











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La familia Fagaceae comprende 8-10 géneros y más de 900 especies (Kremer et al., 2012), donde el género Quercus presenta la mayor diversidad en la familia, con cerca de 400 especies (Aldrich y Cavender, 2011). En el continente americano se estiman 250 especies de encino (Oh y Manos, 2008), de las que 161 se han registrado en México (Valencia, 2004); de esas especies, al menos 60 presentan características favorables para el aprovechamiento maderable en México (Quintanar, 2002). La mayoría de los encinos en México se encuentran en zonas montañosas con clima templado o templado subhúmedo, y más de 95% de las especies se encuentran entre los 1200 m snm y 2800 m snm (Rzedowski, 1978); la mayor riqueza de especies de Quercus se concentra en las montañas del sur (Nixon, 1993).
El predio comunal de Ixtlán de Juárez, Oaxaca, comprende 193,1 km2, de los que 64,2% son de bosque templado, y donde Quercus laurina Humb. & Bonpl. y Q. crassifolia Humb. & Bonpl. cohabitan con Arbutus xalapensis Kunth, Alnus acuminata arguta (Schltdl.) Furlow y Prunus serotina Ehrn., entre otras latifoliadas, y con Pinus spp. (Aquino et al., 2012; Ruiz-Aquino et al., 2015a). En este bosque, coexisten 416 árboles por hectárea de Q. laurina y 419 árboles por hectárea de Q. crassifolia; en su mayoría con fustes rectos, limpios y cilíndricos (Ruiz-Aquino et al., 2014).
Quercus laurina (subgénero Erytrobalanus), es un árbol de 10 m a 25 m de alto, tiene hojas deciduas, anchamente lanceoladas u oblanceoladas, de 7 cm a 10 cm de largo por 2,5 cm a 3 cm de ancho; su fruto es una bellota ovoide de 12 mm de largo (De la Paz Pérez et al., 2006). Se distribuye a lo largo de la Sierra Madre Occidental, de la Sierra Madre del Sur y por la región occidental del cinturón volcánico, entre los (2000-2440) m snm y (3065-3300) m snm (González et al., 2004; De la Paz Pérez y Dávalos, 2008). La madera de Q. laurina presenta grandes posibilidades de ser utilizada industrialmente, debido a su buen desempeño en el maquinado (Flores et al., 2007; Flores et al., 2013). Los árboles de Q. crassifolia (subgénero Erytrobalanus) alcanzan alturas de hasta 23 m, tienen hojas coriáceas, ovadas, obovadas o elípticas, de 7 cm a 17,5 cm de largo por 4,1 cm a 11,5 cm de ancho; su fruto es bianual, con cúpulas hemisféricas de 6 mm a 10 mm de largo (Valencia et al., 2002). Se distribuye en la Sierra Madre Occidental al NO de México, la Sierra Madre del Sur, y a lo largo del Eje Neovolcánico (Tovar y Oyama, 2004), de 600 m snm a 2850 m snm (Coombes, 2012). Por la respuesta de su madera en operaciones de maquinado, puede emplearse en la elaboración de molduras, lambrines, muebles y parquet (De la Paz Pérez y Dávalos, 2008; Flores et al., 2013).
Objetivos
Los objetivos del presente estudio fueron determinar las características anatómicas de la madera de Q. laurina y Q. crassifolia proveniente de Ixtlán de Juárez, e identificar las diferencias entre las dos especies. También se evaluó la influencia de la estructura anatómica en la densidad, la contracción volumétrica, y la dureza de la madera.
Materiales y métodos
Área de estudio y selección de los árboles
Se utilizaron árboles provenientes de dos rodales con dominancia de especies del género Quercus en el bosque templado de Ixtlán de Juárez, Oaxaca. El primer rodal, con dominancia de Q. laurina, a 17°21'22" LN y 96°27'42" LO, a una altitud de 2707 m snm. El segundo rodal, con dominancia de Q. crassifolia, a 17°21'08" LN y 96°27'31" LO, a una altitud 2631 m snm. Se seleccionaron tres árboles para cada especie, sanos y representativos, con un diámetro a 1,3 m sobre el nivel del piso entre 30cmy 40cm,y de 20m a 25m de altura. El derribo de los árboles y el troceo se hizo de acuerdo con Ramos y Díaz (1981), tomando para el estudio microscópico una rodaja de 5 cm de espesor a la altura de 1,30 m, y para la caracterización macroscópica, la siguiente rodaja, de 30 cm de espesor. El material restante de los árboles derribados se utilizó para cuantificar la biomasa aérea (Ruiz-Aquino et al., 2014), y para la caracterización química y energética de la madera y el carbón de los dos encinos (Ruiz-Aquino et al., 2015b). El depósito e identificación de los especímenes se realizó en el herbario de la Universidad de la Sierra Juárez, Oaxaca.
Caracterización macroscópica
La descripción macroscópica se hizo con tablillas de 1 cm x 7 cm x 15 cm, orientadas en los tres planos (De la Paz Pérez y Dávalos, 2008), utilizando la clasificaciones de Tortorelli (1956) para textura, porosidad, hilo y veteado, y de Ortega et al. (1988) para el brillo, olor y sabor; para la descripción del color, se usaron las tablas Munsell (1990).
Caracterización microscópica
De la rodaja para el estudio microscópico se cortaron cubos de 2 cm por lado (10 cubos de albura y 10 de duramen), los que se ablandaron saturándolos al vacío con agua destilada y luego calándolos en agua hirviendo por 10 min. Se cortaron secciones de 20 μm en los tres planos con un xilotomo SM2000R (Leica, Alemania), y se tiñeron con Pardo de Bismarck a 1% por 6 h. Luego se deshidrataron en una serie de alcoholes y finalmente se aclararon en xilol por 2 min; las secciones se montaron con resina sintética (Sandoval, 2005). Para obtener el material disociado, se cortaron astillas y se digirieron en la solución de Franklin (1946) a 60 °C por 24 h. Luego se enjuagaron y se tiñeron con Pardo de Bismarck, para hacer preparaciones temporales. Las mediciones se realizaron en un microscopio de luz DM 500, con cámara integrada ICC50 (Leica, Alemania). Los vasos se clasificaron en vasos con diámetro ancho (> 150 μm) y vasos con diámetro angosto (≤ 150 μm) (Chávez et al., 2010). Se midió el diámetro mayor de los vasos en el corte transversal (n = 600 por especie), y la longitud de elementos de vaso en el material disociado (n = 90 por especie); las mediciones se realizaron de acuerdo con IAWA (1989). También se realizaron 120 mediciones por especie de la longitud y ancho de las fibras, del diámetro de lumen y el espesor de pared de las fibras, del alto y ancho de los radios uniseriados, del ancho y número de series en radios multiseriados, y del número de vasos por milímetro cuadrado. Los elementos mensurables se clasificaron con base en la media de acuerdo a la clasificación de Chattaway (1932), IAWA (1939) y Tortorelli (1956). Para determinar la altura de los radios multiseriados, se midieron con un vernier 32 radios completos en la albura y en el duramen de cada especie, utilizando las tablillas de la caracterización macroscópica, sin hacer distinción del árbol al que pertenecían.
Para el estudio al microscopio electrónico de barrido, se cortaron cubos de madera de 5 mm por lado, se deshidrataron en una serie etanólica creciente, y se recubrieron con oro. Se utilizó un microscopio JSM-6390 (JEOL Ltd., Japón), operando a 15 kV.
Propiedades físicas y dureza
Con el objetivo de establecer la influencia de las estructuras anatómicas sobre dos propiedades físicas y una propiedad mecánica de la madera, se utilizaron probetas de una mezcla de albura y duramen para determinar la densidad básica (DB) (n=36 por especie), la contracción volumétrica (CV, n=36) y la dureza (n=24), siguiendo la norma ASTM D 143-94 (ASTM, 2007). Estas propiedades se clasificaron de acuerdo a Sotomayor (2005), Chávez-Doldán (2007) y Dávalos y Bárcenas (1999), en el mismo orden. El material para estas determinaciones se obtuvo de la primera troza comercial de 1,20 m de longitud de cada árbol, aserrada en cuartos, para obtener polines de 7 cm x 7 cm x 120 cm con sus tres planos típicos definidos (Najera et al., 2005).
Análisis estadístico
Para determinar diferencias entre los elementos celulares de las especies estudiadas, se realizó un análisis de varianza, seguido de un análisis de comparación de medias (Tukey, α = 0,05). Para determinar la relación de los elementos celulares con la densidad básica, la contracción volumétrica y la dureza de la madera, se calculó el coeficiente de correlación de Pearson (r). Los análisis estadísticos se realizaron usando el paquete estadístico SAS (SAS Institute, 1989).
Resultados y discusión
Caracterización macroscópica
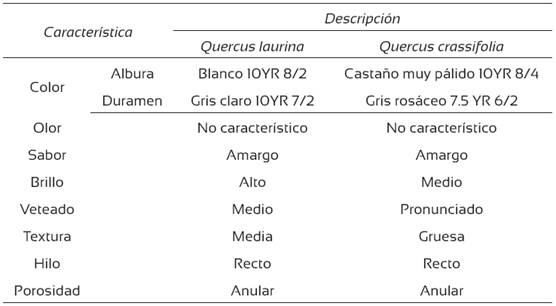
En general, el duramen fue más oscuro que la albura, aunque no se observó una diferencia muy marcada entre el color de la albura y del duramen en las dos especies. La madera de Q. laurina fue más clara que la de Q. crassifolia, tanto en albura como en duramen (Tabla 1). El sabor amargo de la madera es una característica común en las dos especies, resultado de la cantidad de extractivos presentes como taninos y polifenoles. Honorato y Hernández (1998) cuantificaron los taninos de la madera de Q. laurina y de Q. crassifolia, 1,17% y 1,52% en el mismo orden, sin diferencias estadísticas entre las dos especies. La porosidad anular de las dos especies es similar a la encontrada por De la Paz Pérez y Dávalos (2008) y tiene influencia sobre el veteado de la madera. Otras características anatómicas macroscópicas se presentan en la Tabla 1.
Caracterización microscópica
Radios
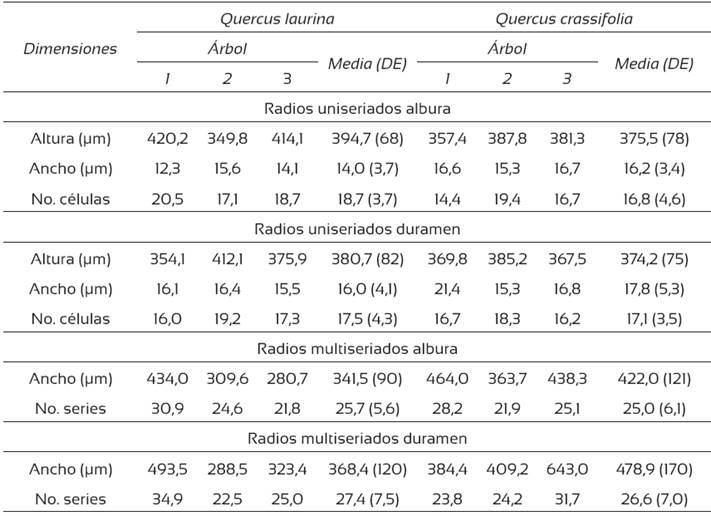
Una de las características anatómicas más notable de la madera de encino es la presencia de radios multiseriados. Las especies que aquí se describen presentan radios uniseriados, multiseriados y agregados. Las dimensiones y el número de células de los radios se consignan en la Tabla 2. En las dos especies, los radios leñosos son homogéneos (Fig. 1A, 1B), formados por células procumbentes (Fig. 1C, 1D), lo que coincide con De la Paz Pérez et al. (2006) y De la Paz Pérez y Dávalos (2008). Los radios uniseriados en la madera de albura de Q. laurina son 3,5% más altos que los de Q. crassifolia; sin embargo, no se encontraron diferencias significativas en la altura de los radios entre albura-albura, duramen-duramen y albura-duramen de ambas especies. Según la clasificación de Chattaway (1932), los radios uniseriados se clasifican como extremadamente bajos; respecto al ancho, los radios uniseriados se clasifican de extremadamente finos a muy finos (IAWA, 1939). Se encontraron diferencias estadísticas significativas entre albura y duramen de Q. laurina (p = 0,006), albura y duramen de Q. crassifolia (p = 0,046), y en albura-albura (p = 0,001) y en duramen-duramen (p = 0,038) de las dos especies. Las medias halladas en el presente estudio para Q. laurina, en los radios uniseriados (número de células) y en los radios multiseriados (número de series), fueron mayores a los encontrados para la misma especie por De la Paz Pérez y Dávalos (2008), 11 células y 21 series. Para Q. crassifolia, De la Paz Pérez et al. (2005) encontraron radios multiseriados muy anchos (387 μm, 20 series), valores menores a los que se obtuvieron en este estudio. No se encontraron diferencias significativas al comparar la altura de los radios multiseriados entre la albura-albura y el duramen-duramen de las dos especies. En Q. laurina, la altura de los radios multiseriados fue de 17,6 mm ± 3,4 mm y de 13,2 mm ± 1,5 mm en albura y duramen, respectivamente; mientras que en Q. crassifolia, la misma altura fue de 16,7 mm ± 3,1 mm y 13,4 mm ± 3,1 mm, en el mismo orden. Estas alturas son comparables con las obtenidas previamente para los radios multiseriados de Q. laurina (1,5 mm a 2,0 mm) y Q. crassifolia (0,7 mm a 1,5 mm) por De la Paz Pérez y Dávalos (2008). Por otra parte, en cada una de las dos especies, los radios multiseriados fueron significativamente más altos en la albura que en el duramen (p < 0,001 en los dos casos). Las dimensiones de los radios son de las variables anatómicas que más influencia tienen en el proceso de secado, debido a que presentan una pared celular primaria flexible, poco engrosada y constituyen puntos débiles en la madera por la ausencia de la pared secundaria (Metcalfe y Chalk, 1985). En conjunción con su densidad alta, acentúan las contracciones altas en la madera, ocasionando problemas en su industrialización. Por otra parte, los radios visibles a simple vista, son un carácter anatómico valioso de la figura de la madera de encino, apreciado en la fabricación de pisos, muebles y chapas.
Vasos y parénquima axial
Con respecto a su longitud, los elementos de vaso se clasifican como medianos en ambas especies (Chattaway, 1932). En el plano transversal, los vasos presentan poros con diámetro tangencial de moderadamente pequeños a moderadamente grandes (IAWA, 1939). En la albura de ambas especies, los poros por milímetro cuadrado son escasos y en el duramen moderadamente escasos (Chattaway, 1932). La consideración del diámetro y la cantidad de poros por milímetro cuadrado es importante en las operaciones de acabado y pegado, ya que a mayor número por milímetro cuadrado, es más difícil darle un buen acabado a la madera y la adhesión de las superficies es menor.También tiene relevancia en los procesos de impregnación de la madera, pues la presencia de vasos numerosos es deseable para facilitar el movimiento y la distribución uniforme de los preservadores (Honorato, 2002). Las dimensiones de los vasos se presentan en la Tabla 3.
Tabla 3 Dimensiones de los vasos de la madera de Quercus laurina y Q. crassifolia.
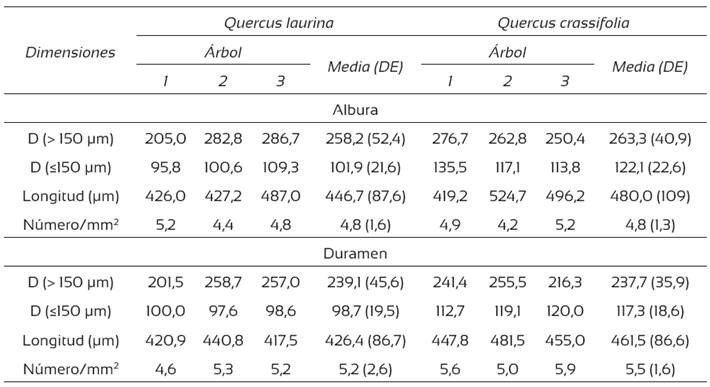
DE = desviación estándar; D = diámetro (μm)
La madera de Q. laurina presentó porosidad anular (Fig. 2A), similar a la encontrada para esta especie por De la Paz Pérez y Quintanar (2001)) y De la Paz Pérez y Dávalos (2008), pero diferente a la porosidad semianular obtenida por Aguilar y Castro (2006) y De la Paz Pérez et al. (2006). Los poros son de contorno oval y circular, solitarios y arreglados en hileras radiales, presentan tílides en la madera de duramen (Fig. 2B). Los elementos de vaso (Fig. 2C) presentan placas perforadas simples y puntuaciones areoladas alternas de forma oval (Fig. 2D), no presentan engrosamiento de la pared celular. El parénquima axial es apotraqueal difuso en agregados.
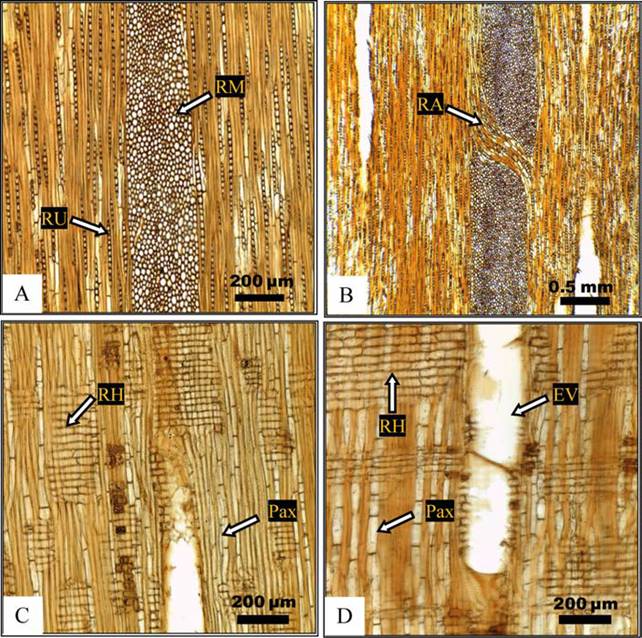
Figura 1 A) Quercus laurina, duramen árbol 3 (10x), RU = radio uniseriado; RM = radio multiseriado. B) Q. crassifolia, albura árbol 2 (4x); RA = radio agregado. C) Q. laurina, duramen árbol 1 (10x), RH = radio homogéneo, Pax = parénquima axial. D) Q. crassifolia, albura árbol 1 (10x), EV = elemento de vaso.
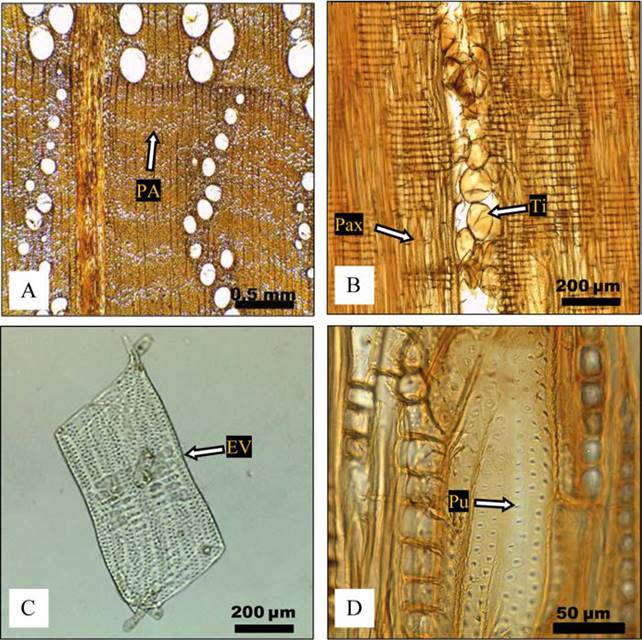
Figura 2 Quercus laurina A) albura árbol 2, porosidad anular, PA = parénquima apotraqueal difuso (4x). B) duramen árbol 1, Pax = parénquima axial, Ti = tílides (10x). C) EV = elemento de vaso en albura (madera temprana), material disociado árbol 1 (10x). D) Pu = puntuaciones areoladas alternas en elemento de vaso, duramen árbol 3 (40x).
La madera de Q. crassifolia, presentó porosidad anular (Fig. 3A), lo que coincide con lo descrito para la misma especie por De la Paz Pérez y Dávalos (2008) y De la Paz Pérez y Quintanar (2001). La porosidad anular mejora las características estéticas de la madera, haciendo el veteado más atractivo. Los poros son de contorno oval y circular, solitarios y arreglados en hileras radiales y presentan tílides en la madera de duramen (Fig. 3B, Fig. 4A). Los elementos de vaso presentan placas perforadas simples (Fig. 4B), y puntuaciones areoladas alternas de forma oval (Fig. 3D), con ausencia de engrosamientos de la pared celular. El parénquima axial es apotraqueal difuso en agregados. Se encontraron diferencias estadísticas entre las especies estudiadas en el número de poros por milímetro cuadrado en madera de duramen (p = 0,007), y en el diámetro de poros menores a 150 μm en albura y duramen (p < 0,001).
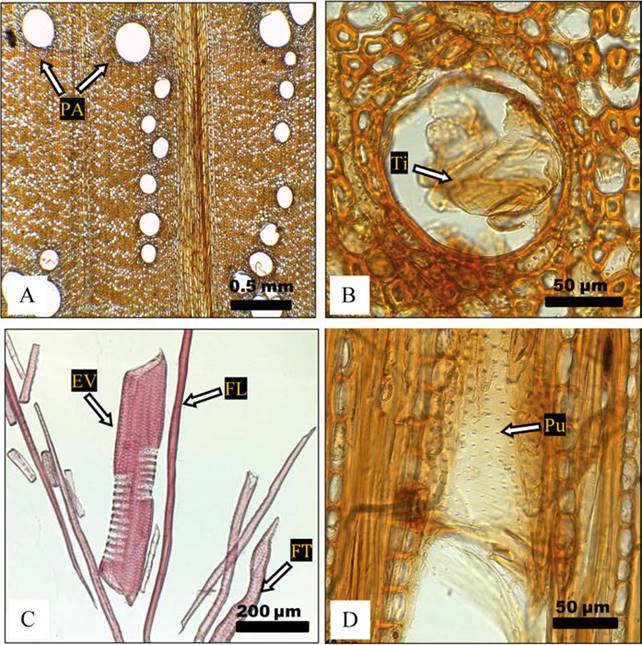
Figura 3 Quercus crassifolia, duramen árbol 2 A) porosidad anular (4x); PA = parénquima apotraqueal difuso. B) Ti = tílide, duramen árbol 3 (40x). C) EV = elemento de vaso en albura (madera tardía), material disociado árbol 3; FT = fibrotraqueida; FL = fibra libriforme (10x). D) Pu = puntuaciones areoladas alternas en elemento de vaso, albura árbol 1 (40x).
Fibras, fibrotraqueidas y traqueidas vasicéntricas
Las dimensiones de las fibras y las características de los radios multiseriados, son las variables que más influencia tienen en las propiedades físicas y mecánicas de la madera y para establecer la factibilidad de utilizar la madera en la producción de pulpa, papel, y tableros de fibras. Las dos especies presentan fibras de tipo libriforme, fibrotraqueidas y traqueidas vasicéntricas; en el duramen de Q. crassifolia se identificaron fibras septadas aisladas (Fig. 4D). Con respecto a la clasificación de IAWA (1937), las fibras de albura de ambas especies se clasifican como moderadamente largas, mientras que las de duramen como fibras medianas (Tabla 4). Con base en el diámetro, las fibras de albura se clasifican como medianas y las de duramen como finas; con referencia al espesor de la pared, Q. crassifolia presentó fibras de pared gruesa en albura y duramen, mientras para Q. laurina la pared es gruesa en albura y muy gruesa en duramen (Tortorelli, 1956).
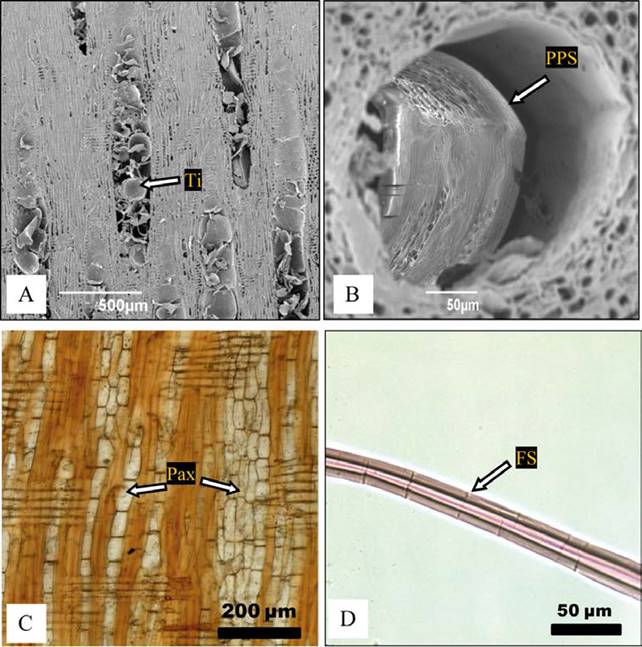
Figura 4 Quercus crassifolia. Microscopio electrónico de barrido: A) Ti = tílides en elemento de vaso, corte radial, duramen árbol 1. B) PPS = placa perforada simple, albura árbol 3. Microscopio de luz: C) Pax = parénquima axial, albura árbol 1 (10x). D) Fibra septada (FS), duramen árbol 2 (40x).
Tabla 4 Dimensiones de las fibras de la madera de Quercus laurina y Q. crassifolia.
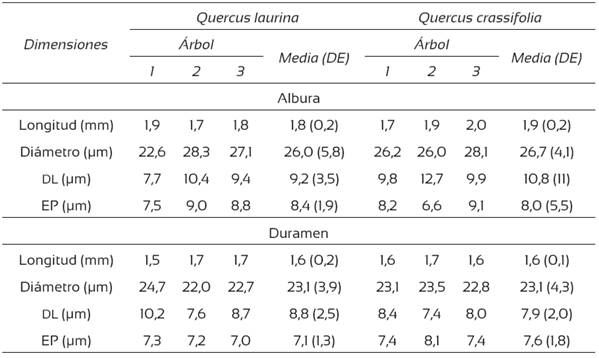
DL = diámetro del lumen; EP = espesor de pared (μm)
Se encontraron diferencias significativas entre la longitud (p < 0,001), el diámetro (p = 0,002) y el espesor de pared (p < 0,001) de fibras de albura y duramen de Q. laurina, y también entre la longitud (p < 0,001) y el diámetro (p < 0,001) de fibras de albura y duramen de Q. crassifolia. No se encontraron diferencias estadísticas significativas en los tres parámetros al comparar las fibras de albura-albura y duramen-duramen de las dos especies (p > 0,05).
Propiedades físicas
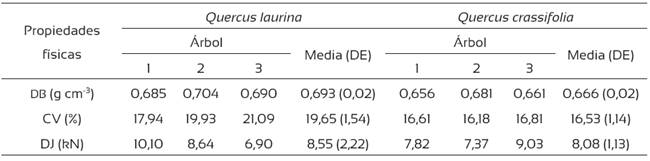
La densidad básica (DB) de la madera de Q. laurina fue de 0,693 g cm-3, superior a los 0,600 g cm-3 encontrados por Honorato y Fuentes (2001) para la misma especie y a los 0,652 g cm-3 de De la Paz Pérez y Dávalos (2008). En Q. crassifolia, la DB fue de 0,666 g cm-3, similar a los 0,687 g cm-3 hallados por De la Paz Pérez y Dávalos (2008) (Tabla 5). Las diferencias en la densidad de la madera ocurren por cambios en la proporción de los elementos constitutivos del xilema, como vasos, fibras y células de parénquima, y a la variación en el espesor de la pared celular del esclerénquima, atribuidas a las condiciones climáticas y de suelo de las distintas regiones donde crecen los individuos (Bernal y Terrazas, 2000; Montaño et al., 2013). La DB de las dos especies fue alta, de acuerdo con la clasificación de Sotomayor (2005). Como se esperaba de una madera de densidad alta, la contracción volumétrica de las dos especies también fue alta, de acuerdo con la clasificación de Chávez-Doldán (2007). Una contracción volumétrica alta indica una menor estabilidad dimensional de la madera, lo que precisa mayor cuidado en el proceso de secado de la madera, ya que habrá una mayor tendencia a la presencia de agrietamiento y rajaduras en el material.
La dureza de la madera de Q. laurina fue superior a la de Q. crassifolia (Tabla 5) y ambas especies se clasifican como de dureza alta, de acuerdo con Dávalos y Bárcenas (1999). La dureza, como las demás propiedades físicas de la madera, están directamente relacionadas con la DB; en este estudio, Q. laurina presentó una DB superior a Q. crassifolia, lo que es consistente con los valores de dureza encontrados.
Relación entre propiedades físicas y parámetros anatómicos
De acuerdo con el análisis de correlación, se encontraron correlaciones significativas entre las propiedades físicas y algunos parámetros anatómicos (Tabla 6). Para Q. laurina, las correlaciones positivas más altas se dieron entre la densidad básica (DB) y el ancho de los radios uniseriados (r = 0,98) y, como se anticipaba, entre la DB y el espesor de la pared de la fibra (EPF) (r = 0,97). También se encontraron correlaciones positivas altas entre la contracción volumétrica y el diámetro de lumen (DL) (r = 1,00) y entre el ancho de los radios multiseriados (ARM) y la dureza (r = 0,83). En Quercus crassifolia, se encontraron correlaciones positivas altas entre la DB y el DL (r = 0,99), entre la contracción volumétrica y el EPF (r = 0,97) y entre la dureza y el ARM (r = 1,00).
Tabla 6 Correlación entre propiedades físicas y parámetros anatómicos de la madera de Quercus laurina y Q. crassifolia.
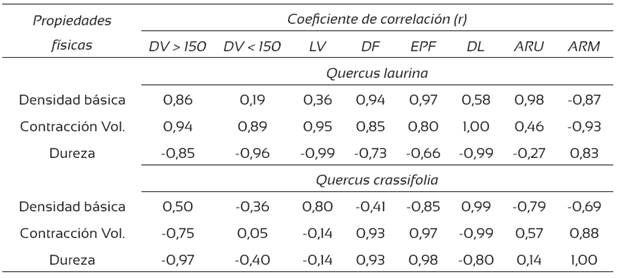
DV = diámetro del vaso; LV = longitud de los elementos del vaso; DF = diámetro de la fibra; EPF = espesor de pared de la fibra; DL = diámetro de lumen de las fibras; ARU = ancho de radios uniseriados; ARM = ancho de radios multiseriados
Cabe señalar que, junto con los vasos, las fibras presentan características distintas en las diferentes capas de las zonas de crecimiento en las angiospermas de porosidad anular como las aquí estudiadas. Sería deseable que los futuros esfuerzos en la descripción anatómica de los encinos incluyeran un análisis de las características anatómicas de la madera temprana y de la madera tardía, así como de la relación que guardan dichas características con las propiedades físicas (e.g. densidad) y mecánicas (e.g. resistencia al impacto) de la madera.
Conclusiones
La madera de Q. laurina presentó una tonalidad más clara con respecto a Q. crassifolia y el atractivo veteado de estos dos encinos permite disponer su utilización en productos decorativos. La madera de las dos especies pudo diferenciarse por el brillo alto y la textura media en Q. laurina, contra un brillo medio y una textura gruesa en Q. crassifolia. Las dos especies mostraron similitud en todos sus caracteres microscópicos, aunque en Q. crassifolia se encontraron fibras septadas. Estadísticamente se determinaron diferencias significativas entre las dos especies en el ancho de los radios uniseriados en albura y duramen, en el número de poros por milímetro cuadrado en el duramen y en el diámetro de poros menores a 150 μm en albura y duramen. Se determinó una correlación alta entre el espesor de pared de las fibras y la densidad básica en Q. laurina y entre el espesor de pared de las fibras y la contracción volumétrica en Q. crassifolia. Ambas especies mostraron correlación alta entre la contracción volumétrica y el diámetro de las fibras. Por otra parte, la dureza alta indica que la madera de estas especies puede utilizarse en la fabricación de pisos de tráfico pesado. Finalmente, la alta contracción volumétrica aquí determinada, conlleva a implementar programas de secado con condiciones de temperatura y humedad rigurosamente controladas.

