










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBoletín de la Sociedad Botánica de México
versión impresa ISSN 0366-2128
Bol. Soc. Bot. Méx no.86 México jun. 2010
Taxonomía y florística
Delimitación de Neobuxbaumia mezcalaensis y N. multiareolata (Cactaceae) con base en análisis multivariados
Delimitation of Neobuxbaumia mezcalaensis and N. multiareolata (Cactaceae) based on multivariate analyses
Gabriel Arroyo–Cosultchi1, Teresa Terrazas2, 4, Salvador Arias3 y Lauro López–Mata1
1 Programa en Botánica, Colegio de Postgraduados.
2 Departamento de Botánica, Instituto de Biología, Universidad Nacional Autónoma de México. 4 Autor para la correspondencia: tterrazas@ibiologia.unam.mx
3 Jardín Botánico, Instituto de Biología, Universidad Nacional Autónoma de México.
Recibido: 8 de junio del 2009.
Aceptado: 6 de noviembre del 2009.
Resumen
Neobuxbaumia mezcalaensis y N . multiareolata son taxones pobremente definidos, donde la segunda especie fue descrita originalmente como variedad de la primera. Aunque varias propuestas de clasificación señalan que son especies independientes, existe aún controversia sobre su estatus taxonómico. Nuestra hipótesis de trabajo es que existen caracteres morfológicos que permiten la delimitación inequívoca de N . mezcalaensis y N. multiareolata. Para probarla, se realizaron análisis multivariados, con un total de 15 poblaciones, y como grupos para comparación también se analizaron representantes de especies relacionadas tales como N. squamulosa y de N. polylopha. Los resultados permiten reconocer que no existen fenotipos intermedios y que los individuos de las poblaciones analizadas se clasifican en sus respectivas especies. Se encontró que el tamaño y forma de la aréola, longitud de la espina central y patrón de distribución de las espinas radiales junto con otros cuatro caracteres referidos por autores previos, permiten diferenciar claramente a N. mezcalaensis de N. multiareolata. Con base en este cúmulo de evidencias proponemos que ambas especies pertenecen a taxones inequívocamente independientes.
Palabras claves: análisis discriminante canónico, cactáceas columnares, morfometría, espinas radiales.
Abstract
Neobuxbaumia multiareolata and N. mezcalaensis are taxa poorly defined, despite the second species was described originally as a variety of first. Although several classification proposals indicate that they represent different species, in the most recent cacti classification, both are considered a single species. The working hypothesis is that there are morphological characters that allow to separating N. mezcalaensis and N. multiareolata. Multivariate analyses were performed, with a total of 15 populations, to test if a boundary between N. mezcalaensis and N. multiareolata exists, and to compare the morphological characters with those of N. squamulosa and N. polylopha. The results allowed recognizing that intermediate phenotypes do not exist and that individuals of each analyzed populations were classified in their respective species. The areole size and form, central spine length and distribution pattern of the radial spines, plus four characters mentioned previously by other authors, allowed to differentiate N. mezcalaensis from N. multiareolata. Based on these evidences we propose that they represent independent species.
Keywords: Canonical discriminant analysis, columnar cacti, morphometrics, radial spines.
Los análisis multivariados han demostrado su utilidad para el reconocimiento de especies con amplia variación morfológica o pertenecientes a complejos de especies, en diversas familias de plantas (González–Andrés y Ortiz, 1996; Levin, 1998; Kephart et al., 1999; Henderson, 2002; Janovec y Harrison, 2002; De Luna y Gómez–Velasco, 2008). Los estudios donde los análisis multivariados han sido una herramienta para resolver complejos de especies en la familia Cactaceae son escasos pero contundentes (Baker y Pinkava, 1987; Chamberland, 1997; Baker y Johnson, 2000; Aguilar–Morales, 2004; Schmalzen et al., 2004; Baker, 2006; Muñoz–Urias et al., 2008). Más recientemente, se han utilizado también para reconocer la variación morfológica generada durante el proceso de domesticación de varios miembros de esta familia (Casas et al., 1999; Cruz y Casas, 2002; Arellano y Casas, 2003; Grimaldo–Juárez et al., 2007). Este tipo de análisis permite reconocer si la variación dentro de un taxón forma grupos discretos con lo cual se argumenta sobre el reconocimiento de especies o categorías infraespecíficas.
El género Neobuxbaumia Backeb. (Cactaceae) se distribuye en el bosque tropical caducifolio y el matorral xerófilo de la Depresión del Balsas, el Valle de Tehuacán–Cuicatlán, la vertiente del Pacífico y la vertiente del Golfo de México (Bravo–Hollis, 1978). Neobuxbaumia no se reconoce como un grupo monofilético en análisis filogenéticos con datos moleculares; sin embargo, forma parte de un clado con Cephalocereus y Pseudomitrocereus (grupo Cephalocereus) (Arias et al., 2003). El género como tradicionalmente se ha definido comprende plantas columnares monopódicas o con ramificación de escasa a profusa, tallos cilíndricos con pocas o numerosas costillas; flores tubular–campanuladas o tubular–infundibuliformes, nocturnas; frutos ovoides con espinas, restos de perianto persistentes, dehiscencia vertical en forma de estrella y pulpa blanca y semi–seca; semillas largas a muy largas, de color negro o café oscuro, lustrosas, lisas y con quilla (Bravo–Hollis, 1978; Barthlott y Hunt, 1993; Anderson, 2001; Arroyo–Cosultchi et al., 2007). Además de los trabajos taxonómicos realizados en varias especies de este género (Bravo–Hollis, 1932; Dawson, 1948, 1952; Bravo–Hollis et al., 1970, 1971 a y b, 1972; Bravo–Hollis, 1978; Scheinvar y Sánchez–Mejorada, 1990; Lau, 1994; Flores–Ortiz, 2004), también se han estudiado aspectos de su anatomía (Terrazas, 2000; Loza–Cornejo y Terrazas, 2003; Fuentes–Pérez, 2004; Evans, 2005), ecología (Valiente–Banuet et al., 1996, 1997; Esparza–Olguín et al., 2002, 2005; Godínez–Alvarez y Valiente–Banuet, 2004; Ramírez–Padilla y Valverde, 2005; Ruedas et al., 2006; Loza–Cornejo et al., 2008), genética (Rivera–Montoya, 2003) y quimotaxonomía (Flores–Ortiz et al., 2003).
El número de especies que conforman Neobuxbaumia varía de ocho a nueve y dos subespecies (Hunt, 1999; Anderson, 2001; Guzmán–Cruz et al., 2003; Hunt, 2006). Bravo–Hollis (1978) menciona que N. mezcalaensis (Bravo) Backeb., es la especie con mayor variación morfológica y de más amplia distribución del género, ya que se distribuye desde Jalisco hasta Oaxaca (Guzmán–Cruz et al., 2003). Bravo–Hollis (1978) observó en una población entre la ciudad de Colima y Armería individuos columnares con escamas papiráceas, largamente triangulares y acuminadas en la flor, pero no observó el fruto y mantuvo su identidad como N. mezcalaensis. Años más tarde, Scheinvar y Sánchez–Mejorada (1990) con base en un mayor número de colectas en los estados de Colima, Guerrero y Michoacán, la describieron como una nueva especie, Neobuxbaumia squamulosa Scheinvar & Sánchez–Mej., por presentar tallos ramificados y escamas papiráceas en las flores y frutos, rasgos que la diferencian de N. mezcalaensis.
Dawson (1952) reconoció tres variedades en Neobuxbaumia mezcalaensis, con base en la altura y diámetro del tallo, distancia interareolar y la altura de la costilla. Con estos caracteres propuso los epítetos N. mezcalaensis var. robusta (E.Y. Dawson) E.Y. Dawson para la población del Río Atoyac, Puebla; N. mezcalaensis var. mezcalaensis para las poblaciones del río Mezcala, Guerrero y N. mezcalaensis var. multiareolata (E.Y. Dawson) E.Y. Dawson para la población de Acahuizotla, Guerrero. Bravo–Hollis et al. (1972) propusieron que N. mezcalaensis var. multiareolata sea considerada a nivel de especie por presentar flores más cortas, pericarpelo globoso, escamas con margen eroso a ciliado, frutos más grandes y redondeados y semillas más grandes, con el nombre N. multiareolata (E.Y. Dawson) Bravo, Scheinvar & Sánchez–Mej. En la propuesta de clasificación clásica más reciente para cactáceas, Hunt (2006) no reconoce a este taxón como especie independiente, por lo que la trata como una subespecie de N. mezcalaensis. Aunque no presenta estudio o evidencias para sustentar su propuesta, es probable que sólo se base en el hábito columnar simple que comparten. Por lo tanto, el problema de su diferenciación taxonómica está pendiente de resolverse ya que no existe evaluación que permita definir claramente si N. mezcalaensis y N. multiareolata corresponden a taxones diferentes o bien a una sola especie. Con base en estos antecedentes, nuestra hipótesis de trabajo fue que existen caracteres morfológicos cuantitativos y cualitativos de estructuras vegetativas y reproductivas que permiten diferenciar inequívocamente a N. mezcalaensis de N. multiareolata. En el contexto particular de la filogenia de cactáceas columnares de Norte América (Pachycereeae), Neobuxbaumia es un género donde la delimitación de algunas de sus especies no está claramente definida (Arias et al., 2003). Los objetivos de esta investigación son establecer si N. mezcalaensis y N. multiareolata son especies independientes mediante el empleo de técnicas estadísticas de análisis multivariados e identificar caracteres que permitan reconocerlas en el campo; así como compararlas con N. squamulosa también segregada de N. mezcalaensis y con N.polylopha (DC.) Backeb, especie que nunca ha sido confundida con ninguna de las anteriores.
Materiales y métodos
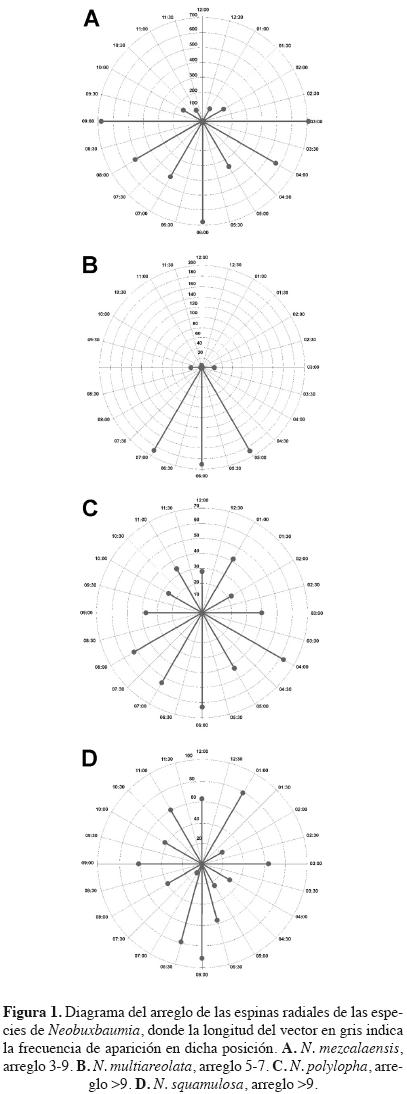
Obtención de datos. Con base en la revisión de ejemplares de herbario depositados en MEXU se planeó una recolecta en 15 poblaciones de Neobuxbaumia (Cuadro 1, Apéndice 1). En estas poblaciones se seleccionaron y marcaron 20 individuos reproductivos por población, excepto en La Huerta, Jalisco donde los individuos son menos de diez. A cada individuo se le midió la altura y el diámetro del tallo a la altura del pecho (DAP), el número de costillas a la altura del pecho; así como diez mediciones por individuo de: grosor y profundidad de las costillas, distancia entre costillas, distancia interareolar y la longitud y anchura de aréolas. Dado que las características de las espinas son importantes para distinguir entre taxones (Buxbaum, 1950), se cuantificó el número y longitud de las espinas centrales y radiales de cinco aréolas por individuo. Además se diseñó un método para registrar la posición angular de las espinas radiales siguiendo un sentido dextrógiro a partir de las 12 (12, 01, 02, 03, etc.). Su posición se determinó según la hora y un intervalo de 30' entre horas (12:00, 12:30, 01:00, 01:30, etc.). Con base en los valores de las frecuencias por posición angular se asignaron tres tipos para la disposición de las espinas radiales (Figura 1).
Se recolectaron de 3 a 5 flores y frutos por individuo y se preservaron en glicerina–alcohol etílico–agua (1:1:1). Semidieron diez y nueve caracteres de las flores y ocho de los frutos con un calibrador electrónico Serie 500 o por conteo directo. De 4 a 5 mediciones fueron obtenidas para los siguientes caracteres de la flor: longitud del podario del tubo, anchura del podario del tubo, longitud del podario del pericarpelo, del filamento interno, del filamento externo, del lóbulo del estigma, anchura del lóbulo y longitud de la escama del tubo y para los caracteres del fruto: longitud de la espina y longitud y anchura del podario. El resto de los caracteres fueron evaluados una sola vez. Además se cuantificó el número de lóbulos del estigma en las flores y el número de aréolas en los frutos por conteo directo y se calculó la relación longitud/anchura del fruto. se determinó el volumen (ml) del fruto por el desplazamiento volumétrico de agua en un vaso de precipitado de 100 ml.
Se contaron las semillas por fruto y seleccionaron diez semillas al azar. A cada semilla se le midió la longitud (mm), anchura (mm) y área (mm2) con un analizador de imágenes (Image–Pro Plus versión 3.1, Media Cybernetics, 1997) adaptado a un microscopio estereoscópico olympus SZ–CTV. Además, se calculó la relación longitud/anchura. Se reconocieron 16 caracteres cualitativos (Cuadro 2) y 46 cuantitativos (Cuadro 3) para ser usados en los análisis multivariados.
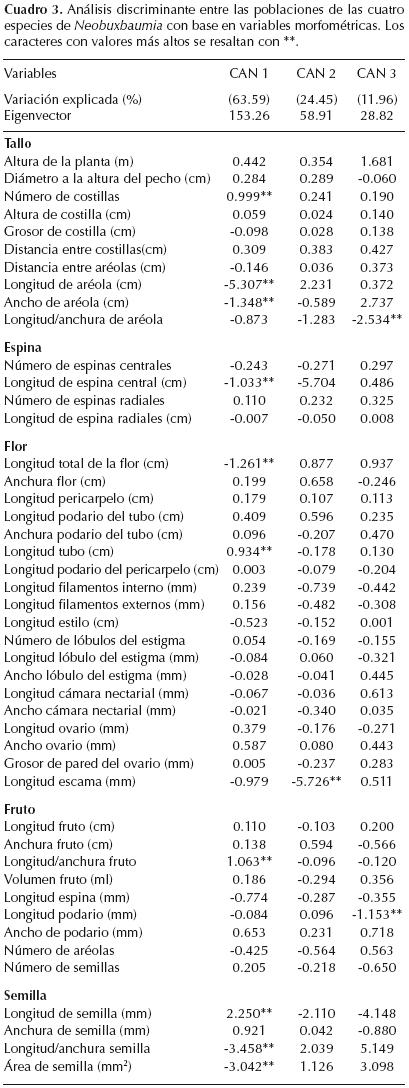
Análisis multivariados. Se calcularon los parámetros de la estadística descriptiva de 46 caracteres comunes a las cuatro especies. Todos los caracteres fueron transformados a logaritmo natural (caracteres continuos) o raíz cuadrada (proporciones) con el propósito de mejorar la distribución normal y que sus varianzas sean homogéneas (Johnson y Wichern, 1982). Los 46 caracteres cuantitativos fueron sometidos a un análisis canónico discriminante (ACD) para identificar el subconjunto de caracteres que separa a las especies. Se aplicó un análisis discriminante clasificatorio con el propósito de corroborar que los individuos están correctamente asignados a las especies identificadas por el análisis canónico discriminante (Johnson y Wichern, 1982). A los 13 caracteres con los valores de carga más alta en las tres primeras funciones canónicas del ACD se les aplicó un análisis de varianza del modelo general lineal, seguido de un análisis de comparación de medias de Tukey para detectar diferencias significativas entre las especies estudiadas. Todos los análisis se realizaron con el paquete estadístico SAS (SAS Institute, 1989).
Se generó una matriz de datos mixta con 16 caracteres cualitativos (Cuadro 2) y los valores de la media o moda de los 13 caracteres cuantitativos seleccionados por el ACD (Cuadro 3) para 15 poblaciones – OTU's (la matriz se puede solicitar a los autores). La matriz se estandarizó en unidades de desviación estándar con el objeto de mantener el principio de igual ponderación entre caracteres (Sneath y Sokal, 1973); con esta matriz se generó la matriz de similitud entre OTU's mediante el coeficiente de Distancia Euclidiana. Este coeficiente conceptualiza la distancia entre dos vectores en el hiperespacio definido por el total de caracteres (Sneath y Sokal, 1973). Como algoritmo de amalgamiento se usó el promedio no ponderado de los pares de grupos UPGMA–unweighted pair–group average–, porque introduce poca distorsión en la distancia entre agrupamientos con respecto a la matriz de similitud (Sneath y Sokal, 1973; Rohlf, 1997).
Resultados
Análisis discriminantes. Las tres primeras funciones canónicas explicaron el 100% de la variación total. El cuadro 3 muestra las 13 variables que presentaron la mayor contribución relativa y que contribuyeron significativamente a la separación de las cuatro especies (λ de Wilks F = 311.81, P < 0.0001, n = 132). El cuadrado de las distancias de Mahalanobis entre los centroides de cada par de especies presentó diferencias significativas (Cuadro 4).
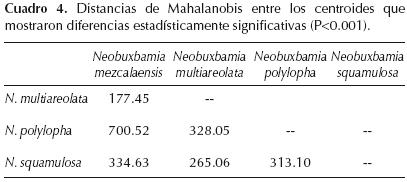
La representación gráfica de las dos primeras funciones canónicas permitió verificar la definición de cuatro grupos (Figura 2). La función canónica 1 separó en el lado derecho de la gráfica a los individuos de las poblaciones de Neobuxbamia polylopha y del lado izquierdo a los individuos de N. mezcalaensis y N. multiareolata. La función canónica 2 permitió separar a los individuos de N. squamulosa de las otras tres especies.
El análisis discriminante clasificatorio mostró que el 100% de los individuos se clasificaron correctamente en sus respectivas especies (Cuadro 5). El análisis de varianza detecto diferencias para los 13 caracteres (P < 0.0001), pero sólo entre algunos pares de especies (Cuadro 6).
Fenograma. El fenograma (Figura 3) corroboró los resultados del análisis canónico discriminante. La separación entre las poblaciones correspondientes a Neobuxbaumia mezcalaensis y N. multiareolata se da a un nivel de corte superior a 1.50 (Figura 3). El primer grupo incluyó las nueve poblaciones de N. mezcalaensis. Los individuos de estas poblaciones tienen en común los siguientes caracteres: costillas jóvenes triangulares agudas, color de tallo verde limón, tricomas en las aréolas jóvenes, espinas radiales con distribución 3–9 (Figura 1A), color blanco–amarillento de las espinas radiales y color de flor blanco–verdoso; los valores más altos para tamaño de aréola, relación longitud/anchura de fruto, longitud y área de la semilla en comparación con las otras tres especies, así como los valores más bajos para la longitud de escama de la flor (Cuadro 6). Santiago Chazumba fue la población más diferente de las nueve estudiadas de N. mez–calaensis (Figuras 3, 4A), ya que posee los individuos con los valores más altos para número de costillas (18), tamaño de aréola (0.63 cm × 0.55 cm), longitud de la espina central (2.71 cm), de la semilla (4.53 mm), relación longitud/anchura de semilla (1.57) y área de la semilla (8.90 mm2) y los valores más bajos para la longitud de la flor (4.00 cm) y del tubo (2.83 cm), en comparación con el resto de las poblaciones.
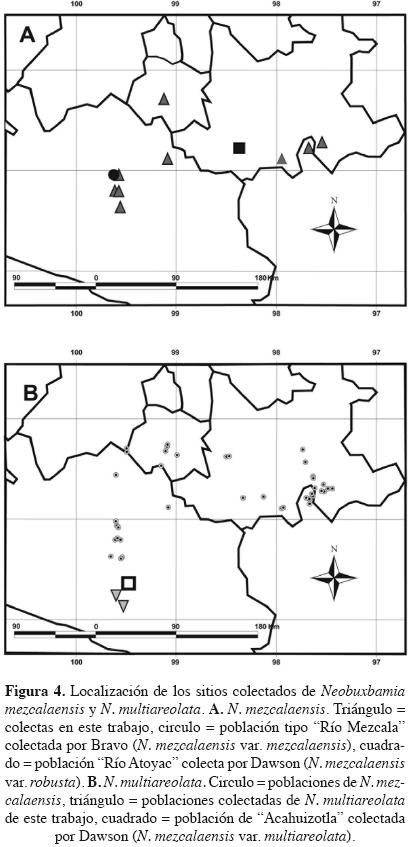
El segundo grupo está formado por las poblaciones de Neobuxbaumia multiareolata (Figura 3). Los individuos de estas poblaciones presentaron una cantidad de tricomas regular en las aréolas jóvenes, espinas radiales con distribución 5–7 (Figura 1B), los valores más bajos para la longitud de la flor, del tubo y la relación longitud/anchura de fruto (Cuadro 6). Las poblaciones de N. squamulosa se separaron por tener tallos que ramifican de color verde grisáceo, flores blancas con escamas papiráceas de 4.7 mm de longitud; mientras que las poblaciones de N. polylopha se agrupan por presentar nectarios extraflorales y carecer de espinas en los frutos, así como por presentar los valores más altos para número de costillas, relación longitud/anchura de aréola, longitud de flor, del podario del fruto y los valores más bajos para tamaño de aréola, de longitud de la semilla y área de semilla (Cuadro 6).
Discusión
Los resultados del ACD permitieron reconocer a las cuatro especies, a partir de 13 caracteres cuantitativos. Adicionalmente un análisis discriminante confirmó que el 100% de los individuos se clasificaron correctamente en sus respectivas especies. Ambos resultados muestran que no hay fenotipos intermedios y que, con base en los caracteres morfométricos, no hay evidencia suficiente de la existencia de subespecies o variedades. En otros estudios con cactáceas, las evidencias morfométricas tampoco apoyan el reconocimiento de categorías infraespecíficas como fue el caso de Astrophytum ornatum (Arias–Montes, 1989), Coryphantha rubustispina (Schmalzen et al., 2004), Escobaría guadalupensis (Baker y Johnson, 2000), Opuntia cantabrigiensis (Muñoz–Urias et al., 2008) y Stenocereus dumortieri (Aguilar–Morales, 2004).
Los 13 caracteres morfométricos con los valores de carga más altos en las tres funciones canónicas permiten discriminar entre las especies estudiadas, por lo que pueden considerarse en su conjunto como diagnósticos. Es importante resaltar que el tamaño y forma de la aréola, el tamaño de la espina central y el patrón de distribución de las espinas radiales permiten separar a Neobuxbaumia mezcalaensis de N. multiareolata, por lo que estos caracteres por sí solos, ayudarán a reconocerlas en ausencia de estructuras reproductivas y se evitará su confusión en el campo.
El fenograma confirmó la separación de las cuatro especies mediante un conjunto de caracteres cualitativos y cuantitativos. Es importante resaltar que Neobuxbaumia multiareolata y N. polylopha resultan más similares morfológicamente que la primera con N. mezcalaensis. La similitud morfológica de las dos primeras especies apoya la propuesta de que son especies hermanas como ya se había sugerido en un análisis filogenético con base en caracteres estructurales (Arias y Terrazas, 2006).
Neobuxbaumia squamulosa es claramente distintiva de las otras especies aquí estudiadas por la presencia de sus tallos ramificados de color verde grisáceo, el patrón de distribución de las espinas radiales > 9, la longitud de las escamas de la flor de 4.7 mm y su consistencia papirácea tanto en las flores como en los frutos. Estos caracteres permitieron reconocer que la población de La Huerta, Jalisco pertenece a N. squamulosa y no a N. mezcalaensis como lo señalan Esparza–Olguín et al. (2002, 2005) y Guzmán et al. (2003). Con base en estos resultados, la distribución de N. mezcalaensis queda restringida a la Depresión del Balsas y al Valle de Tehuacán–Cuicatlán, siempre a altitudes superiores a los 450 m. Seguramente la altitud y los parámetros ambientales asociados a ella juegan o jugaron un papel importante en la diferenciación de estas especies.
La población de Santiago Chazumba es la más disímil entre las poblaciones de Neobuxbaumia mezcalaensis y podría interpretarse como una forma geográfica. No se reconoce como variedad o subespecie de la misma, ya que encontramos que los individuos de esta población son muy variables en sus caracteres vegetativos y no se separan en la representación gráfica del ACD. Tampoco los caracteres morfológicos que la distinguen corresponden a los señalados por Dawson (1948) para N. mezcalaensis var. robusta, seguramente porque únicamente muestreo una población por taxón. Además, la población de Petlalcingo fue más similar en caracteres vegetativos con las poblaciones de Puebla y Morelos. Esta población de Petlalcingo se localiza a 40 km de la población del Río Atoyac (Puebla), de la cual Dawson (1948, 1952) propuso a N. mezcalaensis var. robusta (Figura 4A). Los valores registrados por Dawson para únicamente cuatro caracteres son mayores a los registrados para el promedio de Petlalcingo, por ejemplo: para el tamaño de los individuos (P(=Petlalcingo) = 6 m, 3–12 m, RA(=Río Atoyac) = > 12 m altura y P = 26.1 cm, 17.2–42.3 cm, RA = 39.1 cm dap), la distancia interareolar (P = 1.81 cm, 1.00–3.05 cm, RA = 2.0–2.5 cm) y la altura de la costilla (P = 1.51 cm, 1.00–3.34 cm, RA = > 2.5 cm). Sin embargo, todos los valores caen dentro de la amplitud encontrada para Petlalcingo y de la registrada para la especie. La población de Tierra Colorada (TC) se localiza a 18 km de la población de Acahuizotla de la cual Dawson (1948, 1952) propuso a N. mezcalaensis var. multiareolata. Esta última población se encuentra a 30 km de la población de Zumpango del Río (ZR), la cual es la población más meridional reportada para N. mezcalaensis (Figura 4B). Los tres caracteres (tamaño de los individuos (TC = 3.74 ± 0.08 m, ZR = 7.21 ± 0.10 m), la distancia interareolar (TC = 1.23 ± 0.01 cm, ZR = 1.64 ± 0.02 cm) y la altura de la costilla (TC = 0.86 ± 0.02 cm, ZR = 0.55 ± 0.02), utilizados por Dawson para reconocer a la variedad no se yuxtaponen. A las poblaciones de N. mezcalaensis no se les encuentra en simpatría con las poblaciones de N. multiareolata (Figura 4B). En síntesis, la amplitud de variación encontrada en N. mezcalaensis para sus caracteres morfológicos es amplia y continua, en especial para el tamaño de la planta (altura 2–12 m, χ = 6 m; dap 12.5–42.4 cm, χ = 21.5 cm), la distancia interareolar (0.86–3.05 cm, χ = 1.8 cm) y la altura de la costilla (1.00–3.3, χ = 1.13 cm). Esta amplitud engloba a la variación registrada por Dawson (1948) para sus tres variedades: N. mezcalaensis var. mezcalaensis, N. mezcalaensis var. multiareolata y N. mezcalaensis var. robusta. Ninguno de estos cuatro caracteres fue resaltado por los análisis practicados para diferenciar a las especies estudiadas.
Con base en los resultados de esta investigación se apoya el reconocimiento de Neobuxbaumia mezcalaensis y de N. multiareolata como entidades independientes como ya habían señalado otros autores (Bravo–Hollis et al., 1972; Bravo–Hollis, 1978; Anderson, 2001; Guzmán et al., 2003; Flores–Ortiz, 2004). Los análisis estadísticos aquí realizados corroboran que la cantidad de tricomas, la longitud de la flor y del tubo, así como el tamaño de las semillas son importantes para apoyar la separación de estas especies. Además, se incorporaron por primera vez para segregar a estos dos taxones, el tamaño de la aréola, el patrón de distribución de las espinas radiales en la aréola, el tamaño de la espina central, de la escama de la flor, del podario del fruto, la relación longitud/anchura de fruto (forma del fruto) y la forma y área de la semilla. En consecuencia, nuestros resultados no apoyan la propuesta de Dawson (1948, 1952) de reconocer tres variedades dentro de N. mezcalaensis, como tampoco la de Hunt (2006) de incluir a N. multiareolata y N. mezcalaensis en una especie. Por lo tanto aquí se presenta una clave para las cuatro especies de Neobuxbaumia y una lámina (Figura 5) que ilustra aspectos de los tallos, aréolas, flores y frutos de las especies estudiadas. Concluimos que los análisis permitieron encontrar caracteres que no se habían señalado como importantes para distinguir estas dos especies, en particular algunos caracteres vegetativos permitirán su reconocimiento en campo sin estructuras reproductivas. El método de análisis de la posición de las espinas radiales y la central fue de utilidad pues el patrón de arreglo particular para cada especie permitió distinguir gráficamente entre ellas. Al ser un método repetible y objetivo de asignar un patrón para cada especie, sugerimos emplearlo en estudios de otras cactáceas.
Clave para las cuatro especies de Neobuxbaumia
1. Plantas ramificadas (2–12 ramas), tallo color verde grisáceo; flores y frutos con escamas papiráceas................................................................................................N. squamulosa
1. Plantas no ramificadas (monopódicas), tallo color verde claro a oscuro; flores y frutos con escamas no papiráceas.
2. Número de costillas > 25, aréolas 0.26 × 0.19 cm, espinas amarillo ocre; nectarios extraflorales presentes; semillas < 3.0 mm de largo con un área < 4.5 mm2....................................................................N. polylopha
2. Número de costillas < 25, aréolas > 0.27 × 0.20 cm, espinas blancas o rojas; sin nectarios extraflorales; semillas > 3.0 mm de largo con un área > 4.5 mm2.
3. Aréolas de 0.37 × 0.32 cm, espinas radiales con distribución 5–7; semillas ovoides de 3.25 mm de largo con área de 6.56 mm2...............N. multiareolata
3. Aréolas de 0.46 × 0.40 cm, espinas radiales con distribución 3–9; semillas ovoides de 3.78 mm de largo con área de 8.94 mm2................N. mezcalaensis
Agradecimientos
Se agradece al Consejo Nacional de Ciencia y Tecnología por el apoyo al proyecto (33064–V a TT) y por la beca otorgada al primer autor para realizar estudios de posgrado (190345). Agradecemos a Cesario Catalán, Leobardo Sánchez, Dorismilda Martínez, Loraine Matias y al personal de la Reserva de la Biosfera Barranca de Meztitlán por el apoyo brindado en el trabajo de campo; a Ligia Esparza la disponibilidad de algunas semillas y a Julio César Montero por el trabajo gráfico.
Literatura citada
Aguilar–Morales I. 2004. Variación morfológica de Stenocereus dumortieri (Cactaceae). Tesis licenciatura, Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D.F [ Links ]
Anderson E.F. 2001. The Cactus Family. Timber Press, Portland. [ Links ]
Arellano E. y Casas A. 2003. Morphological variation and domestication of Escontria chiotilla (Cactaceae) under silvicultural management in the Tehuacán Valley, Central Mexico. Genetic Resources and Crop Evolution 50:439–453. [ Links ]
Arias–Montes S. 1989. Variación morfológica de Astrophytum ornatum (DC.) Web. (Cactaceae) en cuatro poblaciones de las zonas áridas Queretana e Hidalguense. Tesis licenciatura, Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D.F. 75 pp. [ Links ]
Arias S., Terrazas T. y Cameron K. 2003. Phylogenetic analysis of Pachycereus (Cactaceae, Pachycereeae) based on chloroplast and nuclear DNA sequences. Systematic Botany 28:547–557. [ Links ]
Arias S. y Terrazas T. 2006. Análisis cladístico del género Pachycereus (Cactaceae) con caracteres morfológicos. Brittonia 58:197–216. [ Links ]
Arroyo–Cosultchi G., Terrazas T., Arias S. y López–Mata L. 2007. Morfología de la semilla de Neobuxbaumia (Cactaceae). Boletín de la Sociedad Botánica de México 81:17–25. [ Links ]
Baker M.A. 2006. A new florally dimorphic hexaploid, Echinocereus yavapaiensis sp. nov. (section Triglochidiatus, Cactaceae) from central Arizona. Plant Systematic and Evolution 258:63–83 [ Links ]
Baker M.A. y Pinkava D.K. 1987. A cytological and morphometric analysis of a triploid apomict, Opuntia xKelvinensis (subgenus Cylindropuntia, Cactaceae). Brittonia 39:387–401. [ Links ]
Baker M.A. y Johnson R.A. 2000. Morphometric analysis of Escobaria sneedii var. sneedii, E. sneedii var. leei, and E. guadalupensis (Cactaceae). Systematic Botany 25:577–587. [ Links ]
Barthlott W. y Hunt D.R. 1993. Cactaceae. En: Kubitzki K., Ro–hwer J.G. y Bittrich V. Eds. The Families and Genera of Vascular Plants, pp. 161–197, Springer–Verlag, Berlin. [ Links ]
Bravo–Hollis H. 1932. Contribución al conocimiento de las cactáceas del estado de Guerrero. Cactáceas del cañón del Zopilote. Anales del Instituto de Biología 3:375–398. [ Links ]
Bravo–Hollis H. 1978. Las Cactáceas de México. Vol. 1. Universidad Nacional Autónoma de México. México, D.F. [ Links ]
Bravo–Hollis H., Scheinvar L. y Sánchez–Mejorada H. 1970. Estudio comparativo del género Neobuxbaumia, Neobuxbaumia polylopha. Cactáceas y Suculentas Mexicanas 15:75–82. [ Links ]
Bravo–Hollis H., Scheinvar L. y Sánchez–Mejorada H. 1971a. Estudio comparativo del género Neobuxbaumia Backberg. II Neobuxbaumia macrocephala (Web) Daws. Cactáceas y Suculentas Mexicanas 16:3–14. [ Links ]
Bravo–Hollis H., Scheinvar L. y Sánchez–Mejorada H. 1971b. Estudio comparativo del género Neobuxbaumia Backberg. III Neobuxbaumia scoparia (Poselg) Backberg. Cactáceas y Suculentas Mexicanas 16:75–83. [ Links ]
Bravo–Hollis H., Scheinvar L. y Sánchez–Mejorada H. 1972. Estudio comparativo del género Neobuxbaumia Backberg. IV Neobuxbaumia multiareolata. Cactáceas y Suculentas Mexicanas 18:59–67. [ Links ]
Buxbaum, F. 1950. Morphology of Cacti. Section I. Roots and Stems. Abbey Garden Press, Pasadena. [ Links ]
Casas A., Caballero J., Valiente–Banuet A., Soriano J.A., y Dávila P 1999. Morphological variation and the process of domestication of Stenocereus stellatus (Cactaceae) in central México. American Journal of Botany 86:522–533. [ Links ]
Chamberland M. 1997. Systematics of the Echinocactus polycephalus complex (Cactaceae). Systematic Botany 22:303–313. [ Links ]
Cruz M. y Casas A. 2002. Morphological variation and reproductive biology of Polaskia chende (Cactaceae) under domestication in Central Mexico. Journal of Arid Environments 51:561–576. [ Links ]
Dawson E.Y. 1948. New cacti of southern Mexico. Allan Hancock Foundation Publications Occasional Papers 1:1–69. [ Links ]
Dawson E.Y. 1952. Notes on Neobuxbaumia. Cactus and Succulent Journal (Los Angeles) 24:167–173. [ Links ]
De Luna E. y Gómez–Velasco M. 2008. Morphometrics and identification of Braunia andrieuxii and B. secunda (Hedwigiaceae, Bryopsida). Systematic Botany 33:219–228. [ Links ]
Esparza–Olguin L.G., Valverde T. y Vilchis–Anaya E. 2002. Demographic analysis of a rare columnar cactus (Neobuxbaumia macrocephala) in the Tehuacan Valley, Mexico. Biological Conservation 103:349–359. [ Links ]
Esparza–Olguín L.G., Valverde T. y Mandujano M.C. 2005. Comparative demographic analysis of three Neobuxbaumia species (Cactaceae) with differing degree of rarity. Population Ecology 47:229–245. [ Links ]
Evans L.S. 2005. Stem surface injuries of Neobuxbaumia tetetzo and Neobuxbaumia mezcalaensis of the Tehuacan Valley of central Mexico. Journal of the Torrey Botanical Society 132:33–37. [ Links ]
Flores–Ortiz C.M. 2004. Quimiotaxonomía del género Neobuxbaumia. Tesis doctoral, Facultad de Estudios Superiores Iztacala,, Universidad Nacional Autónoma de México. México, D.F. 112 pp. [ Links ]
Flores–Ortiz C.M., Dávila P. y Portilla L.B.H. 2003. Alkaloids from Neobuxbaumia species (Cactaceae). Biochemical, Systematics and Ecology 31:581–585. [ Links ]
Fuentes–Pérez M. 2004. Anatomía floral de algunas especies de la tribu Pachycereeae (Cactaceae). Tesis licenciatura, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México. México, Los Reyes Iztacala. 119 pp. [ Links ]
Godínez–Alvarez H. y Valiente–Banuet A. 2004. Demography of the columnar cactus Neobuxbaumia macrocephala: a comparative approach using population projection matrices. Plant Ecology 174:109–118. [ Links ]
González–Andrés F. y Ortiz J.M. 1996. Morphometrical characterization of cytisus and allies (Genisteae: Leguminosae) as an aid taxonomic discrimination. Israel Journal of Plant Science 44:95–114. [ Links ]
Grimaldo–Juárez O., Terrazas T., García–Velásquez A., Cruz–Villagas M. y Ponce–Medina J.F. 2007. Morphometric analysis of 21 pitahaya (Hylocereus undatus) genotypes. Journal of the Professional Association for Cactus Development 9:99–117. [ Links ]
Guzmán–Cruz U., Arias S. y Dávila PD. 2003. Catálogo de Cactáceas Mexicanas. Universidad Nacional Autónoma de México y Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México, D.F. [ Links ]
Henderson, A. 2002. Phenetic and phylogenetic analysis of Reinhardtia (Palmae). American Journal of Botany 89:1491–1502. [ Links ]
Hunt D.R. 1999. CITES. Cactaceae Checklist. Royal Botanic Gardens Kew and international Organization for Succulent Plant Study. Remous Limited, Milborne Port, Reino Unido. [ Links ]
Hunt D.R. 2006. The New Cactus Lexicon. International Cactaceae Systematics Group, Milborne Port, Reino Unido. [ Links ]
Janovec J.P. y Harrison J.S. 2002. A morphological analysis of the Compsoneura sprucei complex (Myristicaceae), with a new combination for the Central American species Compsoneura mexicana. Systematic Botany 27:662–673. [ Links ]
Johnson R.A. y Wichern D.W. 1982. Applied Multivariate Statistical Analysis. Prentice Hall, Englewood Cliffs. [ Links ]
Kephart S., Sturgeon K., Lun J. y Bledsoe K. 1999. Varietal relationships in Silene douglasii (Caryophyllaceae): morphological variability at the population level. Systematic Botany 24:529549. [ Links ]
Lau A.B. 1994. Neobuxbaumia sanchezmejoradae Lau. Cactáceas y Suculentas Mexicanas 39:3–7. [ Links ]
Levin G.A. 1998. Evolution in the Acalypha gracilens/monococca complex (Euphorbiaceae): morphological analysis. Systematic Botany 23:269–287. [ Links ]
Loza–Cornejo S. y Terrazas T. 2003. Epidermal and hypodermal characteristics in North American Cactoideae (Cactaceae). Journal of Plant Research 116:27–35. [ Links ]
Loza–Cornejo S., López–Mata, L. Terrazas, T. 2008. Morphological seed traits and germination of six species of Pachycereeae (Cactaceae). Journal of the Professional Association for Cactus Development 10:71–84. [ Links ]
Media Cybernetics. 1997. Image Pro Plus, Reference Guide. Version 3.0 for Windows. Silver Spring, Maryland. [ Links ]
Muñoz–Urias A., Palomino–Hasbach G., Terrazas T., García–Velázquez A. y Pimienta–Barrios E. 2008. Variación anatómica y morfológica en especies y entre poblaciones de opuntia en la porción sur del Desierto Chihuahuense. Boletín de la Sociedad Botánica de México 83:1–11. [ Links ]
Ramírez–Padilla C.A. y Valverde T. 2005. Germination responses of three congeneric cactus species (Neobuxbaumia) with differing degrees of rarity. Journal of Arid Environments 61:333–343. [ Links ]
Rivera–Montoya I. 2003. Estudio de la variabilidad genética de la cactácea columnar Neobuxbaumia mezcalaensis. Tesis licenciatura, Facultad de Estudios Superiores Iztacala, Universidad Nacional Autónoma de México. México, Los Reyes Iztacala. 84 pp. [ Links ]
Rohlf F.J. 1997. NTSYSpc. Numerical Taxonomy and Multivariate Analysis System. Version 2.0. Exeter Software. Setauket, Nueva York. [ Links ]
Ruedas M., Valverde T. y Zavala–Hurtado J.A. 2006. Analysis of the factors that affect the distribution and abundance of three Neobuxbaumia species (Cactaceae) that differ in their degree of rarity. Acta Oecologica 29:155–164. [ Links ]
SAS Institute. 1989. SAS/STAT User's Guide, Release 6.03 Edition. SAS Institute Inc., Cary. [ Links ]
Scheinvar L.J.A. y Sánchez–Mejorada H. 1990. Neobuxbaumia squamulosa. Scheinvar et Sánchez Mejorada sp. nov. Cactáceas y Suculentas Mexicanas. 35: 13–18. [ Links ]
Schmalzen R.J., Nixon R.T., Best A.L. y Tress J.A. Jr. 2004. Morphometric variation in Coryphantha robustispina (Cactaceae). Systematic Botany 29:553–568. [ Links ]
Sneath P.H y Sokal R.R. 1973. Numerical Taxonomy. The Principles and Practices of Numerical Classification. W.H. Freeman and Company, San Francisco. [ Links ]
Terrazas T. 2000. Occurrence of perforated ray cells in genera of Pachycereeae (Cactaceae). International Association of Wood Anatomists Journal 21:457–462. [ Links ]
Valiente–Banuet A., Arizmendi M.C., Rojas–Martinez A. y Domínguez–Canseco L. 1996. Ecological relationships between columnar cacti and nectar feeding bats in Mexico. Journal of Tropical Ecology 12:103–119. [ Links ]
Valiente–Banuet A., Rojas–Martínez A. Arizmendi M.C., y Dávila P 1997. Pollination biology of two columnar cacti (Neobuxbaumia mezcalaensis and Neobuxbaumia macrocephala) in the Tehuacan Valley, Central Mexico. American Journal of Botany 84: 452–455. [ Links ]

