










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.25 no.2 Ciudad de México ago. 2008
Morphometric analysis of equid cheek teeth using a digital image processor: a case study of the Pleistocene Cedazo local fauna equids, Mexico
Análisis morfométrico de molares de équidos usando un procesador digital de imágenes: un estudio de caso de los équidos de la fauna Cedazo del Pleistoceno, México
Christian Raúl Barrón–Ortiz1,2,*, Gilfredo de la Riva–Hernández1, and Raúl Barrón–Corvera3
1 Departamento de Biología, Centro de Ciencias Básicas, Universidad Autónoma de Aguascalientes, Av. Universidad # 940, 20100, Aguascalientes, Ags., México.
2 Present address: Instituto Nacional de Antropología e Historia, Centro INAH Aguascalientes, Juan de Montoro #226, Zona Centro, 20000 Aguascalientes, Ags., México. * chbarron45@yahoo.com
3 Facultad de Ingeniería, Universidad Autónoma de Zacatecas, Av. Ramón López Velarde # 810, 98000, Zacatecas, Zac. México.
Manuscript received: October 8, 2007
Corrected manuscript received: February 25, 2008
Manuscript accepted: March 13, 2008
ABSTRACT
The occlusal pattern of the cheek teeth has traditionally been one of the most widely used features to determinate equid species. Nevertheless, its large ontogenetic variation and subjective assessment of characters has diminished its reliability. For this reason, many workers have reduced emphasis on the cheek teeth in identifying equid species and have turned their attention to other skeletal elements. We re–emphasize the analysis of the oclusal pattern to determine the number of equid species present in the Cedazo local fauna, Aguascalientes, Mexico. In contrast to previous studies, we only analyzed cheek teeth with an equivalent stage of wear and developed a computer program that integrates a digital image processor to analyze the occlusal pattern and also to take measurements of different features of the occlusal surface. Selecting teeth with an equivalent stage of wear reduced the usable sample size significantly. For this reason, the analysis concentrated on the third upper premolars (P3), because they represent the largest sample size. Specimens of Equus caballus, E. asinus and hybrids were also included to serve as controls. Principal Components and Canonical Discriminant Functions analyses were performed. The extant species were positively discriminated. Moreover, these analyses suggest the presence of two distinct P3 morphotypes for the fossil sample studied, which indicate the possible occurrence of at least two separate equid species for the Cedazo local fauna. However, it is prudent to collect more material and evaluate these results with a larger sample size and using the remaining tooth positions. The computer program developed in this study serves as a useful tool for analyzing the occlusal pattern of equid cheek teeth.
Key words: morphometrics, vertebrate paleontology, equids, Cedazo local fauna, Pleistocene, Mexico.
RESUMEN
El patrón o dibujo oclusal de los molares ha sido tradicionalmente uno de los rasgos más utilizados para determinar especies de équidos. Sin embargo, una gran variación ontogenética y la subjetividad en la valoración de los caracteres dentales ha disminuido su confiabilidad. Es por ello que muchos investigadores han reducido el énfasis en los molares para la identificación de especies de équidos y han concentrado su atención en otros elementos esqueléticos. En el presente trabajo se retoma el análisis del patrón oclusal de los molares para determinar el número de especies de équidos presentes en la fauna Cedazo, Aguascalientes, México. En contraste con otros estudios, solamente se analizaron aquellos molares con un grado de desgaste equivalente. Además, se desarrolló un programa de cómputo que integra un procesador digital de imágenes el cual permitió analizar el patrón oclusal, así como realizar mediciones de diversos rasgos del mismo. El seleccionar molares con un grado de desgaste equivalente redujo el tamaño de muestra efectivo drásticamente. Por tal motivo, solamente se estudiaron los terceros premolares superiores (P3), los cuales presentaban el tamaño de muestra más grande. Así mismo, se incluyeron ejemplares de Equus caballus, E asinus e híbridos de éstos, con la finalidad de actuar como especímenes de control. Se realizó un análisis de Componentes Principales, además de un análisis Canónico de Funciones Discriminantes. Como era esperado, estos análisis segregaron a las especies actuales. Más aún, sugieren la existencia de dos morfotipos de P3 para la muestra fósil estudiada, lo que pudiera indicar la presencia de al menos dos especies distintas de équidos para la fauna Cedazo. Aunque estos resultados son prometedores, es conveniente colectar más material y evaluar los resultados con un tamaño de muestra más grande, que además incluya el resto de las posiciones dentales. El programa de cómputo desarrollado en esta investigación prueba ser una herramienta útil para el análisis del patrón oclusal de molares de équidos.
Palabras clave: morfometría, paleontología de vertebrados, équidos, fauna Cedazo, Pleistoceno, México.
INTRODUCTION
Southeast from the city of Aguascalientes, México, several streams (including El Cedazo, San Francisco, Cobos, Pargas; Figure 1) cut through Pleistocene sediments that have yielded a varied fossil fauna (Mooser, 1958; Mooser and Dalquest, 1975; Guzmán–Gutiérrez et al, 1997). This fossil assemblage was first studied during the 1950's by O. Mooser, who christened it the Cedazo local fauna (Mooser, 1958). Montellano–Ballesteros (1990) assigned it an Irvingtonian to Rancholabrean age, based on the taxa that have been found.
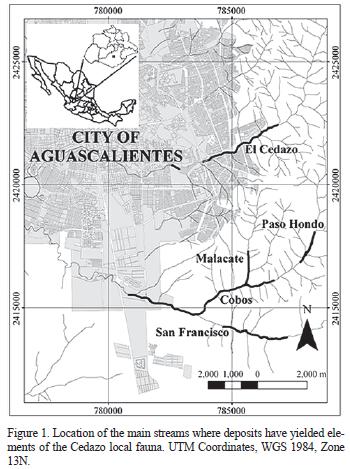
The most abundant material and the one that has been studied in most detail, corresponds to the equids. Nevertheless, the number of equid species present in this assemblage remains unresolved. Some authors have proposed from as many as ten (Mooser, 1958) to seven species (Mooser and Dalquest, 1975), while others have suggested that only two (Guzmán–Gutiérrez et al., 1997; Malgarejo–Damián, 2007) or one equid species (Reynoso–Rosales and Montellano–Ballesteros, 1994) are present. This scenario is not exclusive for the Cedazo local fauna and reflects the still problematic state of the Equus systematics (Winans, 1989; Eisenmann, 2004; Churcher, 2006). This is because many of the characters that have been used to determine species are either too variable for reliability (i.e., cheek teeth) or too similar (i.e., post–cranial elements) to really give much insight into specific variation (Winans, 1989; MacFadden, 1992; Churcher, written communication 2008). The occlusal pattern of the cheek teeth, which has been traditionally one of the most widely used features to determine equid species, changes drastically as the teeth wear down (Gidley, 1901; Carranza–Castañeda and Ferrusquia–Villafranca, 1979). This variation, in addition to the subjective assessment of dental characters, significantly lowers the reliability of the cheek teeth for determining equid species (MacFadden, 1992).
Some authors have suggested a quantitative approach for the analysis ofequid cheek teeth (e.g., Reynoso–Rosales and Montellano–Ballesteros, 1994). However, the dimensions commonly used are restricted mainly to four: antero–posterior length, transverse width, protocone length, and tooth height. The first three dimensions can be useful in the discrimination of equid species when these differ significantly in size. But when that is not the case, a great deal of overlap between species may exist, making discrimination between them difficult.
This study presents an analysis of the equid cheek teeth from the Cedazo local fauna, but with two significant differences: 1) Instead of analyzing all the cheek teeth, only those with an equivalent stage of wear were studied, in order to control ontogenetic variation; 2) a computer program that integrates a digital image processor was developed. This program performs an objective analysis of the cheek teeth and allows the user to take measurements of any feature of the occlusal pattern; eliminating the subjectivity in assessing tooth characters and increasing the number of variables that can be analyzed quantitatively for taxonomic purposes.
MATERIALS AND METHODS
A Kodak Easy Share DX6490 digital camera was used to photograph all the equid upper cheek teeth from the Cedazo local fauna deposited in the following collections: 1) Colección Nacional de Paleontología, Instituto de Geología, UNAM; 2) Laboratorio de Paleontología, desarrollo turístico "El Caracol," Secretaría de Turismo (SECTURE), Aguascalientes, Ags; 3) Colección de Paleontología de la Universidad Autónoma de Aguascalientes; 4) Biól. Alberto Rodríguez, personal collection, Aguascalientes, Ags; and 5) Luis Miguel Vázquez Sánchez, personal collection, Aguascalientes, Ags. Atotal of 80 teeth were photographed: 11 P2,s, 16 P3,s, 15 P4,s, 16 M1,s, 10 M2,s, and 12 M3,s. Each photograph was taken perpendicular to the occlusal surface of the tooth.
To be able to make valid comparisons and to reduce the ontogenetic variation due to tooth wear, only those cheek teeth with an equivalent stage of wear were studied. The stage of wear was determined as it is subsequently described. Crown height relates directly with tooth wear: as the tooth wears down its crown height decreases. Thus, it could be used to determine equivalent stages of wear between equids with a comparable hypsodonty index. We did not use a predetermined crown height, because the teeth showed a range of different sizes; for example, in large and small teeth, a crown height of 3 3 mm does not represent the same stage of wear for both sizes. Therefore, a ratio of tooth size to crown height was used as the criterion to determine an equivalent stage of wear. Transverse width was used as the indicator of tooth size rather than anteroposterior length, because the latter changes with crown and interdental wear, whereas transverse width remains less variable (Gidley, 1901). Using this criterion, we analyzed only those cheek teeth that had a tooth crown height similar to its corresponding transverse width (ratio 1:1); allowing a maximum margin of difference of 5 mm. Crown height was measured along the mid–line of the posterior side of the tooth, from the top of the crown down to the point just before the metastyle converges with the mesostyle.
Selecting teeth with an equivalent stage of wear reduced the usable sample size to just 22 cheek teeth: four P2,s, seven P3,s, three P4,s, four M1,s, three M2,s, and one M3. Moreover, these should not be analyzed as a combined sample; each type of tooth has to be analyzed separately. For this reason, only the P3 were studied, because of their larger sample size. The third upper premolars (P3) analyzed are listed in Table 1. To increase the sample size, the right P3 FC 205 (Mooser, 1958, fig. 10) in the original description of the Cedazo local fauna was included.
In addition, a sample of P3,s of extant equids were also included in the analysis to serve as controls and test the discrimination potential of the morphometric dimensions used in this study. These samples were: 21 P3,s from domestic horses (Equus caballus), three from donkeys (E. asinus) and two from hybrids. These teeth were donated by the meat processing company Frigorífico Zacatecas, except for two of E. caballus that were taken from skulls in the Zoological Collection, Universidad Autónoma de Aguascalientes.
To be able to compare these teeth with the eight P3,s from the Cedazo local fauna listed in Table 1, they were sectioned to show a similar wear stage according to the criterion given above. This was done at the Laboratorio de Petrografía y Paleontología, INEGI, Aguascalientes. Five of the 21 E. caballus premolars were not sectioned as they already were worn to the required tooth height.
Digital image analysis
A computer program that integrates a digital image processor was developed using the computer software MATLAB 7.0 (The MathWorks, 2004). This program, termed here 'Análisis de Patrones Oclusales' (APO) and which is available upon request, allows to study the occlusal patterns of cheek teeth on digital photographs. The program is capable of representing the enamel oclusal pattern as a matrix of single points in a Cartesian grid, based on a digital image of the occlusal surface of the tooth (Figure 2a), as is briefly described below.
The program identifies the occlusal pattern from the image according to color, highlighting a specific value and "shutting off' all of the rest. Due to preservation factors and light, the occlusal pattern did not show a distinct color, so it was necessary to redraw it using a uniform color (Figure 2b). In this way, only those cells from the raster image with this color are given a value of one, whereas the rest of the cells are given a value of zero, thus obtaining a binary image that shows only the enamel occlusal pattern as three distinct lines: one for the outline of the occlusal surface and one for each fossette. To be able to analyze the occlusal pattern as a matrix of single points, the lines of the occlusal pattern in the binary image, which have a thickness of several pixels, must to be converted into one pixel–thick lines. This was done using the 'skeletonization' function of MATLAB, which calculates the center of the lines from the previous binary image. These central lines have a resulting thickness of one pixel (Figure 2c). From this new binary image, the program obtains the (x,y) coordinates of the occlusal pattern (Figure 2d). These coordinates were scaled to the real size of the tooth by placing a small ruler next to the occlusal surface, so that in the image a relation between the number of pixels for a given length of the ruler could be established. Moreover, the total number of these coordinate points was kept constant for every tooth, so that comparisons between different teeth were possible. With the occlusal pattern drawn in (x,y) coordinates, it is easy to obtain the length between any two given points, allowing to measure any feature of the occlusal surface of the tooth.
It is important to mention that the enamel thickness is not constant throughout the occlusal pattern. To cope with this variation, special care was taken to redraw the occlusal pattern with the uniform color right through the middle of the enamel band. This was done so that the resulting point matrix would represent the real central line of the enamel band of the original tooth.
Measurements used in this study
With the aid of the APO software, measurements of 21 dimensions of the occlusal pattern were taken for each tooth studied (Appendix A). These dimensions and the terminology used are shown on Figure 3. Table 2 gives brief descriptions of each dimension.

Measurements of dimensions 2,3, 5 to 7,13 to 15,17 and 19 were taken to study characters that had previously been used by other authors (e.g., Quinn, 1956; Mooser, 1958) for equid taxonomy. Even though these authors used these characters qualitatively, the measurements taken reflected the variations described by them; for example, for 'postprotoconal groove narrow,' we measured the width of the postprotoconal groove (dimension 15, Figure 3). Measurements 1, 4, 8 to 12, 16, 18, 20 and 21 were characters that were considered to have a high potential for specific discrimination, on the basis of our previous investigations.
Because the object of this study was to carry out an analysis of the form of the occlusal pattern, the effects of differences in size was removed by normalizing the measurements. For every tooth, each of the 21 dimensions was divided by a value reflecting each tooth's size. This value is the diagonal of the quadrilateral formed by the anteroposterior length (Ap) and transverse width (Tr). The resulting ratios (Appendix B), where subjected to multivariate analyses.
Quantitative analyses
A Principal Components Analysis (PCA) was performed to identify patterns in the data and recognize different groups present in the sample under study. The relevant variables that contributed to the main principal components were identified. From these, a Canonical Discriminant Analysis (CDA) was performed to evaluate if the groups were correctly identified. These analyses were performed using the software STATISTICA7.0 (StatSoft, 2004), employing the default settings.
Once the groups were recognized, a mean occlusal enamel pattern for each distinct group was calculated using APO. To accomplish this, the teeth were scaled to the mean diagonal length of its corresponding group and then overlapped using a least squares criterion to minimize the distance between each (x,y) point of the occlusal pattern;, and obtaining in this way the best fit. Because APO assigns the same number of coordinate points for each tooth, it was possible to statistically analyze each point to obtain its mean. Drawing the mean of all the points creates a mean occlusal pattern for the teeth analyzed. It should be mentioned that a mean occlusal pattern is obtained under the assumption that the occlusal patterns within each group vary continuously. This is shown to be the case, on the basis of specimen analyses of Equus caballus.
RESULTS AND DISCUSION
Multivariate analyses
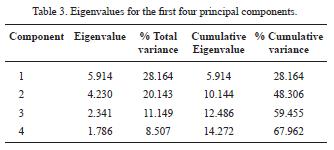
The first four principal components comprise nearly 70 % of the total variation (67.962 %; Table 3). Of these, the first two best explain most of the variation within the data and are the ones that better discriminate the specimens studied (Figure 4).
The first principal component separates six of the eight Cedazo third upper premolars and two oí Equus asinus from the rest of the teeth analyzed; whereas the second component separates all three E. asinus P3,s from the remaining specimens. Plotting both components, four different groups are identified (Figure 4): one comprising the three premolars of E. asinus, a second with six of the eight Cedazo premolars, a third containing the 21 P3,s of E. caballus and the remaining two Cedazo premolars, and a fourth comprised by the two hybrids. Distinction between these groups is clear, except for the hybrids, which marginally overlap with the group of E. caballus. One specimen clusters closely with the group of E. caballus whereas the other one falls between the groups of E. asinus and E. caballus.
The case of the hybrids is special because it is expected that they show features of E. asinus as well as E. caballus. As the sample size is incremented, it is expected that the hybrids cluster between these two species, with some specimens falling just within the E. asinus group and others within the E. caballus group. In the breeding of 'mules,' as the egg carries matrilineal DNA, this could influence the development of the embryo and, in a normal mule (jack x mare), it is the horse matrilineal DNA that may have a major influence (Churcher, written communication 2008). Unfortunately, it is not possible to test this since we do not know if the hybrid P3 that clusters close to the E. caballus group belongs to a mule (jack x mare), as the two hybrid P3,s here studied were obtained from broken and weathered skulls, which were not assignable to a particular hybrid type.
The most revealing conclusion that can be seen from the PCA results is that the specimens of Equus caballus and E. asinus are positively discriminated. Furthermore, six of the eight Cedazo premolars cluster in a distinct group whereas the remaining two fall within the group of E. caballus (Figure 4). These two premolars correspond to specimens c4 and c8 in Table 1. A Canonical Discriminant Analysis supports this arrangement of the data (Figure 5). With a Wilk's lambda of 0.008 (Table 4), this analysis is highly significant (Podani, 2000). Moreover, the CD A analysis shows that all of the specimens where correctly identified.
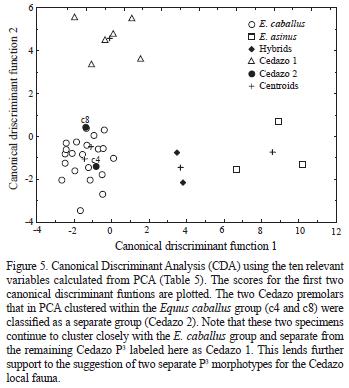
The results suggest the presence of two distinct equid P3 morphotypes for the Cedazo local fauna sample analyzed in this study, which may indicate the occurrence of at least two separate equid species. This contrasts with the revision of the Cedazo local fauna by Mooser and Dalquest (1975) where, on the basis qualitative appreciations of the occlusal pattern, the number of equid species was reduced from ten (originally in Mooser, 1958) to seven. These results also differ from those of Reynoso–Rosales and Montellano–Ballesteros (1994). These authors concluded, based on a univariate and qualitative statistical analysis, that only one equid species was present They identified this as E. conversidens. Guzmán–Gutierrez et al. (1997), and recently Malgarejo–Damián (2007), identified two equid species from the Cedazo local fauna (E. cf. excelsus/E. cf. conversidens and E. mexicanus/ E. conversidens, respectively) which agrees with the results here presented.
It is interesting to note, that one of the many criteria used by Guzmán–Gutierrez et al. (1997) and Malgarejo–Damián (2007) to identify two distinct equid species is size. They both identify a large and a medium size species. Our analysis supports the presence of at least two separate species based on the P3 morphology, regardless of size. Though, one of the morphotypes (Figure 6d) does appear to be slightly smaller.
Given the small sample size analyzed and the still problematic state in Equus systematics, at the present we do not feel confident to assign the P3,s studied to any particular species. A larger sample must be obtained and evaluated to justify a two species concept, and the results must reflect parallel situations for each tooth position. This will reveal if the two species pattern remains, or whether it becomes lost, showing an equid species with a high degree of morphological plasticity.
Regarding the issue of ontogenetic variation, it is well known to be an important factor to consider when working on the taxonomy of hypsodont equids (Gidley, 1901; Ferrusquia–Villafranca and Carranza–Castañeda, 1979; MacFadden, 1992). In the present PCA analysis, a P3 of Equus asinus was analyzed at two different stages of wear: one corresponding to that described in the methodology (tooth crown height similar to the transverse width; identified as 2b in Figure 4) and the other 33 mm above this level (2bb in Figure 4). It can be clearly seen that 2bb wrongly places within the E. caballus group. As was expected, this reinforces the suggestion that, to make a reliable discrimination it is important to keep the sample homogeneous, with cheek teeth in an equivalent stage of wear.
Currently, many authors have significantly reduced emphasis on the dentition, whether tooth rows or single teeth, as basis for discriminating equid species, because of its wide ontogenetic and intraspecific variation, and have turned their attention to other skeletal elements (e.g., Eisenmann and Karchoud, 1982; MacFadden, 1992). Contrary to this, the results here presented suggest that the dentition can be useful in discriminating species of Equus, as long as cheek teeth from a single position and an equivalent stage of wear are compared.
Analysis of the measurements used in this study
An important aspect of PCA is that it allows to identify those variables that contribute the most to the formation of each principal component (Manly, 2004). In this way, it is possible to identify the measurements that were more relevant for discriminating each group (Table 5). Thus, the six Cedazo P3,s are distinguished from the specimens of Equus caballus and the remaining two Cedazo premolars because the metastyle–mesostyle length (dimension 1, Figure 3) is proportionally shorter, as are the width of the postprotoconal grove (dimension 15) and the postfossette (dimension 17). Likewise, the parastyle width (dimension 10), mesostyle height (dimension 5), and the length of the posterior crest of the prefossette (dimension 20) are proportionally longer in the six Cedazo premolars. The P3,s of E. asinus are separated from the rest of the specimens studied because they show a proportionally shorter mesostyle base width (dimension 4) as well as a shallower metastyle–mesostyle valley (dimension 9) and a proportionally wider metacone (dimension 16).

Figure 6 shows the mean occlusal patterns calculated for the P3,s of the three Equus asinus teeth (Figure 6a), the specimens of E. caballus (Figure 6b), the six Cedazo P3,s (Figure 6c), and the remaining two Cedazo P3,s (Figure 6d). The differences in the features mentioned above are apparent, as is the possible occurrence of two distinct P3 morphotypes for the Cedazo sample analyzed, one (Figure 6d) morphologically more similar, in the dimensions studied, to the mean occlusal pattern of E. caballus (Figure 6b).
This similarity with the dimensions studied for the sample of Equus caballus is intriguing. Recently, Weinstock et al. (2005) performed a cladistic molecular study in which they analyzed the mitochondrial DNA sequence of North and South American late–Pleistocene fossil metapodials and specimens of extant species. The North American fossil sample arranged in two distinct clades, which they assign species status. One of this clade included the domestic horse, E. przewalskii, and all of the stout fossil metapodials analyzed by them. Further study will reveal if the Cedazo P3,s that cluster close to the specimens of E. caballus have any true affinity with them.
CONCLUSIONS
The computer program developed for this study proved to be a useful tool for analyzing equid cheek teeth. It allows to take measurements of any feature of the occlusal enamel of the tooth; thus eliminating the subjectivity in assessing tooth characters and increasing the number of variables that canbe incorporated in the quantitative analysis of equid cheek teeth for taxonomic purposes.
The multivariate quantitative analyses of the dimensions here considered positively discriminated the teeth of the control specimens. In addition, the results suggest the presence of two distinct equid P3 morphotypes for the Cedazo local fauna sample analyzed in this study. This indicates the occurrence of at least two separate equid species for this assemblage, which agrees with other recent studies. However, as the sample size is small, additional specimens must be collected and studied to substantiate this conclusion based on only P3,s. Furthermore, similar and parallel analyses should be performed on cheek teeth from the other five maxillary tooth positions and the six mandibular tooth positions.
Although an analysis comprising more species, including zebras and hemiones, is needed to fully evaluate the taxonomic significance of the occlusal pattern, these results suggest that the quantitative study of the occlusal surface of the cheek teeth can be useful in the discrimination oí Equus species, as long as cheek teeth from a single position and an equivalent stage of wear are compared. This could complement other studies that use different cranial and postcranial characters, helping to reveal a much clearer picture of the systematics of this important genus. Such comparative analyses would be useful in establishing the relative reliability of diverse elements in equine taxonomy.
ACKNOWLEDGMENTS
We would like to thank the coordinators of the different collections visited, for kindly making available the fossil material under their custody: Dra. Ma. del Carmen Perrilliat, Colección Nacional de Paleontología, Instituto de Geología, UNAM; Biól. Rubén Guzmán Gutiérrez, Laboratorio de Paleontología, desarrollo turístico "El Caracol," SECTURE, Aguascalientes, Ags; M. en C. Jaime Escoto Rocha, Colección de Paleontología de la Universidad Autónoma de Aguascalientes, UAA; Biól. Alberto Rodríguez, Personal Collection; Luís Miguel Vázquez Sánchez, Personal Collection. We also thank Juan de Dios Magallanes, owner of Frigorífico Zacatecas, and Juan Antonio Salazar, for letting us go through the waste piles and helping to collect modern equid teeth and skulls. We much appreciate the help of Mrs. Graciela Ortiz Lizalde in collecting and safekeeping the extant equid material analyzed in this study. Thanks are due to Gustavo Hernández, Francisco Palomino and Edgar Miguel Ángel Guzmán from the Laboratorio de Petrografía y Paleontología, INEGI, for sectioning some of the upper cheek teeth (P3) of the extant species. We thank Jaqueline and Taily Barrón Ortiz for reviewing the first draft of the manuscript. Very special thanks go to Biól. Elizabeth Romo Rábago for all her constant support. Finally, we would like to thank the reviewers, Bruce MacFadden and C.S. Churcher, for their valuable comments and suggestions which improved the original manuscript.
REFERENCES
Carranza–Castañeda, O., Ferrusquía–Villafranca, I., 1979, El género Neohipparion (Mammalia–Perisodactyla) de la fauna local Rancho el Ocote, (Plioceno medio) de Guanajuato, México: Universidad Nacional Autónoma de México, Revista del Instituto de Geología, 3(1), 29–38. [ Links ]
Churcher, C.S., 2006, Distribution and history of the Cape zebra (Equus capensis) in the Quaternary of Africa: Transactions of the Royal Society of South Africa, 61(2), 89–95. [ Links ]
Eisenmann, V., 2004, Equus: an evolution without lineages?, in 18th International Senckenberg Conference, VI International Palaeontological Colloquium, Late Neogene and Quaternary biodiversity and evolution: Regional developments and interregional correlations, Weimar, Germany: Terra Nostra, Conference volume, 2, 61–64. [ Links ]
Eisenmann, V., Karchoud, A., 1982, Analyses multidimensionnelles de métapodes d'Equus sensu lato (Mammalia, Perissodactyla): Bulletin du Museum national d'Histoire naturelle, Paris, 4e sér, Section C, 4(1–2), 75–103. [ Links ]
Evander, R.L., 2004, A revised dental nomenclature for fossil horses: American Museum of Natural History Bulletin, 285, 209–218. [ Links ]
Ferrusquía–Villafranca, I., Carranza–Castañeda, O., 1979, Revisión del género Neohipparion (Mammalia–Perisodactyla) en México: Universidad Nacional Autónoma de México, Revista del Instituto de Geología, 3(2), 126–137. [ Links ]
Gidley, J.W., 1901, Tooth characters and revision of the North American species of the genus Equus: American Museum of Natural History Bulletin, 14, 91–142. [ Links ]
Guzmán–Gutierrez, R., Acosta–Rincón, O.E., and Palomino–Sánchez, F., 1997, Catálogo de los sitios paleontológicos del estado de Aguascalientes: Aguscalientes, Ags., Gobierno del Estado de Aguascalientes, Cuadernos de Trabajo, 60, 38 p. [ Links ]
MacFadden, B.J., 1992, Fossil Horses, Systematics, Paleobiology and Evolution of the Family Equidae: New York, Cambridge University Press, 362 p. [ Links ]
Malgarejo–Damián, M. P., 2007, Diferenciación cuantitativa de especies de équidos del Pleistoceno de México: México, D.F., Universidad Nacional Autónoma de México, tesis de licenciatura, 132 p. [ Links ]
Manly, B.F., 2004, Multivariate statistical methods, a primer: New York, USA, Chapman & Hall/CRC, 3rd edition, 75–90. [ Links ]
Montellano–Ballesteros, M., 1990 (1992), Una edad del Irvingtoniano al Rancholabreano para la fauna Cedazo del Estado de Aguascalientes: Universidad Nacional Autónoma de México, Revista del Instituto de Geología, 9(2), 195–203. [ Links ]
Mooser, O., 1958, La fauna "Cedazo" del Pleistoceno de Aguascalientes: Universidad Nacional Autónoma de México, Instituto de Biología, Anales, 29, 409–452. [ Links ]
Mooser, O., Dalquest, W.W., 1975, Pleistocene mammals from Aguascalientes, Central Mexico: Journal of Mammalogy, 56(4), 781–820. [ Links ]
Podani, J., 2000, Introduction to the exploration of multivariate biological data: Netherlands, Backhuys Publishers, 263–270. [ Links ]
Quinn, J.H., 1956, Pleistocene Equidae of Texas: Fayetteville, USA, University of Arkansas, Ph.D. thesis, 83 p. [ Links ]
Reynoso–Rosales, V.H., Montellano–Ballesteros, M., 1994, Revisión de los équidos de la fauna Cedazo del Pleistoceno de Aguascalientes, México: Revista Mexicana de Ciencias Geológicas, 11(1), 87–105. [ Links ]
StatSoft, 2004, STATISTICA, ver. 7.0: Tulsa, OK, USA, StatSoft, 1 cd–rom, computer software. [ Links ]
The Math Works Inc., 2004, MATLAB The Language of Technical Computing, version 7.0: MathWorks Inc., 1 cd–rom, computer software. [ Links ]
Weinstock, J., Willerslev, E., Sher, A., Tong, W., Ho, S.Y.W., Rubenstein, D., Storer, J., Burns, J., Martin, L., Bravi, C., Prieto, A., Froese, D., Scott, E., Xulong, L., Cooper, A., 2005, Evolution, systematics, and phylogeography of Pleistocene horses in the New World: a molecular perspective: PLoS Biology, 3(8): e241. [ Links ]
Winans, M., 1989, A quantitative study of North American fossil species of the genus Equus, in Prothrero, D.R., Schoch, R.M. (eds.), The Evolution of Perissodactyls: New York, Oxford University Press, 262–297. [ Links ]

