










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.29 no.3 Ciudad de México dic. 2012
Corals from the Maastrichtian Ocozocoautla Formation (Chiapas, Mexico) - a closer look
Corales de Maastrichtiano de la Formación Ocozocoutla (Chiapas, México)-una mirada más cercana
Hannes Löser
Estación Regional del Noroeste, Instituto de Geología, Universidad Nacional Autónoma de México, Av. Luis Donaldo Colosio s/n y Madrid, campus Universidad de Sonora, 83000 Hermosillo, Sonora, México. loeser@paleotax.de
Manuscript received: December 16, 2011
Corrected manuscript received: April 24, 2012
Manuscript accepted: May 14, 2012
ABSTRACT
The small Maastrichtian coral fauna of Ocozocoautla (Chiapas, Mexico) is being taxonomically reviewed. In contrast to an older study, the material is examined by preparing thin sections and carrying out systematic measurements to ensure that any comparisons of the material with existing species are of statistical significance. Twelve species in ten genera from the suborders Archeocaeniina, Fungiina, Meandrinina, Microsolenina and Poritina are described. The genera Multicolumnastraea and Favioseris are discussed in detail. One genus and its species are described as new: Filkornia is a colonial coral genus with very large calices that belongs to the Late Cretaceous family Felixaraeidae. All species are colonial corals from a shallow marine environment and many of them are known from localities outside the study area, mostly from the Maastrichtian of Jamaica.
Key words: scleractinian corals, taxonomy, Maastrichtian, Chiapas, Mexico.
RESUMEN
La taxonomía de la pequeña fauna de corales del Maastrichtiano de Ocozocoautla (Chiapas, México) está en proceso de revisión. En contraste con un estudio previo, el material se examinó en láminas delgadas y se tomaron mediciones sistemáticas de las dimensiones de los corales para asegurar que cualquier comparación con otras especies tenga significado estadístico. Se describen 12 especies en 10 géneros de los subórdenes Archeocaeniina, Fungiina, Meandrinina, Microsolenina y Poritina. Los géneros Multicolumnastraea y Favioseris se discuten con detalle. Un género y sus especies se describen por primera vez: Filkornia es un coral colonial con cálices muy grandes que pertenece a la familia Felixaraeidae del Cretácico Tardío. Todas las especies son corales coloniales de un ambiente marino somero. Casi todas las especies se conocen de otras localidades fuera del área del estudio, principalmente del Maastrichtiano de Jamaica.
Palabras claves: corales, taxonomía, Maastrichtiano, Chiapas, México.
INTRODUCTION
The whole Maastrichtian period was marked by a steady fall in sea level (Hardenbol et al., 1998) resulting in a decline in shallow marine faunas or in their erosion during the ensuing period of transgression. While Santonian and Campanian coral faunas can be found all over the globe (Santonian of Algeria, Austria, Azerbaijan, Egypt, France, Spain, Tunisia; Campanian of Bulgaria, France, Jamaica, Madagascar, Oman, Spain, Sweden, United States of America, United Arab Emirates; Löser et al., 2005), coral faunas from the Maastrichtian are rare. Moreover, several faunas originally considered Maastrichtian turned out to be older (Swinburne et al., 1992) or became Campanian after redefinition of the Campanian-Maastrichtian boundary (Odin, 1996). Most Maastrichtian coral faunas are represented by faunas of deeper marine environments whose small solitary corals are mainly ahermatypic. Important faunas of shallow marine environments, predominantly hermatypic corals, are known from a few areas, such as in Iran (Kühn, 1933; Khazaei et al., 2010, coral fauna under investigation by the author), Italy (Matteucci et al., 1982), Jamaica (Vaughan, 1899; Wells, 1934, 1935; Baron-Szabo, 2006, 2008), Northwest Mexico (Baron-Szabo et al., 2006), the Netherlands (Goldfuss, 1826, 1829; Umbgrove, 1925; Leloux, 2003, 2004; Löser, 2012), Spain (Bataller, 1936, 1937; Alloiteau, 1958), and the border area between the United Arab Emirates and Oman (Baron-Szabo, 2000).
The taxonomic composition of Maastrichtian coral faunas does not compare well with Santonian and Campanian faunas. Maastrichtian faunas contain many new elements. Nor do the few isolated Maastrichtian faunas compare well with each other because of the large geographic distances which separate them and varying palaeolatitudes. Not all known Maastrichtian coral faunas are taxonomically updated. All this makes it difficult comparisons of the faunas. The Maastrichtian coral fauna of the Ocozocoautla Formation (Chiapas, Mexico) was taxonomically first reported by Filkorn et al. (2005), but without using thin sections.
Corals have delicate, complexly structured skeletons. Some extant corals can be determined by usingjust a binocular microscope. In fossil members the view is obstructed by sediment within the skeleton. For a better understanding of their morphology, fossil corals should therefore be polished and sectioned. For illustrations thin sections are required in both transverse and longitudinal orientations. These methods have been known for over a century. If they are not applied either for technical reasons or because the material is in a poor state of preservation, systematic positioning and proper determination of the material cannot be guaranteed. The contribution by Filkorn et al. (2005) contained detailed descriptions, but the characteristics as stated were not reflected in the illustrations, which showed only complete specimens that had not undergone sectioning or surface polishing. The author had an opportunity to visit the study area and take new samples, which were then subjected to the standard procedure of sectioning, polishing, preparing thin sections and systematic measurement of calicular dimensions. The resulting taxonomy is presented here.
STUDY AREA
The observed material comes from the Ocozocoautla Fm. that crops out northwest of the Ocozocoautla town (Figure 1). There, a total thickness of 630 m of the Ocozocoautla Formation is estimated. This formation is mainly composed of conglomerate and sandstone at its base, grading upwards into fossil rich limestone in its middle part, and sandstone at the top, indicating a varying water depth (Vega et al., 2001). The corals are found in the limestone. The samples were collected somewhat to the west of sample location 5 (Filkorn et al., 2005) from an small area where vegetation were removed. Since the outcrop situation close to the highway is no longer as good as described by Vega et al. (2001) and Filkorn et al. (2005), it was not possible to make systematic collections or improve knowledge on sedimentological or taphonomical aspects. From the small outcrop area it was not clear whether the corals were in situ or not.
MATERIAL AND METHODS
Corals were easily collected as loose samples weathered out from soft sediments. The material is very well preserved. The coral skeletons are rarely deformed. In many cases they show fine septal ornamentation and even microstructures are preserved in places. All coral samples are small with a size of less than 60 mm (the holotype of Filkornia parasolitaria being the largest sample).
Most species indicated by Filkorn et al. (2005) were found in the locality, but original determinations are revised here. The material originally depicted by Filkorn et al. (2005) is kept in the Mexican National Palaeontological Collection at the Universidad Nacional Autónoma de México, Instituto de Geología (IGL), and is numbered 8432 to 8443. It was available for study but was not examined by sections or thin sections. Some topotypical material in the IGL collection numbered 9281 to 9285 was used to prepare thin sections. The newly collected samples numbered L-7000 to L-7045 are stored in the Palaeontological Collection of the Universidad Nacional Autónoma de México, Instituto de Geología, Estación Regional del Noroeste (Hermosillo; acronym ERNO).
The following Maastrichtian coral material (contained in museum and institutional collections; collection acronyms as indicated below) was studied for comparison:
Maastrichtian corals described by Goldfuss (1826, 1829; IPB), Leloux (2003, 2004; NNM, DGP), Umbgrove (1925; NNM) and Löser (2011, 2012; IPB, NNM, DGP) from the Netherlands.
Campanian/Maastrichtian corals from Jamaica described by Duncan and Wall (1865, NHM), Vaughan (1899; USNM and MCZ), Wells (1934; USNM), Wells (1935; NHM) and Baron-Szabo (2006, 2008; USNM).
Maastrichtian corals described by Alloiteau (1958), Vidal (1917, 1921), Bataller (1936, 1937), Reig Oriol (1987, 1992; MGSB) Reig Oriol and Vilella (1995; MGSB) from Spain.
Campanian/Maastrichtian corals from the OMAN/U.A.R. border area described by Baron-Szabo (2000; NHM) and own samples (ERNO).
Maastrichtian corals described by Baron-Szabo et al. (2006) from San Luis Potosí, Mexico (IGM). Maastrichtian corals mentioned by Khazaei et al. (2010) from the SW Iran (UB), which are under investigation.
The sample material was prepared as transverse and longitudinal polished and thin sections. Examination was carried out, using a Zeiss Stemi-2000 binocular microscope. Thin sections were scanned by an Epson Perfection flat-bed scanner with an attached transparency unit at an optical resolution of up to 6400 dpi (dots per inch). The grey-scale bitmaps obtained were improved by grey-scale stretching. Illustrations were directly obtained by scanning thin sections.
To gain more insight into the intraspecific variation of fossil corals and to obtain a better strategy for comparing species, calicular dimensions in thin sections obtained from one to three different samples of each species were systematically measured. To achieve statistical significance, the largest number of possible measurements was taken. This number was mainly determined by the size and quality of the thin section and the size of the single calices in relation to the size of the thin sections. Thin sections were measured and values were calculated using the program PaleoTax/Measure (www.paleotax.de/measure/; Löser 2012, accepted).
SYSTEMATICS
The following abbreviations stand for the various collections:
DGP, Ammonietenhoeve/De Groene Poort, Boxtel, The Netherlands;
ERNO, UNAM, Instituto de Geología, Estación Regional de Noroeste, Universidad Nacional Autónoma de México, Hermosillo, Mexico;
IGM, Instituto de Geología, Mexico City, Mexico;
IPB, Universität Bonn, Geologisch-Paläontologisches Institut, Bonn, Germany; MB, Museum für Naturkunde der Humboldt-Universität Berlin, Germany;
MCZ, Havard University, Museum of Comparative Zoology, Cambridge, Mass., USA;
MGSB, Museo Geológico del Seminario de Barcelona, Spain;
MNHN, Muséum National d'Histoire Naturelle, Paris, France;
NHM, The Natural History Museum, London, England; NNM, Naturalis - Nationaal Natuurhistorisch Museum, Leiden, The Netherlands;
UB, University of Birjand, Department of Geology, Birjand, Iran;
USNM, United States National Museum, Washington, D.C., USA.
The following abbreviations are used for the dimensions of corals:

For each type of measurement (calicular diameter and distance, width and distance of calicular row) in one thin section the following values were obtained:

The abbreviations used in the synonym lists follow Matthews (1973):

Suborder Archeocaeniina Alloiteau, 1952
Family Actinastraeidae Alloiteau, 1952
Genus Actinastrea d'Orbigny, 1849
Actinastrea reticulata (Goldfuss, 1829) Figure 2a-2c
*v Astrea reticulata Goldfuss, 1829, p. 111, pl. 38, fig. 10 v Actinastrea ramosa Michelin. Löser, 1998, p. 79, pl. 1, fig. 1
v Actinastrea benaixensis Alloiteau, 1954. Baron-Szabo, 1998, p. 128, pl. 1, fig. 1 v Actinhelia elegans Goldfuss. Filkorn et al., 2005, p. 124, fig. 2i
Dimensions. See Table 1.
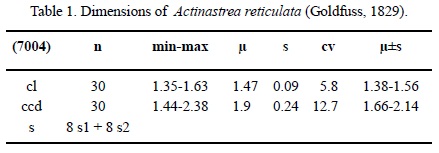
Description. Massive cerioid colony. Calicular outline circular. Calicular pit depressed. Septa compact, in cross section thick on the outside, getting very thin towards the centre. Symmetry of septa radial and regularly octomeral. Cycles of septa regular. Two cycles. Septal cycles differ in length. First septal cycle reaches up to the centre of the calice, the following one is shorter. Septa of the second cycle occasionally attached to those of the first cycle. Septal upper margin not preserved, lateral face shows occasional small thorn, inner margin smooth. Pali or paliform lobes absent. All septa of the first cycle and some of the second cycle are attached to the columella. Costae hardly present, non-confluent, with thick granules on the surface. Synapticulae absent. Columella styliform. Endotheca consists of a few thin tabulae. Wall compact, septothecal. Coenosteum narrow (up to 25 % of the calicular diameter). It consists of costae and single large trabeculae and is granulated on the surface. Budding extracalicinal.
Material. ERNO L-7004, L-7005, L-7010, IGM 8440.
Discussion. Actinhelia d'Orbigny, 1849 is based on Actinhelia elegans (Goldfuss, 1826), a Maastrichtian coral from the St. Pietersberg hill close to Maastricht (The Netherlands). Actinhelia is closely related to Montipora Blainville, 1830 or Astreopora Blainville, 1830. The material under discussion belongs to the genus Actinastrea s.s. (Löser, 2012). Beauvais (1982) doubted that the only available syntype of A. reticulata (IPB GF296) was identical with the illustration in Goldfuss (1829). He assigned the illustrations in Goldfuss (1829) to various species. This is, however, doubtful as no scale is indicated for the enlarged details so that they cannot be assigned. The originally illustrated type is for the present assumed to be the (only) syntype.
Occurrence. Santonian of Austria (Upper Austria), Gosau. Early Campanian of Turkey (Iller Amasya) Amasya, Hobek Tepe. Early Late Campanian of Spain (Cataluña, Lérida) Com. Pallars Jussà, Mun. Pallars Jussà, Pobla de Segur, Torallola. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla.
Genus Columactinastraea Alloiteau, 1952
Columactinastraea hilli (Wells, 1934) Figure 2d-2f
v Centrastrea hilli Wells, 1934, p. 80, pl. 2, figs. 11, 12.
v Columactinastraea parvistella Reig Oriol, 1992, p. 32, pl. 7, figs. 2, 3.
v Columactinastraea pygmaea Felix. Baron-Szabo, 2003, p. 116, pl. 1, figs. 1, 3, 4, 6, pl. 2, figs. 1-7
v Actinastrea sp. Filkorn et al., 2005, p. 118, fig. 2a
Dimensions. See Table 2.
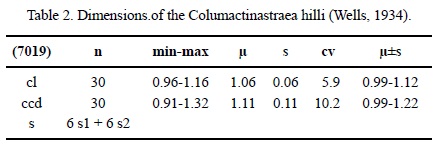
Description. Massive cerioid colony. Calicular outline circular. Calicular pit depressed. Septa compact, in cross section thick on the outside, getting thinner towards the centre. Symmetry of septa radial and regularly hexameral. Cycles of septa regular. Two cycles. Septal cycles differ in length. First septal cycle reaches up to the centre of the calice, the second one is shorter. Septa free. Septal upper margin not preserved, lateral face with numerous small thorns. Pali on the first septal cycle. Some septa of the first cycle may be attached to the columella. Costae hardly present, non-confluent. Synapticulae absent. Columella styliform. Endotheca consists of numerous thin tabulae. Wall compact, septothecal. Coenosteum absent. Budding extracalicinal.
Material. ERNO L-7001, L-7019, L-7031, L-7032, IGM 8432.
Discussion. Filkorn et al. (2005) did not consider this taxo-nomic name to be a possible candidate for their material. It was originally assigned to Centrastrea d'Orbigny, 1849 (= Thamnasteria Lesauvage, 1823). The holotype (USNM I-74492) was available for study and was found to coincide with the material, even though uncut and not available in thin sections.
Occurrence. Early Santonian of Austria (Upper Austria) Gosau, Grabenbach. Late Santonian of Austria (Upper Austria) Russbach-Hochmoos. Early Late Campanian of Spain (Cataluña, Lérida) Com. Pallars Jussà, Mun. Pallars Jussà, Pobla de Segur, Torallola. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla. Late Maastrichtian of Jamaica (St. James) Catadupa.
Suborder Fungiina Verrill, 1868-70
Family Felixaraeidae Beauvais, 1982
Genus Filkornia new genus
Type species. Filkorniaparasolitaria new species.
Diagnosis. Astreoid colony with large calices and large septal numbers.
Description. Astreoid colony. Calices large. Septa regularly but not abundantly perforated. Microstructure of large tra-beculae. Septa in cross section thick on the outside, getting thinner towards the centre. First cycles disproportionately thick compared with younger ones. Symmetry of septa irregularly radial. Cycles of septa subregular. Septal cycles differ in length and thickness. Septa free. Septal upper border coarsely granulated, lateral face with numerous large thorns, inner margin slightly swollen in places. Pali or paliform lobes absent. Some septa of the first cycle are attached to the columella. Costae present, but short, non-confluent. Synapticulae present. Columella consists of a large group of isolated trabeculae, endotheca of numerous thin tabulae. Wall compact, septothecal. Coenosteum absent. Budding intracalicinal.
Etymology. Named after Harry Filkorn, who was the first to investigate the taxonomy of the Ocozocoautla coral fauna.
Occurrence. Maastrichtian of Mexico and Jamaica.
Discussion. The new genus can be compared with Felixaraea Beauvais, 1982 and Pseudofavia Oppenheim, 1930. It was found to differ from the first by its colonial growth form and from the second by its astreoid growth mode and a much larger number of septa. In its calicular centre it resembles Paracycloseris Wells, 1933, which also may belong to the Felixaraeidae (and not to the Cyclolitidae Alloiteau, 1952 as proposed by Baron-Szabo, 2008, because it lacks pen-nulae). The new genus represents a particular growth form. It starts as a solitary coral on a small stem or attached to another coral or to a shell. The calice may reach a large diameter before new calices are formed. For that reason the type species is considered a solitary coral in the literature.
The calice divides after reaching a certain size. When the colony is growing larger, older calices are often completely overgrown (Figures 3a, 3f).
Filkornia parasolitaria new species
? Trochoseris catadupensis Vaughan. Wells, 1934, p. 78, pl. 2, figs. 9, 10.
v Trochoseris catadupensis Vaughan. Filkorn et al., 2005, p. 123, fig. 2h.
v Trochosmilia hilli Vaughan. Filkorn et al., 2005, fig. 2d v Cyathoseris formosa d'Achiardi. Baron-Szabo et al., 2006, p. 1041, fig. 5.7.
v Mycetophyllia multistellata Reuss. Baron-Szabo, 2006, p. 42, pl. 5, fig. 5, pl. 7, fig. 2, 3.
Diagnosis. Filkornia with a calicular diameter of 25 to 35 mm and a variable number of septa ranging from 190 to 240.
Description. Astreoid colony. Calicular outline circular to elliptical. Calicular pit depressed. Septa regularly but not abundantly perforated. Septa in cross section thick on the outside, getting thinner towards the centre. First cycles disproportionately thick compared with younger ones. Septal maximal thickness ca. 1 mm. Symmetry of septa irregularly radial. Cycles of septa subregular. Number of septa varies between 190 - 240, depending on the size of the individual calice. Septal cycles differ in length and thickness. First three septal cycles reach up to the centre of the calice, the following ones are shorter. Septal upper border coarsely granulated, lateral face with numerous large thorns, inner margin slightly swollen in places. Pali or paliform lobes absent. Some septa of the first cycle are attached to the columella. Costae present, but short, non-confluent. Ornamentation of the costae not preserved. Synapticulae present, abundant. Columella consists of a large group of isolated trabeculae, endotheca of numerous thin tabulae. Wall compact, septothecal. Coenosteum absent. Budding intracalicinal.
Etymology. After its strange growth form, pretending to be a solitary coral.
Types. Holotype ERNO L-7043, paratypes ERNO L-7008, 7012, 7018. All types from the Maastrichtian (Ocozocoautla Formation) of Ocozocoautla (Chiapas, Mexico), WGS84 16°48'51"N 93°22'12"W.
Dimensions. See Table 3.
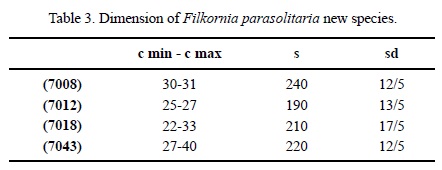
Material. ERNO L-7008, L-7012, L-7017, L-7018, L-7040, L-7041, L-7042, L-7043, L-7044, IGM 8435, 8439.
Occurrence. Maastrichtian (Ocozocoautla Formation) of Ocozocoautla (Chiapas, Mexico); Maastrichtian (Upper Cárdenas Fm.) of Arroyo de la Atarjea (Cárdenas, San Luis Potosí, Mexico); Maastrichtian (Guinea Corn Fm.) of various localities in Jamaica.
Discussion. Material of Filkornia parasolitaria as described before for various genera cannot be applied to this material. Mycetophyllia Milne Edwards and Haime, 1848b, as a member of the Mussidae, suborder Faviina, has compact septa, dentated upper septal margins and almost smooth lateral faces. The columella is poorly developed. It is moreover a coral with a meandrinoid calicular arrangement. Mycetophyllia has been not so far indicated in the Cretaceous. The present material has perforated septa, strongly ornamented lateral septal faces and a well-defined columella. Although the general appearance of Cyathoseris Milne Edwards and Haime, 1849 is reminiscent of this genus (see for instance Pfister, 1980), the genus cannot be applied to the present material. Cyathoseris forms thamnasterioid colonies with a circumoral budding mode. Septa are compact in Cyathoseris.
Both solitary coral species created by Vaughan (1899) and applied to the present material by Filkorn et al. (2005) are poorly defined corals. The types were at hand. One (T. hilli Vaughan, 1899) is unsectioned, the other (T. catadupensis Vaughan, 1899) has a polished surface, which is not well preserved.
Family Fungiidae Dana, 1846
gen. et sp. indet.
v Paracycloseris elizabethae Wells. Filkorn et al., 2005 p. 125, fig. 2k.
vp Paracycloseris nariensis Duncan. Baron-Szabo, 2008, p. 180, pl. 17, fig. 7-9, non 1-6.
Dimensions. See Table 4.

Description. Solitary cupolate coral. Calicular outline circular. Septa irregularly perforated, more in younger than in older septa. Microstructure of medium-sized trabeculae. Septa in cross section thin on the outside, becoming thicker and then thinner again towards the centre. Symmetry of septa radial and regularly hexameral. Cycles of septa sub-regular. Septal cycles differ in length and thickness. First two septal cycles reach up to the centre of the calice, the following ones are shorter. Septa free. Septal upper border coarsely granulated, lateral face with numerous small granules, inner margin smooth. Pali or paliform lobes absent. Costae hardly present, ornamentation not preserved. Synapticulae present, abundant, compound (fulturae). Columella in the form of a group of trabecular elements. Endotheca absent. Wall compact, septothecal.
Material. IGM 8442, 9281-9285.
Discussion. The sample taken by Filkorn et al. (2005) is not sectioned and its characteristics are unknown. Description and illustration are based on additional collection material. The description given in Filkorn et al. (2005) corresponds to that given for Paracycloseris elizabethae Wells, 1934, but not to the material from the study area.
Under the name Paracycloseris elizabethae Wells, 1934, Baron-Szabo (2008, pl. 17) lumps together material which does not belong to the same species, genus and family: figures 1 and 2 represent Paracycloseris elizabethae Wells, 1934 sensu stricto; figures 3 and 4 are refigurations of Trochocyathus nariensis Duncan, 1880 (a Cenozoic species); figure 5 is Leptocyathus hexacristatus Alloiteau, 1936 sensu stricto; figures 7 to 9 correspond to the material represented here. Paracycloseris elizabethae belongs to the family Felixaraeidae, the systematic positions of Trochocyathus nariensis and Leptocyathus hexacristatus are questionable as the degree of perforation of the septal blades and the existence of synapticulae in this material are unknown. It cannot be ruled out for both species that the perforations shown in the figures are just preservation bias. The material corresponding to the material from Chiapas shows that there are synapticular bars (fulturae), which clearly indicates its position in the Fungiina suborder. As it was not possible to collect new material and only a few specimens could be loaned from the rich IGM collection, a new taxon cannot be established at the moment for this material.
Occurrence. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla. Late Maastrichtian of Jamaica (Clarendon) Grantham, Rio Minho.
Suborder Meandrinina Alloiteau, 1952
Family Pachyphyllidae Alloiteau, 1957
Genus Favioseris Wells, 1934
Favioseris anómalos Wells, 1934
v Favioseris anomalos Wells, 1934, p. 82, pl. 4, fig. 19, 20
v ? Dichocoenia trechmanni Wells, 1934, p. 75, pl. 2, fig. 7, 8
v Barysmilia trechmanni Wells. Filkorn et al., 2005 p. 119, fig. 2c; Baron-Szabo, 2006, p. 85, pl. 16, fig. 4, 7
v Favia? gregoryi Wells. Filkorn et al., 2005, p. 122, fig. 2f
vp Dichocoenia anomalos Wells. Baron-Szabo, 2006 p. 81, pl. 16, fig. 2 non 8
Dimensions. See Table 5.
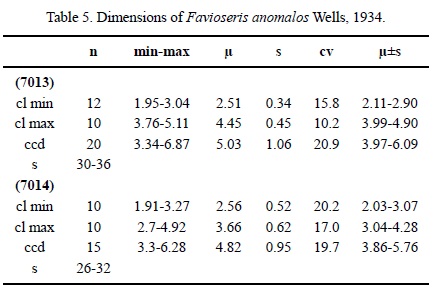
Description. Massive plocoid colony. Calicular outline elliptical. Calicular centres slightly depressed, margins elevated. Septa compact. Microstructure of small-sized trabeculae, septa with a median dark line. Septa (and costae) in cross section in the wall thick, thinner towards the centre. Symmetry of septa irregular. Septal generations differ in length and thickness. First septal generation reaches 40-45 % of the (shorter) calicular diameter, the following generations are shorter. Septa free. Septal upper margin smooth, lateral face with thorns, inner margin T-shaped or swollen in places. Pali or paliform lobes absent. Some septa may be attached to the columella. Costae present, non-confluent, smooth on the surface. Synapticulae absent. Columella small, lamellar. Endotheca of thin tabulae. Wall sub-compact, septothecal. Coenosteum of medium width (about 30 % of the calicular diameter) consisting of costae and numerous dissepiments. Budding extracalicinal.
Material. ERNO L-7013, L-7014, L-7015, L-7033, L-7034, L-7035, IGM 8434, 8437.
Discussion. The present material coincides with the original description and with the holotype of the species (USNM I-74480). Hence, the holotype is a specimen without thin sections and with only a very small polished surface. A detailed analysis of the septal microstructure, the calicu-lar centre and the endotheca is not possible. Topotypical material from the type area was examined. But there are various species which fit the outer appearance of Favioseris anomalos so that our knowledge cannot be widened without sectioning the type specimen.
The material cannot be assigned to Ovalastrea (as proposed by Wells, 1935) because septa are compact and free and synpaticulae are absent. In Ovalastrea, septa are perforated at their inner margins, often connected to each other and synapticulae are present.
The material cannot be placed in the genus Barysmilia Milne Edwards and Haime, 1848a for two reasons: firstly, the holotype of the type species of Barysmilia (Dendrophyllia brevicaulis Michelin, 1841; MNHN M01126) is from Uchaux (Vaucluse, France) and is so poorly preserved that it can hardly be assigned even to a family. It is erroneous to think that Barysmilia is a rhipidogyrid genus (as claimed by Baron-Szabo, 1998, 2006). The type of the type species does not allow this conclusion. The family Rhipidogyridae became moreover extinct before the Albian (Löser, 2005). The material cannot be assigned to Dichocoenia Milne Edwards and Haime, 1848a. Although Dichocoenia has a "meandrinid" microstructure consisting of small trabeculae comparable to the present material, its wall is septothecal and very thick, the costae are very short, and the coenosteum is dense and consists of thick extracalicinal tabulae. Occurrence. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla. Late Maastrichtian of Jamaica (Clarendon), Grantham, Rio Minho; Logie Green section. (?) Late Maastrichtian of Jamaica (St. James), Catadupa.
Favioseris sp.
v Dichocoenia anomalos (Wells, 1934) - Baron-Szabo, 2006, p. 81, pl. 16, fig. 8
Dimensions. See Table 6.
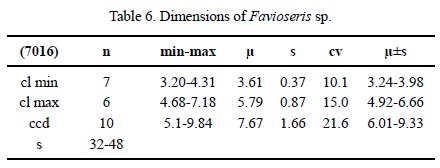
Description. Massive plocoid colony. Calicular outline circular to elliptical. Calicular centres slightly depressed, margins elevated. Septa compact. Microstructure of small-sized trabeculae, septa with median dark lines. Septa (and costae) in cross section in the wall thick, thinner towards the centre. Symmetry of septa irregular. Septal generations differ in length and thickness. First septal generation reaches 40-45 % of the (shorter) calicular diameter, following generations are shorter. Septa free. Septal upper margin smooth, lateral face with thorns, inner margin T-shaped in places. Pali or paliform lobes absent. Some septa may be attached to the columella. Costae present, short, non-confluent, smooth on the surface. Synapticulae absent. Columella lamellar. Endotheca of thin tabulae. Wall perforated, septoparathecal. Coenosteum of medium width (ca. 30 % of the calicular diameter) consists of isolated fragments of costae and few dissepiments. Budding extracalicinal.
Material. ERNO L-7016.
Discussion. The material differs from Favioseris anomalos by the very short costae, the perforate wall and a divergent coenosteum.
Occurrence. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla. Campanian to Maastrichtian of Jamaica.
Family Phyllosmiliidae Felix, 1903
Genus Pachygyra Milne Edwards and Haime, 1848a
Discussion. Pachygyra was formerly assigned to the poorly defined Meandrinidae family (see Löser et al., 2010). Meandrina Lamarck, 1801 might be characterised by small trabeculae but it has smooth septal inner borders, whereas many Late Cretaceous so-called Meandrinids have T-shaped or rhopaloid septal inner margins. Forms with small trabeculae and T-shaped or rhopaloid septal inner margins are separated in the Phyllosmiliidae family. Material of Turbinolia basochesi Defrance, 1828, type species of Phyllosmilia de Fromentel, 1862, from the type locality (France, Boche-du-Rhône, Marseille, port of Figuière; Late Santonian) was examined and confirms the above morphological features.
Pachygyra cf. ilerdensis (Reig Oriol, 1989)
v Dictuophyllia conferticostata Vaughan. Filkorn et al., 2005, p. 123, fig. 2g.
Dimensions. See Table 7.
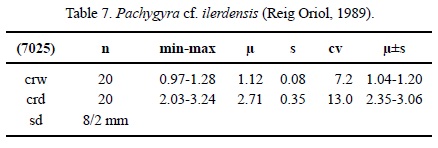
Description. Massive meandrinoid colony. Calicular rows slightly elevated over the colony surface. Calicular rows long and curved. Calices indistinct. Septa compact. Microstructure of small-sized trabeculae, septa with median dark lines. Septa in cross section bicuneiform close to the wall, thinner towards the centre. Symmetry of septa irregular. Septal generations differ in length and thickness. First septal generation reaches up to the centre of the row, the following generations are shorter. Septa free. Septal upper border smooth, lateral face with small thorns, inner margin slightly swollen in places. Pali or paliform lobes absent. Costae present, sub-confluent to non-confluent, smooth on the surface. Synapticulae absent. Columella as a continuous lamella. Endotheca consists of thin tabulae. Wall compact, septothecal. Coenosteum broad, consisting of costae and exothecal dissepiments. Budding intracalicinal.
Material. ERNO L-7009, L-7011, L-7024, L-7025, L-7026, L-7027, L-7028, IGM 8438.
Discussion. The name Dictuophyllia Blainville, 1830 should not be used any more since the type of the type species is an external mould which does not provide much information concerning the skeleton microstructure. Pachygyra conferticostata cannot be used as a specific name on account of its smaller dimensions (width of calicular rows 0.8 - 1 mm) and higher septal density (12/2 mm; data from MCZ 114212, syntype of Diploria conferticostata Vaughan, 1899). The material compares well with Pachygyra ilerdensis, but the septal density of the latter is much lower (5/2 mm). As there are no polished or thin sections, it cannot be ruled out that the septa of younger generations are only visible in thin sections.
Occurrence. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla.
Suborder Microsolenina Morycowa and Roniewicz, 1995
Family Leptophylliidae Vaughan, 1905
Genus Astraeofungia Alloiteau, 1952
Astraeofungia felixi (Reyeros-Navarro, 1963)
v Thamnasteria felixi Reyeros-Navarro, 1963, p. 6, pl. 2, fig. 1, 3, 4.
v Brachyseris morchella Reuss. Reyeros de Castillo, 1983 p. 22, pl. 9, fig. 2, 3.
v Synastrea sp. cf. S. agaricites Goldfuss. Filkorn et al., 2005 p. 125, fig. 2l.
Material. IGM 8443.
Discussion. It was not possible to collect any new material of this species. The specimen in Filkorn et al. (2005) is comparable with A. felixi in the distance of calices and the number of septa. No thin sections are known of the holo-type of Astraeofungia felixi; the thin section illustrated in Reyeros-Navarro (1963: pl. 2, fig. 4) is part of a paratype. The material cannot be representative of Synastrea Milne Edwards and Haime, 1848b as the septa of this genus are much thicker and less perforated.
Occurrence. Aptian of Mexico (Puebla), San Juan Raya. Campanian to Maastrichtian of Mexico (Oaxaca), San Pedro Yucunama. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla.
Family Microsolenidae Duncan, 1884
Genus Microsolena Lamouroux, 1821
?Microsolena sp.
v Mesomorpha catadupensis Vaughan. Filkorn et al. , 2005, p. 124, fig. 2j
Dimensions. See Table 8.
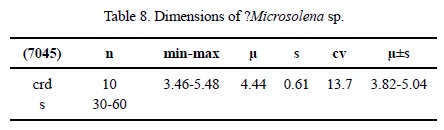
Description. Encrusting or massive thamnasterioid colony. Calicular centres slightly depressed. Septa regularly perforated. Microstructure of large trabeculae. Septa in cross section of equal thickness throughout. No septal symmetry. Half of all septa reach up to the calicular centre. Septa free, upper border with large regular granules. Septal lateral face with pennulae, inner margin smooth. Pali or paliform lobes absent. Costae present, confluent. Synapticulae present, abundant. Columella poorly defined, probably one granule. Endotheca absent. Wall absent. Coenosteum poorly defined because of the type of calicular arrangement. Coenosteum consisting of costae. Budding intracalicinal, completely polystomedal.
Material. ERNO L-7007, L-7021, L-7022, L-7023, L-7036, L-7045, IGM 8441.
Discussion. The material described by Vaughan (1899) as Mesomorpha catadupensis does not belong to Mesomorpha Pratz, 1882-83 but to Synastrea. Mesomorpha is a conceptual genus (the type of the type species is lost) and is generally considered to be a thamnasterioid coral with very small, widely spaced calices, a few non-pennular compact septa and a styliform columella, while Synastrea has large calices and strong pennulae-bearing septa which are only perforated at the inner margin.
The sample depicted by Filkorn et al. (2005) and additional material cannot be identified with Synastrea catadupensis. Like Synastrea it is a thamnasterioid coral with large calices but its septa are greatly perforated. For the moment it may be considered a Late Cretaceous variety of Microsolena, although it differs from it by its broad interseptal spaces and lower septal density. In most cases the newly collected material consists of initial stages of colonies. Only a few larger colonies were found. Occurrence. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla.
Suborder Poritina Veron, 1995
Discussion. The suborder was established without diagnosis, but is nevertheless, valid. Here, the families Actinacididae and Poritidae are included in this suborder. Their former position, the suborder Fungiina, was preliminary. The Fungiina gather various families of which some do not correspond to the name giving genus Fungia and a stepwise classification of these families into other suborders is recommendable (see also Morycowa and Roniewicz, 1995).
Family Actinacididae Vaughan and Wells, 1943
Genus Actinacis d'Orbigny, 1849
Actinacis magna Alloiteau, 1958
*v Actinacis magna Alloiteau, 1958, p. 179, fig. 26, pl. 16, fig. 3, 4, pl. 18, fig. 6.
Dimensions. See Table 9.
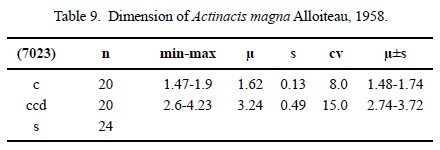
Description. Thick-branching plocoid colony. Calicular outline circular. Calicular pit not depressed. Septa irregularly perforated. Septa in cross section slightly thicker on outside, getting thinner towards the centre. Bilateral symmetry of septa. Cycles of septa subregular. Septal cycles differ in length, hardly in thickness. First septal cycle reaches up to the centre of the calice, following cycles are shorter. Septa of younger cycles are often and regularly connected to septa of older ones, upper border not preserved. Septal lateral faces with few thorns, inner margins smooth. Irregular presence of paliform lobes on first cycle. Most septa of first cycle attached to columella. Costae present, of varying length, generally short, non-confluent, surface not preserved. Synapticulae present. Columella lamellar. Endotheca consisting of numerous tabulae of regular thickness. Wall subcompact, septothecal, with synapticulae. Coenosteum broad, consisting of isolated trabeculae and tabulae. Budding extracalicinal.
Material. ERNO L-7023, L-7029.
Occurrence. Coniacian of Madagascar, Pointe d'Antonibe, Ankotrofotsy. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla.
Genus Multicolumnastraea Vaughan, 1899
Discussion. Multicolumnastraea was assigned to the Agatheliidae Beauvais and Beauvais, 1975 by Filkorn et al. (2005) and Baron-Szabo et al. (2006), but differs from this family in many characteristics. Its septal faces are almost completely smooth (while those of Agatheliidae have numerous thorns), the columella is styliform (absent or weak in Agatheliidae) and the wall is thin and septo-parathecal (thick and trabecular in Agatheliidae). Isolated trabeculae in the coenosteum are the only similarity. They are much smaller and finer in Agatheliidae than they are in Multicolumnastraea. The genus is very similar to Actinacis. Actinacis itself is ill defined. Its type species is hardly known. All illustrations are from places other than the type locality. Thin sections of the type are poorly preserved. More material from the type locality of the type species of both Actinacis and Multicolumnastraea has to be studied to understand the differences between the genera.
Multicolumnastraea cyathiformis (Duncan and Wall, 1865)
*v Heliastraea cyathiformis Duncan and Wall, 1865, p. 8, pl. 1, fig. 1.
Multicolumnastraea cyathiformis Duncan. Vaughan, 1899, p. 236, pl. 37, fig. 5-7, pl. 38, fig. 1; v Filkorn et al., 2005, p. 118, fig. 2b .
v non Multicolumnastraea cyathiformis Duncan. Baron-Szabo, 1998, p. 131, pl. 6, fig. 4 [= Neocoenia renzi] non Multicolumnastraea cyathiformis Duncan. Baron-Szabo, 2003 p. 119, pl. 3, fig. 7 [= ? Hydnophoropis sp.].
v non Multicolumnastraea cyathiformis Duncan. Baron-Szabo et al., 2006, p. 1037, fig. 4.3 [= Placocolumastreasp.] .
Dimensions. See Table 10.
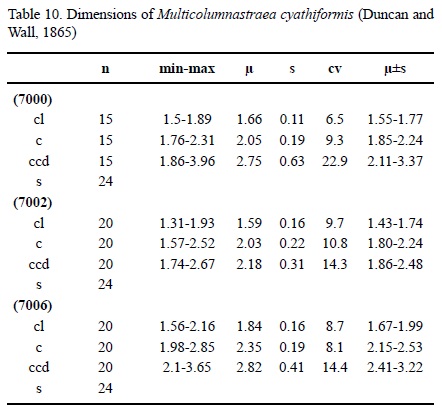
Description. Massive plocoid colony. Calicular outline circular. Calicular centres slightly depressed, margins slightly elevated. Septa compact. Septa in cross section slightly thicker on outside, then of equal thickness. Symmetry of septa radial and regularly hexameral. Cycles of septa regular. Three cycles. Septal cycles differ in length and thickness. First septal cycle reaches 35 % of calicular diameter, following cycles shorter. Septa free, upper border smooth. Septal lateral face with some small thorns, inner margin smooth. Pali present on two septal cycles. Septa only rarely attached to columella. Costae present, of varying length, generally short, non-confluent, surface with large granules. Synapticulae present, mainly in the coenosteum. Columella small, substyliform. Endotheca consisting of regular thick tabulae. Wall generally compact, septoparathecal. Coenosteum broad, consisting of isolated trabeculae and tabulae. Budding extracalicinal.
Material. ERNO L-7000, L-7002, L-7006, L-7020, L-7030, L-7037, L-7038, L-7039, IGM 8433.
Discussion. The material depicted by Baron-Szabo (1998), Baron-Szabo (2003), and Baron-Szabo et al. (2006) differs from Multicolumnastraea cyathiformis by having no pali and a complete different coenosteum.
Occurrence. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla. Late Maastrichtian of Jamaica (Clarendon) Mount Hindmost; Trout Hall. Late Maastrichtian of Jamaica (St. James) Catadupa.
Multicolumnastraea sp. Figure 4k-4l
Dimensions. See Table 11.
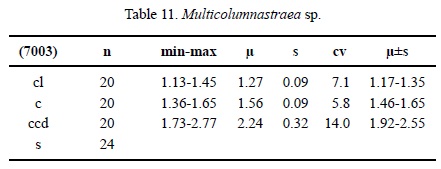
Description. Massive plocoid colony. Calicular outline circular. Calicular centres slightly depressed, margins slightly elevated. Septa compact. Septa in cross section slightly thicker on outside, then of equal thickness. Symmetry of septa radial and regularly hexameral. Cycles of septa regular. Three cycles. Septal cycles differ in length and thickness. First septal cycle reaches 35 % of calicular diameter, other cycles shorter. Septa connected with each other in places, upper border smooth. Septal lateral face with some small thorns, inner margin smooth. Pali present on two septal cycles. Septa rarely attached to columella. Costae present, of varying length, generally short, non-confluent, surface with large granules. Synapticulae present, mainly in the coenosteum. Columella small, substyliform. Endotheca unknown. Wall generally compact, septoparathecal. Coenosteum broad, consisting of isolated trabeculae and tabulae. Budding extracalicinal.
Material. ERNO L-7003.
Discussion. The only sample we have is too small to prepare a longitudinal thin section. It clearly differs from Multicolumastrea cyathiformis on account of its smaller calicular dimensions.
Occurrence. Maastrichtian of Mexico (Chiapas), roadcut north of Ocozocoautla.
DISCUSSION
The fauna is dominated by Late Cretaceous genera. Only Astraeofungia has been known since the Late Jurassic, Columactinastrea since the Early Cretaceus, Actinacis since the Late Albian and Pachygyra since the Cenomanian. One genus (Actinastrea) appeared after the Cenomanian/ Turonian boundary, but the remaining genera are restricted to the Maastrichtian and are not known in older sediments. This includes Filkornia, Multicolumnastraea, very probably also Favioseris (its distribution in the Campanian is questionable), and the material without a generic name, which is assigned here to Microsolena and to the Fungiidae.
At family level there are no first occurrences in the Maastrichtian for the fauna studied. The Felixaraeidae have been known since the Late Turonian (with Felixaraea), the Pachyphylliidae (with Pachyphyllia Alloiteau, 1957) and Fungiidae (with Pseudocunnolites Reig Oriol, 1988) since the Santonian. All other families have been known since the Late Jurassic (Actinastraeidae, Leptophylliidae, Microsolenidae) or late Early Cretaceous (Actinacicidae, Phyllosmilliidae). But the Late Cretaceous family nevertheless diversified in the Maastrichtian.
Nine out of the twelve species were found in other localities (Figure 7). With one exception (Astraeofungia felixi) these species are only known from the Late Cretaceous, and four are only known from the Maastrichtian. This confirms the special character of Maastrichtian coral faunas. There are Maastrichtian coral faunas (exclusively from the Caribbean, e.g. Jamaica and NE Mexico) which share species with the Ocozocoautla fauna. Faunas from the Western or Central Tethys share a few cosmopolitan genera (Actinastrea, Columactinastraea, Astraeofungia).
The above-mentioned exclusive Maastrichtian genera are restricted to the Caribbean. The closing of the Tethys and the opening of the Atlantic Ocean caused the flow of the oceanic water masses to change from west-east to north- south. Even in the Maastrichtian corals showed an increasing provincialism, i.e. a provincialism that did not just emerge when the isthmus between North and South America was beginning to close (see also Coates, 1973).
ACKNOWLEDGEMENTS
I am very grateful to the people who granted me access to the fossil collections under their care, namely to María del Carmen Perrilliat (IGL, Mexico City, Mexico), Jacob Leloux (NNM, Leiden, The Netherlands), Jessica Cundiff (MCZ, Cambridge, Mass., USA), Jill Darrell (NHM, London, England), René Fraaije (DGP, Boxtel, The Netherlands), Sandra Kaiser (IPB, Bonn, Germany), Sebastián Calzada (MGSB, Barcelona, Spain), and Stephen Cairns (USNM, Washington D.C., USA). I am much indebted to Simon Mitchell (Kingston) for passing on to me his insights into the outcrop situation in Jamaica. Fieldwork was supported by Javier Avendaño and José Juan Jiménez. Much of the systematic measuring of the calicular dimensions of the corals was done by L. Puebla Clark (Hermosillo). The Universidad Nacional Autónoma de México (DGAPA) financially supported my work in collections in Europe through a PASPA programme. Insightful reviews by Boguslaw Kolodziej (Kraków) and Thomas Stemann (Kingston) helped to improve the study. For grammatical correction I would like to thank Gertraud Moss (Dresden, Germany).
REFERENCES
Alloiteau, J., 1936, Polypiers fossiles de Madagascar. 1: Formes du Crétacé de la province d'Ananalava: Annales géologiques du Service des Mines de Madagascar, 6, 41-53. [ Links ]
Alloiteau, J., 1952, Embranchement des coelentérés, in Piveteau, J. (ed.), Traité de Paléontologie: Paris, Masson, 376-684. [ Links ]
Alloiteau, J., 1954, Le genre Actinastrea d'Orbigny 1848 dans le Crétacé supérieur francais: Annales Hebert et Haug, 4, 1-104. [ Links ]
Alloiteau, J., 1957, Contribution à la systématique des Madréporaires fossiles: Paris, Centre National de la Recherche Scientifique, 462 pp. [ Links ]
Alloiteau, J., 1958, Monographie des Madréporaires fossiles de Madagascar: Annales géologiques de Madagascar, 25, 1-218. [ Links ]
Baron-Szabo, R.C., 1998, A new coral fauna from the Campanian of Northern Spain (Torallola village, Prov. Lleida): Geologische und Paläontologische Mitteilungen, 23, 127-191. [ Links ]
Baron-Szabo, R.C., 2000, Late Campanian-Maastrichtian corals from the United Arab Emirates-Oman border region: Bulletin of the Natural History Museum London (Geology), 56(2), 91-131. [ Links ]
Baron-Szabo, R.C., 2003, Taxonomie und Ontogenie von scleractinen Korallen der ostalpinen Oberkreide (Hochmoos-und Grabenbachschichten, Gosau-Gruppe, Santon): Jahrbuch der Geologischen Bundesanstalt, 143(2), 107-201. [ Links ]
Baron-Szabo, R.C., 2006, Corals of the K/T-boundary: Scleractinian corals of the Suborders Astrocoeniina, Faviina, Rhipidogyrina, and Amphiastraeina: Journal of Systematic Palaeontology, 4, 1-108. [ Links ]
Baron-Szabo, R.C., 2008, Corals of the K/T-boundary: Scleractinian corals of the suborders Dendrophylliina, Caryophylliina, Fungiina, Microsolenina, and Stylinina: Zootaxa, 1952, 1-244. [ Links ]
Baron-Szabo, R.C., Schafhauser, A., Götz, S., Stinnesbeck, W., 2006, Scleractinian corals from the Cárdenas Formation (Maastrichtian), San Luis Potosí, Mexico: Journal of Paleontology, 80(6), 1033-1046. [ Links ]
Bataller, J., 1936, Contribución al estudio de los poliperos Cretácicos de Cataluña: Ibérica, 1103, 38-46. [ Links ]
Bataller, J., 1937, La fauna coral-lina del Cretàcic de Catalunya i regions limítrofes: Arxius de l'escola superior d'agricultura, (N.S.), 3(1), 1-299. [ Links ]
Beauvais, M., 1982, Révision Systématique des Madréporaires des couches de Gosau: Paris, Comptoir géologique, 5 vols. [ Links ]
Beauvais, L., Beauvais, M., 1975, Une nouvelle famille dans le sous-ordre des Stylinida Alloiteau: les Agatheliidae nov. fam. (Madréporaires mésozoïques): Bulletin de la Société géologique de France, (7), 17, 576-581. [ Links ]
Blainville, H.M., 1830, Zoophytes, in Defrance, J.L.M. (ed.), Dictionnaire des sciences naturelles: Paris, Levrault, 60, 1-548. [ Links ]
Coates, A.G., 1973, Cretaceous tethyan coral-rudist biogeography related to the Evolution of the Atlantic Ocean: Special Papers in Palaeontology, 12, 169-174. [ Links ]
Dana, J.D., 1846-1849, Zoophytes. United States Exploring Expedition during the years 1838, 1839, 1840, 1841, 1842 under the command of Charles Wilkes, U. S. N.: Philadelphia, Lea and Blanchard, 121-708. [ Links ]
Defrance, J.L.M., 1828, Turbinolie, in Defrance, J.L.M. (ed.), Dictionnaire des sciences naturelles: Paris, Levrault, 56, 91-94. [ Links ]
de Fromentel, E., 1862, Zoophytes, terrain crétacé (1): Paléontologie française (A. d'Orbigny, ed.), 8, 1-48. [ Links ]
d'Orbigny, A., 1849, Note sur les polypiers fossiles: Paris, Masson, 12 pp. [ Links ]
Duncan, P.M., 1880, Tertiary and Upper Cretaceous fauna of Western India (1:) Sind fossil corals and Alcyonaria: Memoirs of the Geological Survey of India, Palaeontologia Indica, (14), 1(2), 1-110. [ Links ]
Duncan, P.M., 1884, A revision of the families and genera of the sclerodermic Zoantharia Edwards et Haime, or Madreporaria (M. Rugosa excepted) (1-4): Journal of the Linnean Society of London, Zoology, 18, 1-204. [ Links ]
Duncan, P.M., Wall, G.P., 1865, A notice of the geology of Jamaica, with special reference to the district of Clarendon; with descriptions of the Cretaceous, Eocene and Miocene corals of the islands: Quarterly Journal of the Geological Society of London, 21, 1-14. [ Links ]
Felix, J., 1903, Studien über die korallenführenden Schichten der oberen Kreideformation in den Alpen und den Mediterrangebieten (1) Die Anthozoën der Gosauschichten in den Ostalpen: Palaeontographica, 49, 163-360. [ Links ]
Filkorn, H.F., Avendaño-Gil, J., Coutiño-José, M.A., Vega-Vera, F.J., 2005, Corals from the Upper Cretaceous (Maastrichtian) Ocozocoautla Formation, Chiapas, Mexico: Revista Mexicana de Ciencias Geológicas, 22(1), 115-128. [ Links ]
Goldfuss, A., 1826, Petrefacta Germaniae (1,1): Düsseldorf, Arnz, 76 pp. [ Links ]
Goldfuss, A., 1829, Petrefacta Germaniae (1,2): Düsseldorf, Arnz, 87 pp. [ Links ]
Hardenbol, J., Thierry, J., Farley, M.B., Jacquin, T., Graciansky, P.C. de, 1998, Cretaceous biostratigraphy, in Graciansky, P.C. de, Hardenbol, J., Jacquin, T., Vail. P. (eds.), Mesozoic Cenozoic Sequence Stratigraphy of European Basins: Society of Economic Paleontologists and Mineralogists (SEPM), Special Publication, 60, 4-13, 763-781. [ Links ]
Khazaei, A.R., Skelton, P. W., Yazdi, M., 2010, Maastrichtian rudist fauna from Tarbur Formation (Zagros region, SW Iran): preliminary observations: Turkish Journal of Earth Sciences, 19, 703-719. [ Links ]
Kühn, O., 1933, Das Becken von Isfahan-Saidabad und seine altmiozäne Korallenfauna: Palaeontographica, (A), 79(1/2), 143-218. [ Links ]
Lamarck, J.B.P. de, 1801, Système des animaux sans vertèbres: Paris, Comptoir géologique, 432 pp. [ Links ]
Lamouroux, J.V.F., 1821, Exposition méthodique des genres de l'ordre des polypiers: Paris, Agasse, 115 pp. [ Links ]
Leloux, J., 2003, Columactinastraea anthonii sp. nov. (Scleractinia, Astrocoeniina), a new coral species from the Maastrichtian (Upper Cretaceous) of The Netherlands: Scripta Geologica, 126, 185-201. [ Links ]
Leloux, J., 2004, Notes on taxonomy and taphonomy of two Upper Maastrichtian (Upper Cretaceous) scleractinian corals from Limburg, The Netherlands: Scripta Geologica, 127, 313-339. [ Links ]
Lesauvage, M., 1823, Mémoire sur un genre nouveau de polypier fossile: Mémoires de la Société d'Histoire Naturelle de Paris, 1, 241-244. [ Links ]
Löser, H., 1998, Lower Campanian corals from Amasya (Turkey): Abhandlungen und Berichte für Naturkunde und Vorgeschichte, 20, 77-87. [ Links ]
Löser, H., 2005, Stratigraphy of Cretaceous coral genera: Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 238, 231-277. [ Links ]
Löser, H., 2011, Systematic revision of the Placocoeniidae (Scleractinia; Late Cretaceous): Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 261, 195-200. [ Links ]
Löser, H., 2012, Revision of Actinastrea, the most common Cretaceous coral genus: Paläontologische Zeitschrift, 86, 15-22. [ Links ]
Löser, H., 2012, Intraspecific variation in the genus Stelidioseris (family Actinastraeidae, suborder Archeocaeniina, order Scleractinia; Jurassic-Cretaceous): Geologica Belgica, 15(4), 382-387. [ Links ]
Löser H., Barattolo F., Calzada Badia S., Chikhi-Aouimeur F., Dhondt A., Erlich, R.N., Fözy, I., Geister J., Hiss, M., Kolodziej, B., Leloux, J., Lewy, Z., Madhavaraju, J., Minor, K.P., Mitchell, S., Moosleitner, G., Niebuhr, B., Peza, L., Remane, J., Romana, R., Sanders, D., Sikharulidze, G.Y., Sinnyovski, D., Steuber, T., Tröger, K.-A., Turnsek, D., Vecchio, E., Vilella i Puig, J. & Zítt, J., 2005, List of Localities: Catalogue of Cretaceous Corals, 3, 1-366. [ Links ]
Löser, H., Castro, J.M., Nieto, L.M., 2010, A small Albian coral fauna from the Sierra de Seguilí (Alicante province, SE Spain): Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 255, 315-326. [ Links ]
Matteucci, R., Schiavinotto, F., Sirna, G., Russo, A., 1982, Palaeoenvironmental significance of Maastrichtian biological communities in the Pachino area (Sicily) and preliminary data on their distribution in the Mediterranean Upper Cretaceous, in Palaeontology, Essential of Historical Geology: Modena, S.T.E.M. Mucchi, 77-96. [ Links ]
Matthews, S.C., 1973, Notes on open nomenclature and on synonymy lists: Palaeontology, 16(4), 713-719. [ Links ]
Michelin, H., 1841, Iconographie zoophytologique. Description par localités et terrains des polypiers fossiles de France (1): Paris, Bertrand, 1-40. [ Links ]
Milne Edwards, H., Haime, J., 1848a, Observations sur les polypiers de la famille des astréides: Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 27, 465-469. [ Links ]
Milne Edwards, H., Haime, J., 1848b, Note sur la classification de la deuxième tribu de la famille des astréides: Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 27, 490-497. [ Links ]
Milne Edwards, H., Haime, J., 1849, Mémoire sur les polypiers appartenant à la famille des oculinides, au groupe intermédiaire des Pseudoastréides et à la famille des Fongides: Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 29, 67-73. [ Links ]
Morycowa, E., Roniewicz, E., 1995, Microstructural disparity between Recent fungiine and Mesozoic microsolenine scleractinians: Acta Palaeontologica Polonica, 40(4), 361-385. [ Links ]
Odin, G.S., 1996, Definition of a global boundary stratotype section and point for the Campanian/Maastrichtian boundary: Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Science de la Terre, 66 suppl., 111-117. [ Links ]
Oppenheim, L.P., 1930, Die Anthozoen der Gosauschichten in den Ostalpen: Berlin, privately published, 604 pp. [ Links ]
Pfister, T., 1980, Systematische und paläoökologische Untersuchungen an oligozänen Korallen der Umgebung von San Luca: Schweizerische Paläontologische Abhandlungen, 103, 1-121. [ Links ]
Pratz, E., 1882-1983, Über die verwandschaftlichen Beziehungen einiger Korallengattungen mit hauptsächlicher Berücksichtigung ihrer Septalstructur: Palaeontographica, 29, 81-124. [ Links ]
Reig Oriol, J., 1987, Revisión y validez del género Anisoria (Escleractinia Cretácica): Trabajos del Museo Geológico del Seminario C. de Barcelona, 222, 3-9. [ Links ]
Reig Oriol, J., 1988, Dos nuevos géneros de corales cretácicos: Batalleria, 1, 39-45. [ Links ]
Reig Oriol, J., 1989, Sobre varios géneros y especies de escleractinias fósiles del Cretácico Catalán: Barcelona, privately published, 69 pp. [ Links ]
Reig Oriol, J., 1992, Madreporarios cretácicos de España y Francia: Barcelona, privately published, 48 pp. [ Links ]
Reig Oriol, J., Vilella, J., 1995, Un nuevo subgénero de corales (Maastrichtiense de Isona, Llerida): Batalleria, 5, 37-39. [ Links ]
Reyeros-Navarro, M.M., 1963, Corales del Cretácico inferior de San Juan Raya, Estado de Puebla: Paleontologia Mexicana, 17, 1-21. [ Links ]
Reyeros de Castillo, M.M., 1983, Corales de algunas formaciones cretácicas del estado de Oaxaca: Paleontología Mexicana, 47, 1-67. [ Links ]
Swinburne, N.H.M., Bilotte, M., Pamouktchiev, A., 1992, The stratigraphy of the Campanian-Maastrichtian rudist beds of Bulgaria and a reassessment of the range of the genus Pironaea: Cretaceous Research, 13, 191-205. [ Links ]
Umbgrove, J.H.F., 1925, Die Anthozoa uit het Maastrichtsche tufkrit. [The Anthozoa of the Maastrichtian tufaceous chalk.]: Leidse geologische mededelingen, 1(1), 83-126. [ Links ]
Vaughan, T.W., 1899, Some Cretaceous and Eocene corals from Jamaica: Bulletin of the Museum of Comparative Zoology, 34(1), 227-250. [ Links ]
Vaughan, T.W., 1905, A critical review of the literature of the simple genera of Fungida, with a tentative classification: Proceedings of the United States National Museum, 28(1401), 371-424. [ Links ]
Vaughan, T.W., Wells, J.W., 1943, Revision of the suborders, families and genera of scleractinia: Geological Society of America, Special Paper 44, 1-363. [ Links ]
Vega, F.J., Feldmann, R.M., García Barrera, P., Filkorn, H.F., Pimentel, F., Avendaño, J., 2001, Maastrichtian Crustacea (Brachyura: Decapoda) from the Ocozocuautla Formation in Chiapas, southeast Mexico: Journal of Paleontology, 75(2), 319-329. [ Links ]
Veron, J.E.N., 1995, Corals in space and time. The biogeography and evolution of the Scleractinia: Sydney, UNSW Press, 321 pp. [ Links ]
Verrill, A.E., 1868-1870, Notes on the Radiata in the museum of Yale College, with descriptions of new genera and species. No. 6. Review of the corals and polyps of the West Coast of America: Transactions of the Connecticut Academy of Arts and Sciences, 1(2), 377-567. [ Links ]
Vidal, L.M., 1917, Nota paleontológica sobre el cretácico de Cataluña: Congreso de la Asociación española para el progreso de las ciencias, 3-19. [ Links ]
Vidal, L.M., 1921, Contribución a la Paleontología del Cretácico de Cataluña: Memorias de la Real Academia de Ciencias y Artes de Barcelona, (3), 17(2), 89-107. [ Links ]
Wells, J.W., 1933, Corals of the Cretaceous of the Atlantic and Gulf Coastal Plains and Western Interior of the United States: Bulletins of American Paleontology, 18, 83-292. [ Links ]
Wells, J.W., 1934, Some fossil corals from West Indies: Proceedings of the United States National Museum, 83, 71-110. [ Links ]
Wells, J.W., 1935, Corals from the Cretaceous and Eocene of Jamaica: Annals and Magazine of natural History, Series 10, Volume 15, Issue 86, 183-194. [ Links ]

