










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.27 no.1 Ciudad de México abr. 2010
Los Glyptodontinae (Xenarthra, Glyptodontidae) del piso/edad Chapadmalalense (Plioceno tardío): revisión y aportes a su conocimiento
The Glyptodontinae (Xenarthra, Glyptodontidae) of the Chapadmalalan stage/age (late Pliocene): review and contributions to knowledge
Cristian Oliva1,*, Alfredo Eduardo Zurita2, Alejandro Dondas3 y Gustavo Juan Scillato–Yané4
1 Museo Municipal de Ciencias Naturales Carlos Darwin, Urquiza 123, Punta Alta, Provincia de Buenos Aires, Argentina. *E–mail: cristianoliva78@yahoo.com.ar.
2 Centro de Ecología Aplicada del Litoral (CECOAL–CONICET) y Universidad Nacional del Nordeste, Ruta 5, Km 2.5, Corrientes, Provincia de Corrientes, Argentina.
3 Museo Municipal de Ciencias Naturales Lorenzo Scaglia, Av. Libertad 3099, Plaza España, Mar del Plata, Provincia de Buenos Aires, Argentina.
4 Departamento Científico Paleontología de Vertebrados, Museo de La Plata, CONICET, Paseo del Bosque s/n B1900FWA La Plata, Argentina.
Manuscript received: April 4, 2009.
Corrected manuscript received: Octoober 8, 2009.
Manuscript accepted: November 3, 2009.
RESUMEN
Los primeros registros de Glyptodontidae Glyptodontinae corresponden al Mioceno y Plioceno temprano del extremo norte de América del Sur (Boreostemma spp.), mientras que al sur de este continente (Argentina) los hallazgos más antiguos se remontan al "Araucanense" (Mioceno tardío – Plioceno temprano) (Glyptodontidium tuberifer Cabrera). En este contexto, las formas terciarias de Glyptodontinae de la parte austral de América del Sur ("Araucanense"–Marplatense) eran poco conocidas, y estaban mayormente limitadas a fragmentos de la coraza dorsal. Más precisamente, en el Chapadmalalense superior (Plioceno tardío) se reconocía la existencia de dos especies: Paraglyptodon dubius Castellanos y P. chapalmalensis (Ameghino in Rovereto), esta última fósil guía de este Piso/Edad, mientras que el "Mesopotamiense" (Huayqueriense–Montehermosense?) (Mioceno tardío–Plioceno temprano) se caracterizaba por la especie P. paranensis Castellanos. La revisión sistemática aquí efectuada sugiere que, para el Piso/Edad Chapadmalalense superior, la única especie válida es P. chapalmalensis, en tanto que P. dubius es un sinónimo de ésta. A su vez, P. paranensis, ("Mesopotamiense"), limitado a un osteodermo mal preservado, debe ser considerado como un nomen vanum. Por último, damos a conocer el registro craneano más antiguo y completo conocido de un representante de esta subfamilia para el sur de América del Sur, tentativamente asignado a cf. Paraglyptodon chapalmalensis. Entre los principales caracteres observados sobre este nuevo material se destacan: a) todos los molariformes superiores claramente trilobulados, a excepción del M1; b) narinas de contorno subtrapezoidal; c) órbitas abiertas posteriormente, y en comunicación con la fosa temporal; d) área del tubérculo lacrimal y escotaduras orbitarias de morfología muy similar a las de Glyptodon y Paraglyptodon uquiensis; e) cráneo abruptamente truncado por el acortamiento de los nasales y premaxilares, con un perfil dorsal relativamente recto e inclinado hacia delante.
Palabras clave: Glyptodontidae, Glyptodontinae, Paraglyptodon chapalmalensis (Ameghino in Rovereto), Chapadmalalense, Argentina, América del Sur.
ABSTRACT
The first records of Glyptodontinae Glyptodontidae come from the Miocene and Pliocene of northern South America (Boreostemma spp.), while the oldest records in southern South America (Argentina) correspond to the "Araucanian" (late Miocene–early Pliocene) (Glyptodontidium tuberifer Cabrera). In this context, the Tertiary taxa of Glyptodontinae from southern South America ("Araucanian"–Marplatan) were scarcely known, and they were limited to a few fragments of the dorsal carapace. More precisely, in the Upper Chapadmalalan Stage/Age (late Pliocene) it is possible to recognise two species: Paraglyptodon dubius Castellanos and P. chapalmalensis (Ameghino in Rovereto); the latter is a guide fossil for this lapse. On the other hand, the "Mesopotamian" (Huayquerian–Montehermosan?; late Miocene–early Pliocene) was characterized by the presence of P. paranensis Castellanos. In this context, the systematic revision suggests that, in the Upper Chapadmalalan, the only valid species is P. chapalmalensis, while P. dubius is a synonym of this. In turn, P. paranensis, ("Mesopotamian"), a taxon represented by a very bad preserved osteoderm of the dorsal carapace, must be considered as a nomen vanum. Finally, we report and describe in this paper the oldest and most complete Tertiary skull of a southern South America Glyptodontinae, here tentatively classified as cf. Paraglyptodon chapalmalensis. Among the most interesting observed characters in this new material it stand out: a) all uper molarifoms (except M1) clearly trilobated; b) narines showing a subtrapezoidal contour; c) the orbits are posteriorly open and freely communicated with the temporal fossae; d) lacrimal tubercle area and orbital notch very similar to that observed in Glyptodon and Paraglyptodon uquiensis; e) skull abruptly truncated by the shortening of the nasals and premaxillae, with a dorsal profile relatively straight and inclined forward.
Key words: Glyptodontidae, Glyptodontinae, Paraglyptodon chapalmalensis (Ameghino in Rovereto),Chapadmalalan, Argentina, South America.
INTRODUCCIÓN
Entre la gran diversidad de xenartros Cingulata, los Glyptodontidae constituyen unos de los clados más diversificados y mejor representados, habiendo sido reconocidos más de 65 géneros para esta familia (McKenna y Bell, 1997).
De las cinco subfamilias existentes hasta el momento (Glyptatelinae Castellanos,1932; Propalaehoplophorinae Ameghino, 1891; Glyptodontinae Gray, 1869; Hoplophorinae Huxley, 1864; y Doedicurinae Ameghino, 1889), sólo los gliptodontinos (y probablemente los gliptatelinos) han participado del Gran Intercambio Biótico Americano (GIBA; ver Webb, 2006; Carlini et al., 2008a).
En este marco, la filogenia y paleobiogeografia de los Glyptodontidae Glyptodontinae han sido recientemente reinterpretadas, en tanto sus primeros registros (que probablemente revelen los estadios iniciales de su cladogénesis) provienen del Mioceno medio–Plioceno del extremo norte de América del Sur (Venezuela y Colombia), siendo asignados a un nuevo género: Boreostemma Carlini et al. (2008b).
Hasta el presente es posible reconocer cuatro especies para este taxón: B. venezolensis (Simpson, 1947) (Mioceno medio? de Venezuela); B. acostae (Villaroel, 1983) y B. gigantea (Carlini et al., 1997) (Mioceno medio de La Venta, Colombia); B. pliocena Carlini et al., 2008b (Plioceno de Venezuela).
A diferencia de lo que ocurre con las formas boreales (Boreostemma Carlini et al. y Glyptotherium Osborn, 1903) el conocimiento que se tiene de los gliptodontinos terciarios (Mioceno–Plioceno) del extremo sur de América del Sur es escaso (Carlini et. al., 2008b); en este sentido, resulta claramente necesaria una revisión sistemática moderna del grupo.
El registro más antiguo de Glyptodontinae para el extremo austral de Sudamérica corresponde a Glyptodontidium tuberifer Cabrera, 1944, un taxón pobremente representado del "Araucanense" (Mioceno tardío – Plioceno temprano) de las provincias de Catamarca y Tucumán, Argentina (Cabrera, 1944). Desde una perspectiva paleobiogeográfica, este hallazgo coincide con el acmé de la "Edad de las Planicies Australes", situación que debe haber facilitado el proceso de migración de los gliptodontinos hacia ámbitos australes (Ortiz–Jaureguizar y Cladera, 2006; Zurita et al., 2008a).
Para el "Conglomerado osífero de la Formación Ituzaingó" ["Mesopotamiense" (Mioceno tardío – Plioceno temprano)] de los alrededores de la ciudad de Paraná, provincia de Entre Ríos, Argentina, ha sido descrita la especie Paraglyptodon paranensis Castellanos, 1953, sólo conocida con base en un osteodermo aislado y mal preservado de la coraza dorsal (Ameghino, 1889: pl. LIV, fig. 4), cuya pertenencia a un Glyptodontinae es sumamente cuestionable (ver conclusión).
Luego de una prolongada ausencia durante el Montehermosense (Mioceno tardío–Plioceno temprano) (ca. 6.8–3.9 Ma), es posible reconocer la presencia de la subfamilia en el Chapadmalalense de la costa atlántica de la provincia de Buenos Aires (Argentina) con Paraglyptodon chapalmalensis (Ameghino in Rovereto, 1914). Este taxón tiene importancia bioestratigráfica puesto que caracteriza el Piso/Edad Chapadmalalense superior (ca. 3.5–3.2 Ma) (ver Cione y Tonni, 1995, 1999, 2005). Se encuentra limitado hasta el momento al material tipo (MACN 6162), consistente en un fragmento de coraza dorsal, proveniente de los niveles superiores de la Formación Chapadmalal, en los alrededores de la localidad de Mar del Plata.
Otra especie registrada para la misma región y unidad litoestratigráfica que P. chapalmalensis es P. dubius Castellanos, 1953, establecida con base en dos osteodermos articulados (MACN 6285) originalmente clasificados por Ameghino (1908) y Rovereto (1914) como "Sclerocalyptus" sp. y "Sclerocalyptus" chapalmalensis, respectivamente.
El registro fósil de Glyptodontinae en el Marplatense (Plioceno tardío–Pleistoceno temprano) (ca. 3.2–1.8 Ma) de la Región Pampeana es escaso, tal como ocurre con otros grupos de gliptodontes. Sin embargo, para el "Uquiense" (Plioceno) de la provincia de Jujuy (Argentina), Castellanos (1953) describió Paraglyptodon uquiensis Castellanos, 1953, conocido por un cráneo parcialmente completo, la porción proximal del fémur izquierdo y algunos osteodermos de la coraza dorsal provenientes de sedimentitas de la Formación Uquía (ver Reguero et al., 2007).
Durante el Pleistoceno (ca. 2.6–0.0011 Ma), los Glyptodontinae australes experimentan una importante diversificación con el género Glyptodon Owen, 1838. La especie más antigua es Glyptodon munizi Ameghino, 1881 del Ensenadense (Pleistoceno temprano–medio) (ca. 1.07 – 0.98) de la localidades de Mar del Plata, San Pedro y las "Toscas del Río de La Plata" (provincia de Buenos Aires) (ver Soibelzon et al., 2006, 2008). En el Bonaerense (Pleistoceno medio) (ca. 0.5–0.13 Ma) y Lujanense (Pleistoceno tardío – Holoceno temprano) (ca. 0.13–0.0085 Ma) este gliptodontino se convierte en el Glyptodontidae con mayor distribución latitudinal (Zurita et al., 2009).
La presente contribución tiene como principales objetivos: a) dar a conocer un nuevo y significativo material craneano asignado a la subfamilia Glyptodontinae para el Piso/Edad Chapalmalalense superior, y b) discutir la validez de las especies reconocidas para el lapso "Mesopotamiense"–Chapadmalalense (Mioceno tardío–Plioceno tardío) en la Argentina.
Abreviaturas
MACN: Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (Buenos Aires, Argentina); MCN: Museo de Ciencias Naturales de Caracas (Venezuela); MLP: Facultad de Ciencias Naturales y Museo de La Plata (La Plata, Argentina); MMP: Museo Municipal de Ciencias Naturales "Lorenzo Scaglia" (Mar del Plata, Argentina); UMMP: University of Michigan Museum of Paleontology (USA); USNM: collections of the former United State National Museum in the National Museum of Natural History, Smithsonian Institution; AMNH (F: AM): American Museum of Natural History, Department of Vertebrate Paleontology and Frick Collection (USA).
PALEONTOLOGÍA SISTEMÁTICA
Superorden Xenarthra Cope, 1889
Orden Cingulata Illiger, 1811
Familia Glyptodontidae Gray, 1869
Subfamilia Glyptodontinae Gray, 1869
Género Paraglyptodon Castellanos, 1932
Glyptodon Owen (Ameghino, 1908).
Sclerocalyptus Ameghino (Ameghino partim. 1908; Rovereto, 1914, partim).
Especie tipo. Glyptodon chapalmalensis Ameghino, 1908.
Paraglyptodon chapalmalensis
(Ameghino in Rovereto, 1914: 206, lám. 27, fig. 1)
Figuras 2a y 2f
Sclerocalyptus sp. Ameghino, 1908.
Sclerocalyptus chapalmalensis Ameghino in Rovereto, 1914.
Paraglyptodon dubius Castellanos, 1953.
Paraglyptodon chapalmalensis (Ameghino) Pascual et al., 1966.
Holotipo. MACN 6162, fragmento compuesto por 16 osteodermos correspondientes a la región lateral y latero–dorsal de la coraza (Rovereto, 1914: pl. XXVII; fig. 1; Pascual et al., 1966: pl. XLIII; fig. B) (Figura 2a).
Procedencia estratigráfica y geográfica. Sección cuspidal de la Formación Chapadmalal (Plioceno superior). Mar del Plata, provincia de Buenos Aires, Argentina (Figura 1).
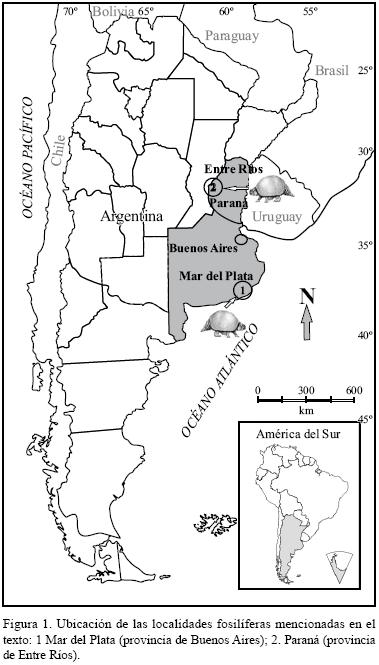
Material referido. MACN 6285 (Holotipo de Paraglyptodon dubius Castellanos, 1953), dos osteodermos articulados pertenecientes a la región lateral de la coraza (Rovereto, 1914: pl. XXIX, fig. 2; Pascual et al., 1966: pl. XLIII, fig. C) (Figura 2f).
Procedencia geográfica y estratigráfica. Sección cuspidal de la Formación Chapadmalal (Plioceno superior). Mar del Plata, provincia de Buenos Aires, Argentina (Figura 1).
Descripción. Los osteodermos de P. chapalmalensis (MACN 6162, MACN 6285) poseen de cuatro a seis lados; los mayores miden 43 mm de diámetro antero–posterior, por 32 mm de diámetro dorso–ventral y 18 mm de espesor, en tanto que los más pequeños tienen 34 mm, 31 mm y 18 mm de diámetro antero–posterior, diámetro dorso–ventral y espesor respectivamente. La superficie expuesta de cada uno de ellos es claramente más rugosa y punteada que en Boreostemma y Glyptodontidium, aunque no tan marcadamente como en las formas terminales de la subfamilia (P. uquiensis y Glyptodon spp.). La figura central es subcircular, plana a ligeramente cóncava, siendo en todos los casos de mayor tamaño y estando sobreelevada respecto de las figuritas periféricas. Éstas últimas, se hallan presentes en número de cuatro a cinco en MACN 6285 y de seis a ocho en MACN 6162, formando figuras en común con las de placas adyacentes en los cuatro márgenes disponibles del osteodermo; son pequeñas, irregulares y mal delimitadas, debido a la presencia de surcos radiales poco definidos. Éstos y el surco anular son poco profundos pero considerablemente amplios; en este contexto son más anchos que en Glyptodontidium tuberifer pero más angostos que en Glyptodon. En la intersección del surco central con los periféricos existen algunos forámenes pilíferos de reducido diámetro.
Comentarios taxonómicos. P. chapalmalensis fue originalmente reconocido por Ameghino (1908: 426) como perteneciente al género pleistoceno Glyptodon (G. chapalmalensis), asumiendo correctamente que se trataba de un Glyptodontinae y no de un Hoplophorinae. En esa oportunidad, Ameghino (1908) no llevó a cabo ninguna descripción y/o diagnosis del mismo, por lo que debe considerarse como un nomen nudum (ver Mones, 1986). Poco después, Rovereto (1914: lám. 27, fig. 1) figuró el material en cuestión. En este contexto, la especie es válida a partir de Rovereto, pero como éste la atribuyó a Ameghino, la notación debe ser "Glyptodon" chapalmalensis Ameghino in Rovereto, 1914: 206, 245, lám. 27 (fig. 1) (ICZN, Art. 50). Esto se debe a que el ICZN (Art. 12.2.7.) establece que una especie ilustrada sin descripción antes de 1930 es válida. Posteriormente, Castellanos (1932) asignó este taxón al género Paraglyptodon Castellanos (P. chapalmalensis) y describió por primera vez el material.
El análisis efectuado en la presente revisión demuestra que se trata efectivamente de un Glyptodontinae. El surco central y los radiales son amplios y tienen su sección en "U"; a esto debe sumarse que la superficie expuesta de cada osteodermo es extremadamente rugosa, sin figuritas periféricas accesorias. Estas características son consideradas como sinapomorfías de la subfamilia (ver Carlini et al., 2008b).
Un estudio comparado del holotipo (MACN 6162) con el género Glyptodon y las formas terciarias G. tuberifer (MLP 29–X–8–3) y P. uquiensis (MACN 5330 y 5544) indica que existe una clara diferenciación morfológica entre todos ellos y que, en consecuencia, P. chapalmalensis es una especie válida, definida por buenos caracteres diagnósticos. Sus osteodermos presentan un grado de evolución en su cara expuesta (grado de rugosidad y morfología de los surcos) intermedio entre aquellos de G. tuberifer ("Araucanense") y P. uquiensis ("Uquiense"), que es congruente con la antigüedad del taxón (Chapadmalalense superior). Adicionalmente, en P. chapalmalensis la superficie expuesta de las osteodermos dorsales es más rugosa y los surcos centrales y radiales se encuentran más definidos y son proporcionalmente más anchos que aquellos de Boreostemma , Glyptodontidium, y Glyptotherium , no alcanzando, sin embargo, el grado de desarrollo observado en P. uquiensis y las especies conocidas del género Glyptodon.
Por su parte, el material MACN 6285 fue originalmente incluido por Ameghino (1908: 246) en un listado sistemático bajo la denominación de "Sclerocalyptus" sp., y luego reconocido como "Sclerocalyptus" chapalmalensis por Rovereto (1914: 206; pl. XXIX, fig. 2). Curiosamente, y a pesar de la notable similitud entre estos osteodermos y aquellos de P. chapalmalensis (= "Glyptodon" chapalmalensis) y poseer la misma procedencia geográfica y estratigráfica, estos autores los consideraron como pertenecientes a un Glyptodontidae Hoplophorinae (= Sclerocalyptinae). Correctamente, Castellanos (1953: 28) interpretó estos restos como correspondientes a un Glyptodontinae, incluyéndolos en el género Paraglyptodon, y reconociendo la especie P. dubius, dado que el epíteto específico "chapalmalensis" se encontraba pre–ocupado. Más tarde, Pascual et al. (1966: pl. XLIII, fig. C) asignaron implícitamente dicho material a P. chapalmalensis, sin agregar mayor evidencia al momento de sustentar esta clasificación taxonómica.
El estudio comparado aquí efectuado indica que los caracteres observados por Castellanos (1953) para P. dubius (e.g. "...figuritas periféricas mal delimitadas... superficie poco rugosa... escultura poco precisa... surcos mal delimitados") son consecuencia de la erosión parcial que ha sufrido la superficie expuesta de los osteodermos; sin embargo, las partes bien preservadas no muestran ninguna diferencia significativa respecto de aquellas homólogas de P. chapalmalensis, por lo que consideramos este material como perteneciente a esta especie, coincidiendo con la interpretación de Pascual et al. (1966). Adicionalmente, resulta destacable que ambos materiales tienen la misma procedencia geográfica y estratigráfica.
cf. Paraglyptodon chapalmalensis
Material referido. MMP 4676, cráneo en buen estado de conservación que no preserva el arco cigomático del lado izquierdo, el proceso descendente del maxilar derecho y gran parte de la región dorsal de los nasales, de los frontales y del basioccipital.
Procedencia geográfica y estratigráfica. Sección media–cuspidal de la Formación Chapadmalal (Plioceno tardío). Mar del Plata, provincia de Buenos Aires, Argentina (Figura 1).
Descripción. El espécimen considerado exhibe una serie de caracteres craneanos combinados que sugieren su asignación a la subfamilia Glyptodontinae (Hoffstetter, 1958; Gillette y Ray, 1981): a) el cráneo está abruptamente truncado por el acortamiento de los nasales y premaxilares y su perfil dorsal es relativamente recto e inclinado hacia adelante; b) la apertura nasal es de contorno subtrapezoidal; c) las órbitas están abiertas posteriormente, comunicándose libremente con la fosa temporal; d) las áreas del tubérculo lacrimal y escotaduras orbitarias muestran una morfología muy similar a las de Glyptodon. Adicionalmente, los molariformes superiores son morfológicamente muy similares a aquellos de Glyptodon, P. uquiensis y Glyptotherium ; a excepción del M1, todos ellos son claramente trilobulados.
En este contexto, se distingue claramente de Panochthus Burmeister, 1866 (Hoplophorinae Panochthini); en efecto, este último taxón presenta un perfil dorsal del cráneo convexo, posee barras post–orbitarias y carece de surco postero–labial en los molariformes superiores; a ésto debe sumarse una notable neumatización de la región rostral, en tanto que los nasales se extienden hacia delante y hacia abajo, adquiriendo una clara forma de "embudo".
Se diferencia, por otra parte, de Doedicurus Burmeister, 1874 (Doedicurinae) por exhibir un mayor grado de lobulación en los molariformes proximales (especialmente en el M3), por carecer de barras post–orbitarias, por la morfología de la apertura nasal (muy similar a aquella de P. uquiensis y Glyptodon) y además porque Doedicurus posee una estructura del basicráneo sumamente compleja, que probablemente sea propia del género o de la subfamilia.
Por último, cf. Paraglyptodon. chapalmalensis (MMP 4676) muestra una anatomía general del basioccipital ciertamente similar a la de los Glyptodontidae Glyptodontinae. Esta se caracteriza por ser relativamente simple y por la presencia de forámenes hipoglosos (estructuras no observadas en Hoplophorinae y Doedicurinae, que se hallan por el contrario, presentes en la mayoría de los gliptodontinos).
En vista frontal (Figura 2b) se observa que las aperturas nasales, aunque parcialmente preservadas, tienen un contorno subtrapezoidal, como en Glyptotherium , Glyptodon, mientras que P. uquiensis las narinas tienen un menor desarrollo dorso–ventral. Los forámenes infraorbitarios presentan el mismo desarrollo y ubicación que en Glyptodon y P. uquiensis. Por otra parte, el proceso descendente del maxilar muestra un menor diámetro transverso y se ubica más medialmente que en las dos formas mencionadas, como en Glyptotherium y a diferencia de P. uquiensis, en el cual estas estructuras están mucho mas expandidas en sentido transverso.
En vista lateral (Figura 2c), se aprecia que el perfil dorsal del cráneo tiene una inclinación equivalente a la de Glyptodon, aunque la región occipital se encuentra un poco más desarrollada. Por otro lado, la porción escamosa del arco cigomático es antero–posteriormente más extensa que en el género pleistoceno, debido principalmente a la mayor elongación postorbitaria de este ejemplar. Como en Glyptodon, P. uquiensis y Glyptotherium (ver Ameghino, 1889; Gillette y Ray, 1981) las órbitas se comunican libremente con la fosa temporal. Las escotaduras orbitarias tienen un contorno subcircular y se aprecia un evidente reborde óseo en su borde inferior, tal como en P. uquiensis y Glyptodon (ver Soibelzon et al., 2006); éste se encuentra mucho menos desarrollado en G. texanum Osborn, 1903 [AMNH (F:AM) 59583 (Gillette y Ray, 1981: fig. 5b)], G. arizonae Gidley, 1926 [UMMP 34826 (Gillette y Ray, 1981: fig. 10a)], G. floridanum Simpson, 1929 [USNM 6071 (Gillette y Ray, 1981: fig. 11a)] y G. cf cylindricum (Brown, 1912) [(MCN (–)] (Carlini et al., 2008b: 128, fig. 2D).
En vista dorsal (Figura 2d), se observa que la mayor parte de la región rostral está seriamente dañada, por lo que no es posible inferir la exacta morfología de esta área, aunque se encuentra claramente menos expandida en sentido transverso que en P. uquiensis. Sin embargo, sí se preserva el tubérculo lacrimal derecho, que posee una morfología muy similar a la de Glyptodon. Otros caracteres que el MMP 4676 comparte con el género pleistoceno son la presencia de una evidente cresta sagital y la marcada oblicuidad de las suturas fronto–parietales y crestas nucales. A diferencia de Glyptodon, el borde dorsal del arco cigomático es subparalelo al plano sagital del cráneo y no describe un semicírculo; esto es debido al mayor alargamiento del cráneo en el ejemplar aquí descripto.
En vista oclusal (Figura 2e), el proceso paraoccipital exhibe un grado de desarrollo comparable al de Glyptodon y Glyptotherium y el foramen infraorbitario está situado a nivel del plano de proyección extendido entre M2 y M3 como en estos géneros. Sin embargo, la fosa glenoidea es más extendida antero–posteriormente y proporcionalmente más reducida en sentido transverso que en éstos.
Los molariformes superiores tienen una morfología muy similar a la de P. uquiensis, Glyptodon, y Glyptotherium , diferenciándose esencialmente de éstos últimos por presentar un M1 más simple, no lobulado. Lamentablemente, el mal estado de preservación de la superficie oclusal de los molariformes no permite confirmar la existencia de las ramificaciones secundarias de vasculodentina, un carácter usualmente considerado como una sinapomorfía de la subfamilia (Hoffstetter, 1958; Gillette y Ray, 1981; Scillato–Yané et al., 1995; Carlini y Scillato–Yané, 1999).
El M1 es de sección subtriangular, comparativamente más primitivo que en Glyptotherium , donde presenta una trilobulación incipiente, y Glyptodon, donde se halla perfectamente trilobulado, siendo más notorio este carácter en las especies del Bonaerense y Lujanense (G. clavipes Owen, 1839, G. reticulatus Owen, 1845).
El M2, si bien ya se encuentra claramente trilobulado, se diferencia esencialmente de los demás molariformes por la escasa prolongación lateral de los prismas, siendo en este aspecto muy similar al de Glyptotherium y en menor medida al de Boreostemma .
Tal como se observa en otros Glyptodontinae (P. uquiensis, Boreostemma spp., Glyptotherium spp. y Glyptodon spp.) los molariformes se hallan marcadamente trilobulados a partir del M3, experimentando una evidente expansión labio–lingual, y adquiriendo los lóbulos distales un surco de ubicación póstero–labial.
Comentarios taxonómicos. La evidencia morfológica sugiere la asignación de este nuevo y significativo material a la subfamilia Glyptodontinae. En líneas generales, existe una marcada semejanza entre éste, P. uquiensis y Glyptodon. Entre las principales evidencias halladas pueden mencionarse: a) es posible inferir la presencia de una apertura nasal de contorno subtrapezoidal; b) las órbitas están abiertas posteriormente, comunicándose libremente con la fosa temporal; c) aunque parcialmente fragmentada, el área rostral presenta, en vista dorsal, el típico contorno subtriangular de Glyptodon y P. uquiensis, y posiblemente habría tenido un acortamiento semejante; d) de igual modo, el tubérculo lacrimal y el reborde óseo presente en el margen antero–inferior de la escotadura orbitaria, muestra una morfología muy similar a la de los dos taxones mencionados (ver Ameghino, 1889; Soibelzon et al., 2006); e) los molariformes superiores están, a excepción del M1, claramente trilobulados, y morfológicamente son muy similares a aquellos de Glyptodon, P. uquiensis y Glyptotherium .
Su clasificación tentativa como cf. Paraglyptodon chapalmalensis se debe a que tanto el holotipo de la especie (MACN 6162) como el ejemplar aquí descripto (MMP 4676) tienen una procedencia geográfica y estratigráfica muy similares; en efecto, ambos provienen de la sección superior de la Fm. Chapalmalal y fueron exhumados de los alrededores de la localidad de Mar del Plata, provincia de Buenos Aires, Argentina. Adicionalmente, no se ha hallado evidencia alguna de la existencia de otra especie de Cingulata Glyptodontinae para dicha unidad; de hecho, los osteodermos que han sido exhumados de estos niveles por parte del personal del Museo de Ciencias Naturales de Mar del Plata (Dondas, obs. pers.) son muy similares en su morfología a aquellos asignados a P. chapalmalensis. Sin embargo, el desconocimiento de la anatomía craneana que afecta a la especie terciaria impide, por el momento, realizar una asignación segura y definitiva del ejemplar.
CONCLUSIONES
Como ha sido señalado previamente, los integrantes de la subfamilia Glyptodontinae reúnen claras sinapomorfías cráneo–dentarias y exoesqueletarias que sustentan su monofilia (ver Ameghino, 1889; Hoffstetter, 1958; Pascual et al., 1966; Gillette y Ray, 1981; Soibelzon et al., 2006; Scillato–Yané et al., 1995; Carlini et al., 2008a, 2008b). Entre éstas se destacan: a) coraza dorsal gruesa y sólida, con un patrón de ornamentación en "roseta" constante y sin figuritas periféricas accesorias; b) osteodermos con superficie externa fuertemente punteada a rugosa y con la sección de los surcos en "U"; c) osteodermos marginales de gran tamaño, en forma de fuertes tubérculos cónicos; d) coraza caudal consistente en una serie de anillos móviles (con última hilera compuesta por placas usualmente cónicas) seguidos de un tubo caudal corto; e) cráneo antero–posteriormente comprimido y alto, truncado proximalmente por el acortamiento de los nasales y premaxilares; f) barras post–orbitarias incompletas; g) apertura nasal relativamente alta, de contorno subtrapezoidal; h) molariformes con alto grado de lobulación, por lo general provistos de ramificaciones secundarias de vasculodentina.
En este marco, el estudio sistemático efectuado sobre las especies de antigüedad Chapadmalalense (Plioceno) asignadas a esta subfamilia indica que P. chapalmalensis es efectivamente un Glyptodontinae, al tiempo que muestra significativos caracteres diagnósticos. La superficie expuesta de sus osteodermos presenta un grado de evolución intermedio entre aquellos de G. tuberifer ("Araucanense"; Mioceno tardío–Plioceno) y P. uquiensis ("Uquiense"; Plioceno tardío–Pleistoceno temprano), especialmente a nivel de la morfología de los surcos y de la rugosidad externa observada.
A su vez, el análisis comparado realizado sobre P. dubium, corrobora su sinonimia con P. chapalmalensis, en tanto no es posible observar ninguna diferencia morfológica significativa con éste; adicionalmente, ambos taxones comparten una misma procedencia geográfica y estratigráfica, hecho que refuerza aún más la hipótesis planteada.
Por último, es claro que los rasgos diagnósticos reconocidos por Castellanos (1953) para "Paraglyptodon" paranensis, son producto del desgaste que afecta la superficie externa del osteodermo y puntualmente, su mala preservación impide observar características morfológicas distintivas que permitan su asignación a la subfamilia Glyptodontinae. Es importante destacar que la presencia de surcos amplios y figuritas periféricas pequeñas, subcirculares y mal delimitadas son rasgos también presentes en los Hoplophorinae Palaehoplophorini Hoffstetter (ver Hoffstetter, 1958; Pascual et al., 1966) un clado basal de gliptodóntidos bien representado en la Formación Ituzaingó (ver Cione et al., 2000). En definitiva, esta forma debe ser considerada como nomen vanum (sensu Mones, 1986: 233; Simpson, 1948: 31), condición sustentada además por sus imprecisas procedencias geográfica y estratigráfica.
Teniendo en cuenta los resultados obtenidos, se concluye que el único Glyptodontinae válido reconocido hasta el momento para el Plioceno del extremo austral de América del Sur y fósil guía del Piso/Edad Chapadmalense superior (ca. 3.5–3.2 Ma; ver Cione y Tonni, 1995, 1999, 2005) es Paraglyptodon chapalmalensis.
Por lo tanto, creemos prudente asignar de manera tentativa el ejemplar MMP 4676 a cf. Paraglyptodon chapalmalensis dado que indudablemente representa un Glyptodontinae y proviene de los mismos niveles sedimentarios y de la misma región geográfica que MACN 6162 y MACN 6285. Se evita así agregar un nuevo taxón a la ya compleja taxonomía de los Glyptodontidae originando, de esta manera, un sobredimensionamiento sistemático innecesario. Si bien no es imposible, resultaría improbable el hallazgo de otro Glyptodontinae en la misma localidad fosilífera y en los mismos niveles estratigráficos que P. chapalmalensis, aunque no se descarta definitivamente esta posibilidad.
Desde un punto de vista morfológico, cf. P. chapalmalensis muestra claras afinidades con los más derivados Glyptodontinae: P. uquiensis y Glyptodon spp. (e.g., morfología y brevedad relativas de la región rostral, estructura general del proceso descendente del arco cigomático y del tubérculo lacrimal, presencia de reborde óseo en el margen antero–ventral de la escotadura orbitaria). En este contexto, es particularmente destacable la alta lobulación que exhiben los molariformes superiores, claramente trilobulados (a excepción del M1) y con los prismas labiolingualmente expandidos a partir del M3. No obstante, no alcanza el grado presente en P. uquiensis y las especies del género Glyptodon, en los cuáles ya existe una trilobulación marcada y una fuerte expansión transversa de la serie dentaria a partir del M2; este último carácter sugiere una posible diferenciación a nivel genérico entre P. chapalmalensis (MMP 4676) y P. uquiensis (MACN 5327), que no sería comprobable a nivel exoesqueletario.
Confiamos en que futuro hallazgos de materiales craneanos y de la coraza dorsal asociados permitan solucionar definitivamente los interrogantes aquí planteados. Independientemente de esto, cf. Paraglyptodon. chapalmalensis (MMP 4676) constituye, hasta el momento, el registro craneano más antiguo y completo conocido de un Glyptodontidae Glyptodontinae para el Terciario del extremo sur de América del Sur, siendo por otra parte, el primero documentado para el Neógeno de la Región Pampeana de Argentina.
AGRADECIMIENTOS
Los autores desean expresar su reconocimiento al Dr. Alejandro Kramarz (MACN) y al Lic. Lucas Pomi (MLP) por facilitar la revisión de los materiales alojados en la colecciones de dichas instituciones. Este agradecimiento se hace extensivo a los Dres. Daniel Perea y Martín Ubilla (Facultad de Ciencias, Universidad de la República, Uruguay) por las sugerencias efectuadas, que mejoraron sustancialmente la calidad de esta contribución. Este trabajo fue parcialmente financiado por el proyecto PICTO–UNNE 00164.
REFERENCIAS
Ameghino, F., 1881, La antigüedad del hombre en el Plata: Paris, G. Masson, Buenos Aires, Igon Hermanos, v. 2, 557 pp. [ Links ]
Ameghino, F., 1889, Contribución al conocimiento de los mamíferos fósiles de la República Argentina: Actas de la Academia Nacional de Ciencias de Córdoba, 6, 1–1027. [ Links ]
Ameghino, F., 1891, Nuevos restos de mamíferos fósiles descubiertos por Carlos Ameghino en el Eoceno inferior de la Patagonia austral. Especies nuevas, adiciones y correcciones: Revista Argentina de Historia Natural, 1, 289–328. [ Links ]
Ameghino, F., 1908, Las formaciones sedimentarias de la región Litoral de Mar del Plata y Chapalmalal: Anales del Museo Nacional de Buenos Aires, 3(10), 343–428. [ Links ]
Brown, B., 1912, Brachyostracon, a new genus of glyptodonts from México: American Museum of Natural History Bulletin, 31(17), 167–177. [ Links ]
Burmeister, G., 1866, Fauna Argentina: Anales del Museo Público de Buenos Aires, serie 1, tomo 1. [ Links ]
Burmeister, G., 1874, Monografía de los Glyptodontes en el Museo Público de Buenos Aires: Anales del Museo Público de Buenos Aires, serie 1, tomo 2. [ Links ]
Cabrera, A., 1944, Los Gliptodontoideos del Araucaniano de Catamarca: Revista del Museo de La Plata (Nueva Serie), 3(15), 5–76. [ Links ]
Carlini, A.A., Scillato–Yané, G.J., 1999, Evolution of Quaternary Xenarthrans (Mammalia) of Argentina: Quaternary of South America and Antarctic Peninsula,12, 149–175. [ Links ]
Carlini, A.A., Vizcaíno, S.F., Scillato–Yané, G.J., 1997, Armored Xenarthrans: A unique taxonomic and ecologic assemblage, en Kay, R.F., Madden, R.H., Cifelli, R.L., Flynn, J.J. (eds.), Vertebrate Paleontology in the Neotropics. The Miocene fauna of La Venta, Colombia: Washington, Smithsonian Institution Press, 213–226. [ Links ]
Carlini, A.A., Zurita, A.E., Aguilera, O., 2008a, North American Glyptodontines (Xenarthra, Mammalia) in the upper Pleistocene of northern South America: Palaeontologische Zeitschrift, 82(2), 125–138. [ Links ]
Carlini, A.A., Zurita, A.E., Scillato–Yané, G. J., Sánchez, R., Aguilera, O., 2008b, A new Glyptodont species from Codore Formation (Pliocene), Estado Falcón (Venezuela), and the 'Asterostemma' problem: Palaeontologische Zeitschrift, 82(2), 139–152. [ Links ]
Castellanos, A., 1932, Nuevos géneros de gliptodontes en relación con su filogenia: Physis, 11(38), 92–100. [ Links ]
Castellanos, A., 1953, Descripción de restos de Paraglyptodon uquiensis n. sp. de Uquía (Senador Pérez), Jujuy: Memorias del Museo de Entre Ríos, 32 (Paleontología), 1–32. [ Links ]
Cione, A.L., Tonni, E.P., 1995, Bioestratigrafía y cronología del Cenozoico de la región Pampeana, en Alberdi, M.T., Leone, G., Tonni, E.P., (eds.), Evolución biológica y climática de la región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental: Madrid, Consejo Superior de Investigaciones Científicas, Museo Nacional de Ciencias Naturales, Monografías, 12, 47–74. [ Links ]
Cione, A.L., Tonni, E.P., 1999, Biostratigraphy and chronological scale of uppermost Cenozoic in the Pampean area, Argentina: Quaternary of South America and Antarctic Peninsula, 12, 23–51. [ Links ]
Cione, A.L., Tonni, E.P., 2005, Bioestratigrafía basada en mamíferos del Cenozoico Superior de la provincia de Buenos Aires, Argentina, en 16° Congreso Geológico Argentino, La Plata, Argentina, Relatorio, 183–200. [ Links ]
Cione, A.L., Azpelicueta, M.M., Bond, M. Carlini, A.A., Casciotta, J.R., Cozzuol, M. A., de La Fuente, M., Gasparini, Z., Goin, F.J., Noriega, J.I., Scillato–Yané, G. J., Soibelzon, L., Tonni, E.P., Verzi, D., Vucetich, M.G., 2000, Miocene vertebrates from Entre Ríos province, eastern Argentina: Revista del Instituto Superior de Correlación Geológica, Serie Correlación Geológica, 14, 191–237. [ Links ]
Cope, E., 1889, The Edentata of North America: American Naturalist, 23, 657–664. [ Links ]
Gidley, J.W., 1926, Fossil Proboscidea and Edentata of the San Pedro Valley, Arizona: United States Geological Survey, Professional Paper 140B, 83–95, 13 plates, 4 figures. [ Links ]
Gillette, D.D., Ray, C.E., 1981, Glyptodonts of North America: Smithsonian Contributions to Paleobiology, 40, 1–251. [ Links ]
Gray, J., 1869, Catalogue of carnivorous, pachydermatous, and edentate Mammalia in the British Museum: London, Taylor and Francis, 398 pp. [ Links ]
Hoffstetter, R., 1958, Xenarthra, en Piveteau, J. (ed.): Traité de Paléontologie, 6. L' origine des mammifère et les aspects fondamentau de leur évolution, v. 2, Mammifères, évolution: París, Masson, 535–636. [ Links ]
Huxley, T.H., 1864, On the osteology of the genus Glyptodon: Proceedings of the Royal Society of London, 13, 108 pp. [ Links ]
Illiger, C., 1811, Prodromus systematis mammalium et avium; additis terminis zoographicis utriusque classis, eorumque versione germanica: Berlin, C. Salfeld, XVIII + 301 pp. [ Links ]
McKenna, M.C., Bell, S.K., 1997, Classification of Mammals: Above the Species Level: New York, Columbia University Press, 631 pp. [ Links ]
Mones, A., 1986, Nomen Dubium vs. Nomen Vanum: Journal of Vertebrate Palaeontology, 9(2), 232–234. [ Links ]
Ortiz–Jaureguizar, E., Cladera, G.A., 2006, Paleoenvironmental evolution of Southern South America during the Cenozoic: Journal of Arid Environments, 66, 498–532. [ Links ]
Osborn, H.F., 1903, Glyptotherium texanum, a new glyptodont, from the lower Pleistocene of Texas: Bulletin of the American Museum of Natural History, 19 (17), 491–494. [ Links ]
Owen, R., 1838, Fossil Mammalia, en Darwin, C. (ed.), The Zoology of the voyage of H.M.S. Beagle, under the command of Capitain Fitzroy, during the years 1832–1836: Londres, Smith Elder and Co., 40 pp., 7 láminas. [ Links ]
Owen, R., 1839, Note on the Glyptodon, en Parish, W. (ed.), Buenos Aires and the Provinces of the Rio de La Plata; Londres, John Murray, 1–178. [ Links ]
Owen, R., 1845, Descriptive and illustrated catalogue of the fossil organic remains of Mammalia and Aves: Museum of the Royal College of Surgeons of London, 391 pp. [ Links ]
Pascual, R., Ortega–Hinojosa, E.J., Gondar, D.G, Tonni E.P., 1966, Vertebrata, en Borrello, A.V (ed.), Paleontografía Bonaerense. Vertebrata I: Buenos Aires, Argentina, Comisión de Investigaciones Científicas de la provincia de Buenos Aires, 202 pp. [ Links ]
Reguero, M., Candela, A.M., Alonso, R.N., 2007, Biochronology and biostratigraphy of the Uquía Formation (Pliocene–early Pleistocene, NW Argentina) and its significance in the Great American Biotic Interchange: Journal of South American Earth Sciences, 23, 1–16. [ Links ]
Rovereto, C., 1914, Los estratos araucanos y sus fósiles: Anales del Museo Nacional de Historia Natural de Buenos Aires, 25, 250 pp. [ Links ]
Scillato–Yané, G.J., Carlini, A.A., Vizcaíno, S.F., Ortiz–Jaureguizar, E., 1995, Los Xenarthros, en Alberdi, M. T., Leone, G., Tonni, E. P. (eds.), Evolución biológica y climática de la región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental: Madrid, Consejo Superior de Investigaciones Científicas, Museo Nacional de Ciencias Naturales, Monografías, 12, 183–209. [ Links ]
Simpson, G.G., 1929, Pleistocene mammals fauna of the Seminole Field, Pinellas County, Florida: Bulletin of the American Museum of Natural History, 56, 561–599. [ Links ]
Simpson, G.G., 1947, A Miocene gliptodonte from Venezuela: American Museum Novitates, 1368, 1–10. [ Links ]
Simpson, G.G., 1948, The beginning of the age of mammals in South America, 1: Bulletin of the American Museum of Natural History, 91, 1–232. [ Links ]
Soibelzon, E., Zurita, A.E., Carlini, A.A., 2006, Glyptodon munizi Ameghino (Mammalia, Cingulata, Glyptodontidae): redescripción y anatomía: Ameghiniana, 43(2), 377–384. [ Links ]
Soibelzon, E., Tonni, E.P., Bidegain, J.C., 2008, Cronología, magnetoestratigrafía y caracterización bioestratigráfica del Ensenadense (Pleistoceno inferior–medio) en la Ciudad de Buenos Aires: Revista de la Asociación Geológica Argentina, 63(3), 421–429. [ Links ]
Villaroel, C., 1983, Descripción de Asterostemma? Acostae, nueva especie de propalaehoplophorino (Glyptodontidae, Mammalia) del Mioceno de La Venta, Colombia: Geología Norandina, 7, 29–34. [ Links ]
Webb, S.D., 2006, The Great American Biotic Interchange: Patterns and Processes: Annals of the Missouri Botanical Garden, 93(2), 245–257. [ Links ]
Zurita, A.E., Carlini, A.A., Gillette, D., 2008a, Glyptotherium–Glyptodon (Xenarthra, Glyptodontidae, Glyptodontinae): anatomy and palaeobiogeography: Journal of Vertebrate Paleontology (Supplement), 28(3), p. 15. [ Links ]
Zurita, A.E., Carlini, A.A., Scillato–Yané, G.J., Arenas Mosquera, J.E., Jesús Goméz, A., 2008b, Los Glyptodontinae (Xenarthra, Glyptodontidae) del Mioceno de La Venta, Colombia, en 3° Congreso Latinoamericano de Paleontología de Vertebrados, Nequén, Argentina, p. 34. [ Links ]
Zurita, A.E., Miño–Boilini, A.R., Soibelzon, E., Carlini, A.A., Paredes–Ríos, F., 2009, The diversity of Glyptodontidae (Xenarthra, Cingulata) in the Tarija Valley (Bolivia): systematic, biostratigraphic and paleobiogeographic aspects of a particular assemblage: Neues Jahrbuch für Geologie und Palaontologie 251(2), 225–237. [ Links ]

