: Opinion". Plant and Soil, 185: 75-97. [ Links ]
Rice, S.A., and Bazzaz, F.A., 1989. "Quantification of plasticity of plant traits in response to light intensity: comparing phenotypes at a common weight". Oecologia, 78: 502-507. [ Links ]
SAS., 2000. Help and documentation, SAS 9.1.3, SAS Institute Inc., Cary, NC. [ Links ]
Tasissa, G. and Burkhart, H., 1998. "An application of mixed effects analysis to modeling thinning effects on stem profile of loblolly pine". For. Ecol. Manage., 103: 87-101. [ Links ]
Treviño, E., Akca, A., Návar, J., Jimenez, J., and Aguirre, O., 1996. "Detection of land use change by satellite imagery in the municipality of Linares, Nuevo Leon, Mexico". Proceedings of the Fifth International Conference on Desert Development. Vol I. Texas Tech University, August 12-17, 1996. [ Links ]
Woerner, M.P., 1991. Métodos químicos para el análisis de suelos calizos de zonas áridas y semiáridas. Facultad de Ciencias Forestales, UANL, Linares, N.L., México. 105 pp. [ Links ]
Woods, S, Archer, S.R. and Schwinning, S., 2007. Taproot elongation in woody plant seedlings: a factor in species encroachment potential. Ecological Society of America Annual Meetings, San Jose, CA. [ Links ]
Zegada-Lizarazu, W., García-Apaza, E., Ephrath, J. and Berliner, P., 2007. "Above and below ground development of Acacia saligna shrubs grown under different irrigation frequencies in an arid environment". Plant and Soil, 297: 157-169. [ Links ]
Zou, C., Archer, S.R, Barnes, P., and Mc-Murtry, C., 2005. "Soil moisture redistribution as a mechanism of facilitation in savanna tree-shrub clusters". Oecologia, 145: 32-40. [ Links ]

