










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.20 no.1 Ciudad de México abr. 2010
Artículos
Non–selective in situ grazing of diatoms by juvenile Green Abalone (Haliotis fulgens Philippi, 1945) in the Baja California Peninsula
Consumo in situ no selectivo de diatomeas por juveniles de abulón azul (Haliotis fulgens Philippi, 1945) en la Península de Baja California
Uri Argumedo Hernández1 and David A. Siqueiros–Beltrones2,*
1 Dpto. Economía, Universidad Autónoma de Baja California Sur. A. P. 19–B. La Paz, B. C. S., C. P. 23000. México.
2 Becario COFAA y EDI del IPN. Dpto. Plancton y Ecología Marina, Centro Interdisciplinario de Ciencias Marinas, IPN. A. P. 592. La Paz, B. C. S., México. *E–mail: dsiquei@ipn.mx
Recibido: 23 de marzo de 2009.
Aceptado: 21 de enero de 2010.
ABSTRACT
For experimental or rearing purposes, young abalones are fed benthic diatoms successfully during their early life stages without considering if the diatoms provided are found in the abalone habitat, nor if grazing selectivity exists. Our aim in this study was to gather further evidence that would indicate whether young Haliotis fulgens graze diatoms in situ selectively. Thirty–four specimens of young H. fulgens were collected in a fishing site in BCS, México on two dates. We identified the diatoms in their gut contents (144 species) and determined their relative abundances. Fourteen diatoms (80% of the total abundance) were considered most important and twelve were among the most important in the diet of young abalone in BCS. The diversity values (H') of the diatoms in their guts ranged from 1.05 to 4.47, depending on dominance, which reflected the structure of diatom assemblages in situ. The above observations and the low similarity measured between samples show that species richness and equitability are similar even if the species composition of diatoms ingested by young abalone differs between specimens. The results lead us to conclude that grazing of diatoms by young H. fulgens is non–selective, and that the structure of diatom assemblages is more important than the particular taxa in their diet.
Key words: Diatoms, abalone, in situ grazing, diversity, selectivity.
RESUMEN
Durante sus etapas tempranas de vida los abulones jóvenes son alimentados exitosamente con diatomeas bentónicas sin considerar si las especies suministradas se hallan en su medio natural, o si existe algún tipo de preferencia o selectividad por ciertas diatomeas. El objetivo de este estudio fue recabar evidencias acerca de si los juveniles de Haliotis fulgens Philippi presentan un consumo selectivo de diatomeas cuando se alimentan in situ. Se recolectaron treinta y cuatro juveniles de H. fulgens en un sitio de pesca de BCS, México en dos fechas. Se disecaron y se examinaron contenidos intestinales. Se identificaron 144 taxa de diatomeas y se determinaron sus abundancias relativas; 42% fueron epifitas. Catorce taxa representaron el 80% de la abundancia total; de estos 12 han sido propuestos como los más importantes en la dieta de juveniles de abulón en la costa W de BCS. Los valores de diversidad (H') variaron entre 1.05 y 4.47 y dependieron más de la dominancia que de la riqueza de especies, reflejando la diversidad de diatomeas en el hábitat. Estas observaciones y la baja similitud medida entre contenidos intestinales, muestran que la riqueza y diversidad de especies de diatomeas ingeridas por juveniles de abulón serán similares aun cuando la composición de especies de diatomeas sea distinta entre especímenes. Esto nos permite concluir que la ingestión de diatomeas por juveniles de abulón es no selectiva y que la estructura de las asociaciones de diatomeas es más importante que los taxa específicos en su dieta.
Palabras clave: Diatomeas, abulón, consumo in situ, diversidad, selectividad.
INTRODUCTION
In México, as well as in other countries, natural populations of abalone (Haliotis spp.) have been much depleted. Thus, research on abalone biology and ecology has been focused on the recovery of abalone fisheries by proposing an aquaculture approach to the problem (Searcy–Bernal & Salas–Garza, 1990). However, it has been noted that much of the aquaculture practice concerning feeding of abalone, particularly juveniles, is highly empirical, and that its in situ feeding habits have not been well documented (Siqueiros Beltrones et al., 2004).
During early life stages, cultured abalones are successfully fed benthic diatoms, whether for experimental or rearing purposes. However, this practice does not require that the diatoms provided occur naturally in their natural habitat, nor it matters if young abalones show preference for any particular diatom species.
Updated inventories indicate that many species of benthic diatoms, which represent a food source for abalones during their early stages, occur in their rocky environment (Siqueiros–Beltrones & Valenzuela–Romero, 2004; Siqueiros–Beltrones et al., 2004 Siqueiros–Beltrones et al., 2005). In Thailand, Haliotis asinina thrives in a habitat (coral reef) where few small macroalgae exist (Cladophora sp., Centroceras sp. and Ectocarpussp.); thus H. asinina has to feed mostly on benthic diatoms throughout its life (Sawatpeera et al., 1998). However, most studies on abalone diet in their natural environment examine the gut contents of adult specimens (Serviere–Zaragoza et al., 1998), which feed mostly on macroalgae (Tutschulte & Connell, 1988). Consequently, few studies have focused on the in situ diet of young abalone whether it is macroalga based (Guzmán del Próo et al., 2003) or diatom based (Siqueiros–Beltrones et al., 2004). Recently gut content analyses (size 10 to 100mm) of Haliotis fulgens Philippi and H. corrugata Wood in BCS, México showed up to 9 species of macroalgae and very few diatoms in their diet (Guzmán del Próo et al., 2003. However, the reexamined samples revealed that abundant diatoms were present. This observation prompted the examination of young specimens of H. fulgens and H. corrugata from the same locality. As a result 98 diatom taxa were recovered from the gut contents, mostly epiphytes, out of which 37 showed to be numerically more important in their diet (Siqueiros–Beltrones et al., 2005). The low intra– and inter–specific similarity of diatom taxa measured in the gut contents from the last study posed no evidence of in situ selective grazing on diatoms by young abalone. The aim of this study was to gather further evidence for determining if young Haliotis fulgens feeding on diatoms is non–discriminative in their natural environment. (Figure 1)
Because our null hypothesis (Ho) stated that diatom feeding by young H. fulgens in their natural habitat is non–selective it required supportive evidences; thus, the following subordinate hypotheses were contrasted: 1) Abalone gut contents will show high species richness and diversity of ingested diatoms as do the assemblages in the wild, thus reflecting non–discrimination in grazing; 2) Species composition of diatoms in the gut contents will differ between abalone specimens and likewise from the substrate samples because of the patchy distribution of benthic diatom assemblages; this should be reflected in low similarity index values.
MATERIAL AND METHODS
Juvenile specimens of H. fulgens were collected at Clam Bay, an abalone fishing site inside a small cove located 6 km SE of Bahía Tortugas. Maximum depth is measured at 6 m and is protected from wave action and NW winds. The specimens were collected at wadding depth during low tide by surveying the rocky beach. Due to abalone scarcity, the search for specimens demanded a thorough examination of rocks exposing the underside and retrieving them with a spatula. In this manner 34 specimens (16–74 mm in length) collected in November 2003 (27) and in June 2004 (7) were preserved in 70% ethanol.
We dissected the specimens under a stereoscope and removed their guts. The gut contents of each specimen (sample) were extracted by holding one extreme in place while pressing with a dissecting needle length–wise. Gut contents were then transferred with a pipette into a test tube. To oxidize, the organic matter of each sample was treated with a mixture of commercial alcohol and nitric acid at 1:3:5 (Siqueiros–Beltrones & Voltolina, 2000); these proportions varied according to the organic matter in each sample. The oxidized material was rinsed with distilled water to a pH > 6. The clean diatom frustules were mounted on two permanent slides per sample using Pleurax.
Diatoms were identified using scientific reports from studies made in similar localities in the region (Siqueiros–Beltrones, 2002; Siqueiros–Beltrones et al., 2004, 2005; Witkowski et al., 2000). The relative abundances for all diatom taxa in each sample were estimated (n = 500, Siqueiros–Beltrones, 2002).
To contrast our Ho, evidence was gathered based on the analyses of species composition and structure of diatom assemblages found in the gut contents of the collected specimens. Species diversity and equitability were estimated using Shannon's H' and Pielou's J', along with Simpson's 1–λ (Brower & Zar, 1984). Similarity between gut contents was measured on the basis of presence/absence of taxa with Jaccard index (Magurran, 1988), and also considering their relative abundances using Bray–Curtis index (Brower & Zar, 1984). In a first approach, we included all taxa in the analyses and constructed the dendrograms by simple union. In a second approach, only the most abundant taxa were considered, i.e., those representing 80% of the total abundance (N). Also, following Siqueiros–Beltrones et al. (2005), similarity was measured between diatom assemblages in the gut contents of the June specimens and from a rock.
RESULTS
Overall, 144 diatom taxa were identified as mostly epiphytes, 89 taxa in June 2004 and 130 taxa in November 2003. Lower species richness (S) in June samples could relate to smaller sample size. Over 80% of total abundance (N) was represented by only 14 taxa of epiphytic diatoms (Table 1). Table 2 shows how the distribution of the relative abundances among the taxa determined the structure of the assemblages. High dominance values in certain samples were due to the high numbers of Cocconeis californica var. californica Witkowski et al., Eunotogramma sp. 1 and Rhoicosphenia genuflexa (Kützing) Medlin; these are epiphytic forms that reached up to 50% of the total abundances. Diversity values were high in general (H'mean = 3.3) although ranging from 1.05 to 4.47, while H'max values reflected the highly variable species richness among the samples from 3.6 (S=12) to 5.6 (S=48). In agreement with our H1, high diversity values are supportive of (Ho) non–selective grazing of diatoms by juvenile H. fulgens.
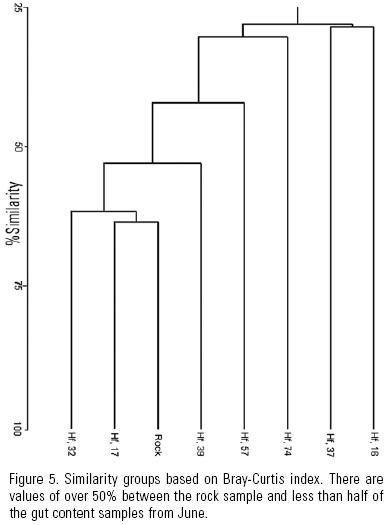
Similarity between the gut contents of all the specimens measured both with the Jaccard and Bray–Curtis indices was very low, and dendrograms showed no group formation even though the shared presence of dominant taxa should have promoted it (Fig. 2). This may also indicate that grazing of diatom taxa was indiscriminate in accordance with H1. The similarity analysis based on the 14 most abundant taxa, yielded (high) values of 75 to 93% (Fig. 3) with the Jaccard index, while the Bray–Curtis yielded a wider range of variation (32 to 83%) due to the difference in proportional representation of the same (common and dominant) diatom taxa in different samples (Fig. 4). This relation usually reads the other way around in the derived dendrograms when comparing Jaccard and Bray–Curtis indices. Moreover, similarity between the gut contents of the June specimens and the available diatom flora from a rock substratum (89 taxa) based on Jaccard's similarity was very low (>50%). Although this could indicate selective feeding by the abalone, the difference in S is an important factor.
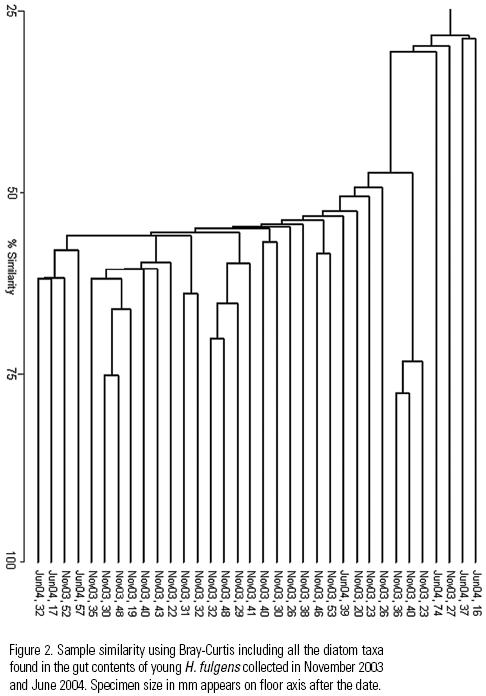
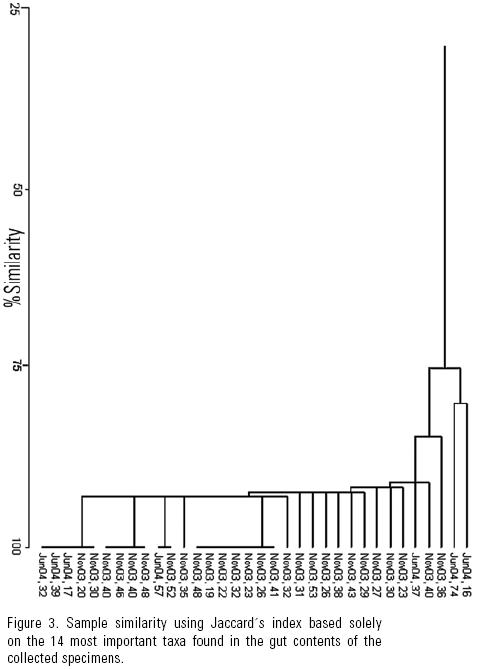
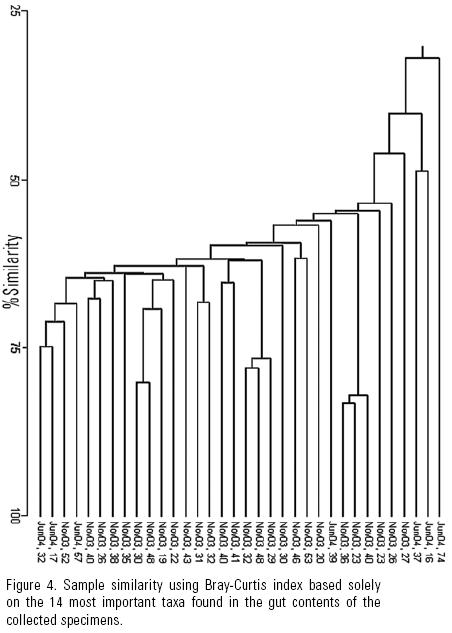
The Bray–Curtis index indicated certain similarity between less than half the specimens. This is attributed to the presence of different important taxa in the gut contents of each abalone specimen (Figure 5). Thus, high similarity in the presence/absence of taxa and high S lead us to infer no diatom taxa are preferred by juvenile H. fulgens, and that the importance of certain diatom taxa in their gut contents detected by the Bray–Curtis index depends on the patchy distribution of diatoms, that is, on whatever taxon is abundant in a given diatom patch. This agrees with our hypothesis that the young of green abalone feed non–selectively on diatoms in the wild.
DISCUSSION
High species richness of the diatoms found in the gut contents of H. fulgens juveniles further supports the proposal that they represent an important component of their diet (Siqueiros–Beltrones et al., 2005), which has seldom been considered as such for young and adult haliotids as opposed to what is accepted for postlarva abalone (Siqueiros–Beltrones & Voltolina, 2000). Moreover, epiphytic diatom represented 42% of the total species richness. Considering that juvenile abalone feed largely on macroalgae, the abundance of diatoms (epiphytes) shows their particular importance in their diet. Moreover, Haliotis asinina thrives in a habitat (coral reef) in Thailand where few small macroalgae exist (Cladophora sp., Centroceras sp. and Ectocarpus sp.); thus, it has to feed mostly on benthic diatoms throughout its life (Sawatpeera et al., 1998), whether these are epiphytes or not. Several diatom taxa that may be considered important in the diet of young and adult abalone occur commonly on rocky substrata. But the few in situ studies available have noted the importance of only certain (although imprecisely identified) diatoms (Cocconeis spp.) in the diet of postlarva abalone, specifically on coralline algae covering the rock (Daume et al., 1997, 1999; Kawamura et al., 1998).
However, in the central region of the Baja California Peninsula 42 species of macroalgae associated to rocks, which serve as refuge and food for Haliotis spp., have been identified (Carreón–Palau, 2000). Thus, epiphytic diatom forms are highly available, and young abalone feed commonly on macroalgae surfaces (pers. obs.).
Most diatom taxa in the present study are epiphytes, where 95% of the diatom total abundance is represented by 38 taxa, very similar to what Siqueiros–Beltrones et al. (2005) reported. In both studies Cocconeis speciosa, Gomphoseptatum aestuari, Gomphonemopsis pseudoexigua, Cocconeis costata var. hexagona, Grammatophora hamulifera, Cocconeis diminuta, Tabularia investiens, Cocconeis costata var. pacifica, Rhoicosphenia adolphi, Campylopyxis garkeana, Grammatophora marina, and Cocconeis scutellum (Table 1), whose nutritional contribution should be assessed, stood out in the gut contents of young abalone (H. fulgens and H. corrugata), and are thus considered a natural component of their diet in the Pacific coast of Baja California Sur, Mexico.
The observed species richness (S) was higher (46 more taxa) than the one reported from H. corrugata and H. fulgens gut contents by Siqueiros–Beltrones et al. (2005). Although these authors examined 12 specimens (98 taxa), our June sample size was only 7, yielding 89 taxa; nonetheless, it shows that a correspondence does exist between S and the number of abalone specimens examined. However, collection of young abalone in the wild poses great difficulty because of their scarceness and legal restrictions.
The structure analysis of the diatom samples showed that few abundant taxa and many rare or uncommon taxa characterized the diatom assemblages in the gut contents, and 5% of the total abundance included over 100 diatom taxa. Can these be considered accompanying flora of lesser importance in the diet of H. fulgens juveniles during the studied seasons? Or, is this diversity required for their optimal growth? This could explain why young abalone grow faster in the wild (Guzmán del Próo et al., 2004).
Our observations agree with Siqueiros Beltrones et al. (2005), who indicated that only a tenth of the diatom taxa in the gut contents of young abalone are abundant (important), suggesting that these might be selected during grazing. However, our interpretation is that the importance of certain diatom taxa in the gut contents of juvenile H. fulgens depends not on selectivity but is more dependent on non–discriminative grazing biased by the (patchy) distribution of diatoms in the wild. This is what the similarity dendrograms from this and previous studies (Siqueiros Beltrones et al., 2005) show, i.e., very low similarity between the gut contents of abalone specimens from the same or different species or with the surrounding flora. Thus, whatever diatom species is abundant in a certain diatom patch will be better represented in the ingested material and, according to recent diatom species inventories (Siqueiros Beltrones & Valenzuela–Romero, 2004; Siqueiros–Beltrones et al., 2005), it is likely that many other diatom taxa turn out to be important constituents of young abalone diet depending on the time of the year, considering that the total inventory of diatom taxa in the abalone habitat reaches over 320 taxa (Siqueiros–Beltrones et al., 2004). However, certain selectivity may occur due to differential ability of young abalone to digest the ingested diatoms, where smaller forms may pass their digestive system intact (Kawamura et al., 1998); or break the diatom walls with their radula (Roberts et al., 1999), which is related to the different degree of silicification of the available diatoms.
The above warns on the complexity of the relationships between young abalone and diatoms, which may surpass the implications of our hypothesis (even though it could not be refuted) and require higher resolution analyses. However, the range of variation of the diversity measurements similar to that reported by Siqueiros Beltrones et al. (2005), permits us to infer that the structure of diatom assemblages that serve as food for young Haliotis spp. is more important than the particular taxa involved. High diversity of diatoms in the gut contents reflects high diversity of diatom assemblages in situ. The low values of H' observed might reflect grazing on monospecific patches of certain taxa, which result also in high dominance values (λ). In general, diversity values are high, supported by corresponding values of J' (and low dominance) indicating that diatom taxa are represented homogeneously. The H' values for benthic diatom assemblages with a stable structure generally range between 1 < H' < 5 (Siqueiros–Beltrones, 2005). These variations may represent succession stages and reflect a heterogeneous distribution whose permanence shows its stability far from an improbable Hmax under normal conditions (Siqueiros–Beltrones, 2005). Such stability appears to favor many grazing species found in rocky habitat, including abalone.
The results of this study may be used as a reference when choosing a better diet for abalone juveniles under culture conditions for rearing or research purposes, whether looking for a particular species or an assemblage of those present in the abalone habitat or in their diet.
ACKNOWLEDGEMENTS
Financial support was granted by Consejo Nacional de Ciencia y Tecnología (project: 2002/40778) and Instituto Politécnico Nacional (project CGPI–20030211). We also acknowledge support by the Sociedad Cooperativa Bahía Tortugas, Omar López, Guillermina Valenzuela, and Oscar Hernández Almeida for field work. We thank Domenico Voltolina for his commentaries to a former manuscript. Diana Dorantes Fisher edited the final English version. We also thank two anonymous referees for their helpful commentaries.
REFERENCES
Brower, J.E. & J.H. Zar. 1984. Field and laboratory methods for general ecology. Wm. C. Brown Co. Dubuque, Iowa, U.S.A. 226 p. [ Links ]
Carreón–Palau, L. 2000. Microhábitat, distribución y abundancia de juveniles de abulón, Haliotis fulgens y H. corrugataen Bahía Tortugas B.C.S., México. MSc thesis. Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. México. 79 p. [ Links ]
Daume, S., S. Brand & W. Woelkerling. 1997. Effects of post–larval abalone (Haliotis rubra) grazing on the epiphytic diatom assemblage of coralline red algae. Molluscan Research 18: 119–130. [ Links ]
Daume, S., S. Brand–Gardner & W. Woelkerling. 1999. Preferential settlement of abalone larvae: Diatom films vs. nongeniculate coralline red algae. Aquaculture 174: 243–254. [ Links ]
Guzmán del Próo, S., E. Serviere–Zaragoza & D. Siqueiros–Beltrones. 2003. Natural Diet of Juvenile Abalone Haliotis fulgens and Haliotis corrugata (Mollusca:Gastropoda) in Bahía Tortugas, Mexico. Pacific Science 57 (3): 319–324. [ Links ]
Guzmán del Próo, S., J. Carrillo–Laguna, J. Belmar–Pérez, L. Carreón–Palau & A. Castro. 2004. Transplanting of wild and cultivated juveniles of green abalone (Haliotis fulgens Philippi 1845): growth and survival. Journal of Shellfish Research 2 (3): 855–860. [ Links ]
Kawamura, T., D.R. Roberts & C.M. Nicholson. 1998. Factors affecting the food value of diatom strains for post–larval abalone Halioits iris. Aquaculture 160: 81–88. [ Links ]
Magurran, A.E. 1988. Ecological diversity and its measurement. Princeton University Press. N. J. 179 p. [ Links ]
Roberts, R.D., T. Kawamura & C.M. Nicholson. 1999. Growth and survival of postlarval abalone (Haliotis iris) in relation to development and diatom diet. Journal Shellfish Research 18 (1): 243–250. [ Links ]
Sawatpeerra, S., E.S. Upatham, M. Kruatrachue, V. Ingsrisawang, T. Singhagraiwan, Y.P. Chitramvong & K. Parkpoomkamol. 1998. Determination of gut contents of thai abalone Halioitis asinina Linnaeus. Journal Shellfish Research 17 (3): 765–769. [ Links ]
Searcy–Bernal, R. & A. E. Salas–Garza . 1990. Investigaciones sobre el cultivo del abulón en la Universidad Autónoma de Baja California. Serie científica Universidad Autónoma de Baja California Sur, No. Especial 1: 44–50. [ Links ]
Serviere–Zaragoza, E., D. Gómez–López & G. Ponce–Díaz. 1998. The natural diet of green abalone (Haliotis fulgens Philippi) In the Southern part of its range, Baja California Sur, Mexico, assessed by an analysis of gut contents. Journal of Shellfish Research 17 (3): 777–782. [ Links ]
Siqueiros–Beltrones, D.A. 2002. Diatomeas bentónicas de la Península de Baja California; diversidad y potencial ecológico. Secretaría de Educación Pública–Universidad Autónoma de Baja California Sur–Centro Interdisciplinario de Ciencias Marinas del Instituto Politécnico Nacional. México. 102 p. [ Links ]
Siqueiros–Beltrones, D.A. 2005. Una paradoja sobre uniformidad vs. orden y estabilidad en la medida de la diversidad de especies según la teoría de la información. Ludus Vitalis 13 (24): 1–10. [ Links ]
Siqueiros–Beltrones D.A. & D. Voltolina. 2000. Grazing selectivity of red abalone Haliotis rufescens poslarvae on benthic diatom films under culture conditions. Journal of the World Aquaculture Society 31 (2): 239–246. [ Links ]
Siqueiros–Beltrones, D.A. & G. Valenzuela–Romero. 2004. Benthic diatoms assemblages in an abalone (Haliotis spp.) habitat in the Baja California peninsula. Pacific Science 58 (3): 435–446. [ Links ]
Siqueiros–Beltrones, D.A., G. Valenzuela–Romero, O.U. Hernández–Almeida, U. Argumedo–Hernández & F.O. López–Fuerte. 2004. Catálogo iconográfico de diatomeas de hábitats rocosos y su incidencia en la dieta de abulones (Haliotis spp.) jóvenes de Baja California Sur, México. CICIMAR–Oceánides 19 (2): 1–79. [ Links ]
Siqueiros–Beltrones, D.A., S. Guzmán del Próo & E. Serviere–Zaragoza. 2005. Main diatom taxa in the natural diet of juvenile Haliotis fulgens and H. corrugata (Mollusca: Gastropoda) in Bahía Tortugas and Bahía Asunción, B.C.S., México. Pacific Science 59 (4): 581–592. [ Links ]
Tutschulte, T.C. & J.H. Connell. 1988. Feeding behavior and algal food of three species of abalone (Haliotis) in southern California. Marine Ecology Progress Series 49: 57–64. [ Links ]
Witkowski, A., H. Lange–Bertalot & D. Metzeltin. 2000. Diatom flora of Marine coast I. A. R. G. Gantner–Verlag. Konigstein, Germany. 925 p. [ Links ]

