










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.17 no.3 Ciudad de México dic. 2007
Artículos
Pastoreo del mesozooplancton sobre el fitoplancton en la Bahía de La Paz, B. C. S., México
Mesozooplankton grazing on phytoplankton in La Paz Bay B. C. S., Mexico
Sergio Hernández-Trujillo1,2, Alejandro Zárate-Villafranco1, Rocío Pacheco-Chávez1, Gabriela Esqueda-Escárcega1, José Reyes Hernández-Alfonso1 y Gerardo Aceves-Medina1,2
1 Becario COFAA. CICIMAR-IPN. Departamento de Plancton y Ecología Marina. A. P. 592, 23000 La Paz, Baja California Sur. México.
2 Becario EDI. CICIMAR-IPN. Departamento de Plancton y Ecología Marina. A. P. 592, 23000 La Paz, Baja California Sur. México.E-mail: strujil@ipn.mx
Recibido: 23 de junio de 2006
Aceptado: 25 de octubre de 2007
Resumen
Durante octubre de 2002 se generó una serie de tiempo para medir la tasa de alimentación diaria del mesozooplancton, estudiando los cambios de la concentración de clorofila a en una estación en la Bahía de La Paz, México. El coeficiente de consumo o tasa de pastoreo (g), el coeficiente de crecimiento de fitoplancton (k), la tasa de aclaramiento del agua (F) y la tasa de ingesta (I) fueron estimados como una referencia inicial para el área de estudio. La tasa de pastoreo tuvo valores entre 0.72-1 y 2.74 ind·d-1; mientras que el coeficiente de crecimiento del fitoplancton fue de 0.03 a 0.76 d-1. La tasa de aclaramiento de agua se determinó entre 11.9 y 45.6 ml·ind-1·d-1 y la tasa de ingesta entre 5.2 y 18.4 Cla·ind-1·d-1. El mesozooplancton pudo remover del 1.0 al 43 % de la producción potencial diaria. Las tasas de aclaramiento y de ingesta tuvieron una variación diaria similar a lo largo del estudio; el coeficiente de crecimiento día a día, fue menor al de pastoreo a lo largo del estudio. Los resultados mostraron que el mesozooplancton puede consumir alrededor del 17% de la producción diaria de fitoplancton en la zona estudiada. Esta primera aproximación sobre la alimentación del mesozooplancton en la bahía de La Paz, permite una mayor comprensión de la dinámica planctónica en esta área subtropical.
Palabras clave: Tasa de pastoreo, tasa de aclaramiento, tasa de ingesta, zooplancton.
Abstract
During October 2002 a time series was generated to measure the daily grazing rates of mesozooplankton, it was recorded the changes of the chlorophyll a concentration at a station in La Paz Bay, Mexico. The grazing coefficient (g), phytoplankton growth coefficient (k), clearance rate (F), and ingestion rate (I) were estimated as initial reference for the study area. Grazing rates presented values ranging from 0.72-1 to 2.74 ind·d-1; phytoplankton growth coefficient ranked from 0.03 to 0.76 d-1 Clearance rate was from 11.9 to 45.6 ml·ind-1·d-1, and the ingestion rate fluctuated from 5.2 to 18.4 Cla·ind-1·d-1. The mesozooplankton was capable of remove 1.0 to 43 % of potential daily production. The ingestion and clearance rates had similar daily variation pattern along the study; phytoplankton growth coefficient values were lower than grazing rate. The results showed that mesozooplankton might consume about 17% of phytoplankton production in this coastal area. This assessment on mesozooplankton feeding at La Paz Bay is a step to a better understanding of the plankton dynamic in this subtropical area.
Keywords: Grazing rate, clearance rate, ingestion rate, zooplankton.
Introducción
Una de las vías por las que la materia y la energía pasan de un nivel trófico inferior a uno superior es la alimentación, por lo que su cuantificación es un factor clave en el estudio de las interacciones tróficas. En el caso del zooplancton marino, este tipo de análisis requiere de métodos distintos (Båmstedt et al., 2000) y se han propuesto varias formas de abordarlo. El consumo de fitoplancton por el zooplancton es conocido como pastoreo o herbivoría y es considerado como la vía principal de transferencia de energía a otros niveles tróficos (Conover & Huntley, 1980). Por tal razón al zooplancton herbívoro se le reconoce un importante papel en los ecosistemas marinos. Los más útiles indicadores del impacto trófico del zooplancton en el fitoplancton, en estudios de alimentación, son la tasa de filtración o aclaramiento del agua, la tasa de ingesta o ración diaria y el coeficiente de consumo o tasa de pastoreo (García-Pámanes et al., 1991, Båmstedt et al., 2000).
La ecología trófica tiene variaciones en sus indicadores que pueden ser originadas por el tamaño del organismo, el sexo, el estadio de vida, la concentración y tipo de alimento o la composición química del alimento, entre otros factores, éstos influyen de modo diferencial en los distintos hábitas (García-Pámanes et al., 1991), por lo que la estimación para un área particular debe hacerse in situ (Wang & Conover, 1986), ya que es problemático extrapolar información de un ambiente a otro, especialmente del templado al tropical (Turner et al., 1998). Los estudios de ecología alimenticia del zooplancton marino se han efectuado principalmente en hábitats templados y sólo algunos en ambientes tropicales. Actualmente se reconoce que el zooplancton es una importante fuente de mortalidad del fitoplancton en los ambientes marinos, lacustres y estuarinos (Landry & Hassett 1982; Jeppesen et al., 1996; Gallegos, 1989; Calbet & Landry, 2004). En aguas costeras mexicanas templadas García-Pámanes (1989), García-Pámanes et al. (1991) y Lara-Lara y Matus-Hernández (1997), han sido los que han estimado el proceso de pastoreo, por lo que este estudio tuvo como objetivo principal obtener las tasas de pastoreo, de filtración, de ingesta y de crecimiento del fitoplancton a través de una serie de tiempo en la Bahía de La Paz, B. C. S., México.
Area de estudio
La Bahía de La Paz se localiza en la porción sur-occidental del Golfo de California, la temperatura mínima superficial del mar es de 20°C en invierno-primavera y la máxima 31°C en verano (Espinoza & Rodríguez, 1987), a diferencia de la temperatura ambiente que registra un promedio de 8 °C en invierno y 37°C en verano; el clima es seco, con una precipitación menor a 200 mm anuales con máximos en septiembre (Cruz-Ayala, 1996). Los vientos predominantes de invierno son del noroeste (2-3 m-1·s-1), con rachas de hasta 10 m-1·s-1, mientras que en verano los vientos tienen una componente sur con intensidades medias de 2-3 m-1·s-1 (Robles-Gil-Mestre, 1998). El régimen de mareas es mixto semidiurno, con las mareas más bajas entre febrero y abril, y las más altas entre julio y septiembre (Obeso-Nieblas et al., 1993). En la zona sur de la bahía, y en estaciones equivalentes a la que aquí se ocupa, la clorofila a presenta un patrón estacional inverso a la temperatura y a la transparencia del agua. Las menores concentraciones de clorofila a integrada ocurren en los meses más cálidos (<10 mg·m-2) y los máximos se presentan durante la época fría (142.8 mg·m-2) y parecen estar asociados con fuertes procesos de eutrofización en verano y procesos de mezcla de la columna de agua en invierno (Martínez-López, et al., 2001).
Material y metodos
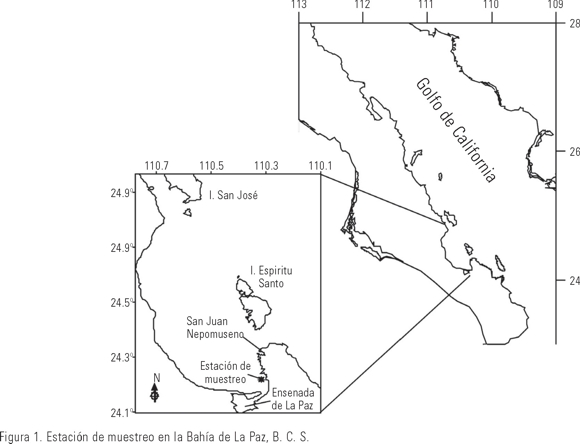
Del 16 al 24 de octubre de 2002, diariamente se llevaron a cabo muestreos en una estación oceanográfica localizada frente a Punta Prieta (Fig. 1) en la Bahía de La Paz, Baja California Sur (24°14' N y 110° 19' O). Se tomaron muestras de agua a tres profundidades (0, 5 y 10 m) con botellas Niskin de 5 L de capacidad, para determinar la concentración de clorofila a de acuerdo a Jeffrey y Humphrey (1975). Posteriormente se recolectó más agua en bidones de 20 L de capacidad, para filtrarla en el laboratorio (Filtros Whatman GF/C de 47 mm ø) y que sirvió para separar el fitoplancton del macrozooplancton, así como para preparar las réplicas de cada experimento efectuado cada día. La temperatura del mar se midió con un CTD Seabird, los datos de la velocidad del viento fueron obtenidos de la estación meteorológica ubicada en el CIBNOR. Las muestras de zooplancton fueron obtenidas mediante arrastres subsuperficiales con una red cilindro-cónica de 0.60 m de diámetro y una luz de malla de 300 µm. El material zooplanctónico obtenido diariamente fue transportado al laboratorio y con ayuda de microscopios estereoscópicos fueron extraídos los copépodos y se colocaron 60 ejemplares de Acartia lilljeborgii Giesbrecht, 1889 en cada una de las tres botellas de incubación que contenían el agua de mar superficial previamente filtrada, con fitoplancton, la cual sirvió de alimento a los copépodos. Las botellas de incubación se mantuvieron por 24 horas en completa oscuridad a la temperatura en que se obtuvieron en el arrastre. Se determinó la concentración de clorofila a al inicio y al final de cada experimento en cada una de las botellas de incubación, así como a sus respectivas réplicas. La tasa de pastoreo (CoefCons·t-1) se determinó de acuerdo a Båmstedt et al. (2000):
g = (lnClat0 - Clat0-1)/t
donde g es el coeficiente instantáneo de alimentación o pastoreo y Clat0 y Clat1 son las concentraciones de clorofila a al inicio y al final del experimento, respectivamente; t es el tiempo de incubación en horas.
La tasa de filtración se calculó de acuerdo a Marín et al. (1986):
F = V·g/N
donde, V es el volumen de la botella de incubación en ml, g es el coeficiente instantáneo de alimentación o pastoreo y N, es el número de consumidores por botella experimental. El cálculo de la tasa de ingesta (Cla·ind-1·t-1) fue determinado de acuerdo a Båmstedt et al. (2000), de la siguiente manera:
I = F · C
donde F es la tasa de filtración y C es el promedio de la concentración experimental de clorofila a. La tasa de crecimiento de fitoplancton se calculó de acuerdo a Landry y Hasset (1982):
k = (ln control) - (ln inicial)/t
donde el control es la concentración de clorofila a que hay en las botellas de incubación sin consumidores; el inicial es la concentración de clorofila a en las botellas experimentales que contienen consumidores; t es el tiempo en horas. El porcentaje de la producción potencial de clorofila a consumida por el zooplancton fue calculado mediante la relación entre la tasa de crecimiento de fitoplancton y la tasa de pastoreo:
P = (k/g)*100
Se llevó a cabo un análisis de varianza para detectar las diferencias en la tasa de pastoreo, la tasa de filtración, la tasa de ingesta y la tasa de crecimiento de fitoplancton (p = 0.05) con cada día de experimentación.
Resultados
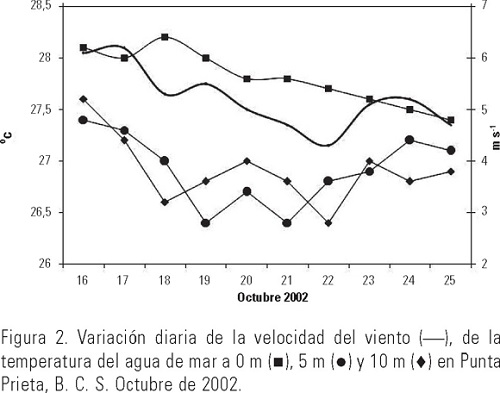
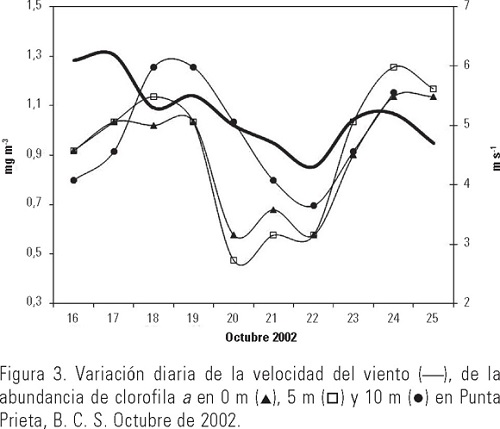
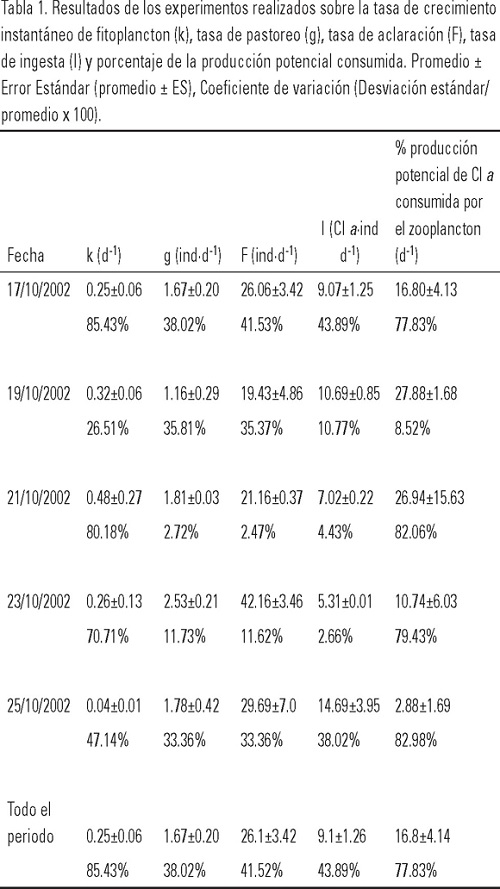
El intervalo de la temperatura del mar fue de 27.4 a 28.2 °C; observándose que a 0 m disminuyó hacia los últimos días del muestreo y que a 5 y 10 m esta reducción se presentó gradualmente; se registró una diferencia de hasta 2°C entre 0 y 5-10 m (Fig. 2). Esta información fue utilizada diariamente para mantener en los experimentos de laboratorio condiciones similares de temperatura. La velocidad del viento en el sitio de muestreo, fluctuó entre 4.3 y 6.2 m·s-1, mostrando una tendencia a disminuir a partir del primer día de experimentación. Respecto a la concentración de clorofila a, el intervalo obtenido fue de 0.47 a 1.3 mg·m-3. Puede observarse en los tres niveles la gran similitud en el patrón de variación diaria, los máximos se registraron del tercer al cuarto día a los 10 m y otro en el noveno día en los tres niveles medidos (Fig. 3). Las tasas de filtración de agua (F) fueron significativamente distintas entre días (p < 0.05) y estuvieron entre 11.9 y 45.6 ml-1ind-1d-1, mostrando una tendencia ascendente desde el segundo día de incubaciones, alcanzando un máximo en el cuarto, para después descender (Tabla 1). La tasa de ingesta (I) fue estimada desde 5.2 a 18.4 mg de clorofila consumida por ind-1d-1, registrándose dos máximos, uno al segundo y otro al quinto día de incubaciones; en este caso también hubo diferencias significativas entre días (p < 0.05). El coeficiente de crecimiento instantáneo de fitoplancton (k) fluctuó entre 0.03 y 0.76 d-l sin ser significativa la diferencia entre días (p > 0.05); el coeficiente de consumo de clorofila a (g) se estimó entre 0.72 y 2.74 d-l, observándose un sistemático y significativo aumento (p < 0.05) del pastoreo a lo largo de los días y una pequeña reducción en el quinto día de experimentación (Fig. 4). El porcentaje de la producción potencial de clorofila a consumido por el zooplancton osciló significativamente (p < 0.05) entre 1.0 y 43%, con una media de ≈17%.
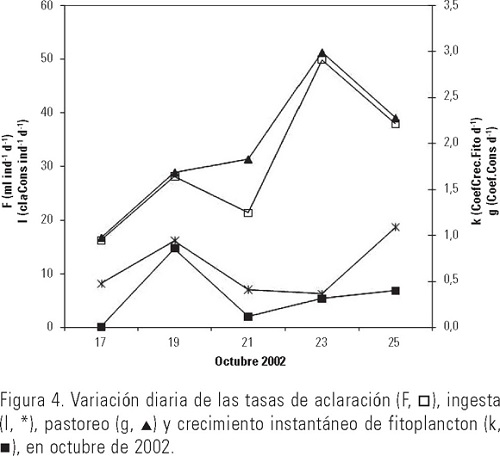
Discusión
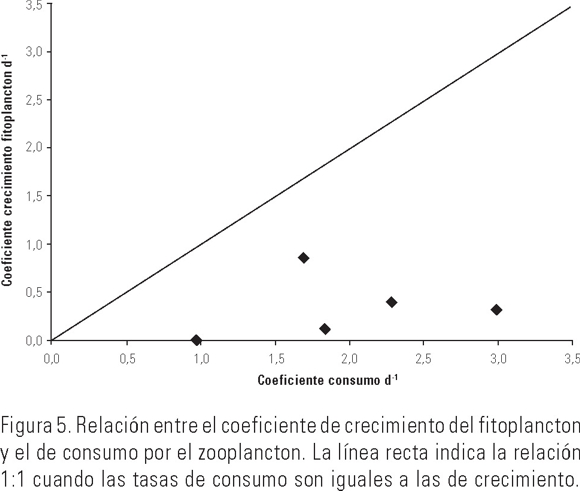
A lo largo del periodo de estudio la temperatura superficial del mar y la concentración de clorofila a tuvieron una variabilidad que pudo estar influenciada, en buena parte, por la turbulencia ocasionada por la velocidad del viento, ya que a partir de la disminución de la velocidad del mismo en los primeros siete días (6.2 a 4.3 m·s-1) coincidieron el enfriamiento del agua, la disminución de la concentración de clorofila a (Figs. 2 y 3) y la reducción del coeficiente de crecimiento de fitoplancton en 87%. En la última etapa de los experimentos, en el sitio de muestreo se registró un ligero aumento en la velocidad del viento, y por tanto de la turbulencia, lo que pudo ser la causa más probable de elevación de la concentración de clorofila a y del crecimiento de fitoplancton, aunque no lo suficiente para ser detectado por el análisis estadístico. En general, las tasas de filtración del zooplancton estimadas en nuestro estudio estuvieron en el intervalo encontrado por García-Pámanes (1989) y por Lara-Lara y Matus-Hernández (1997) para la Bahía de Todos Santos, Baja California (Tabla 2). También se observaron variaciones diarias significativas de más del 100% de las tasas de filtración y de pastoreo a lo largo del estudio, lo que coincide con los resultados de Lara-Lara y Matus-Hernández (1997), ya que ellos también reportaron cambios diarios de más de 100% en dichos indicadores tróficos. Los intensos eventos de mezcla que se registran en la Bahía de Todos Santos, serían uno de los factores para esa diferencia con la Bahía de La Paz, en la que estos procesos son de menor magnitud y localizados estacionalmente (Obeso-Nieblas et al., 1993). En el caso del mesozooplancton de la Bahía de La Paz, la magnitud de los indicadores tróficos fue en general, mayor y el porcentaje del consumo potencial promedio de fitoplancton fue 2% superior que lo reportado por Lara-Lara y Matus-Hernández (1997) para la Bahía de Todos Santos, lo que indica el importante impacto que ocasiona esta fracción del zooplancton en las poblaciones de fitoplancton en la Bahía de La Paz durante la temporada de otoño de 2002, lo que lo coloca como fuente importante de alimento para otros organismos zooplanctónicos, como los copépodos. De los resultados obtenidos, puede concluirse que el intervalo observado para la tasa de filtración se encuentra comprendido entre las estimaciones obtenidas con varios métodos y distintos tipo de alimento (Tabla 2). Sin embargo, en las primeras estimaciones que fueron efectuadas entre las década de 1970 y la de 1980 se utilizaron como alimento determinadas especies de fitoplancton que arrojaron volúmenes de filtración por copépodo de más de 1000 ml·ind-1·d-1, lo que contrasta con los estimados en trabajos posteriores durante la década de 1990 cuyos métodos son el 14C y clorofila a y que emplean alimento natural en los experimentos. De hecho, la comparación más adecuada es la que se puede establecer con aquellos resultados obtenidos con el método de clorofila a empleando alimento natural. Así, puede observarse que las estimaciones de Lara-Lara y Matus-Hernández (1997) para una zona templada del Pacífico mexicano, fueron alrededor de 80% más bajas a las obtenidas en este trabajo. En cambio, para la zona tropical en la que Turner et al. (1998) estimaron la tasa de filtración, se observa que lo obtenido en la zona de estudio fue menor en alrededor de 20%, en términos globales. En todos los casos la gran variabilidad diaria que se presentó indica la necesidad de contar con observaciones continuas sobre las mismas especies para poder entender la dinámica de esta fracción del zooplancton en el ecosistema. La relación entre el crecimiento del fitoplancton y el consumo del mismo por parte de los copépodos en la Bahía de La Paz, permitió observar que estos últimos consumieron en mayor cantidad el fitoplancton, impidiendo que se acumulara la clorofila (Fig. 5); la relación productores/consumidores en esta época del año se encontró desbalanceada hacia los copépodos, lo que apoya los resultados obtenidos acerca de la ingesta de clorofila a y el subsiguiente consumo potencial de ésta por parte del mesozooplancton. Esta primera aproximación al pastoreo en las poblaciones de fitoplancton en la Bahía de La Paz, permite establecer una referencia inicial sobre la dinámica productor-consumidor en este ambiente pelágico costero, así como el nivel de variación que se presenta día a día no sólo de estos indicadores tróficos, sino también de algunas de las variables que se relacionan directamente con la disponibilidad de fitoplancton para los copépodos pelágicos. Es necesario realizar más estimaciones de pastoreo en la Bahía de La Paz con un diseño experimental que incluya un mayor número de observaciones y réplicas, así como a nivel de especies dominantes y por época del año para poder establecer el papel que tiene el zooplancton en la ecología trófica en la zona de estudio.
Agradecimientos
A los alumnos de posgrado Gladis López Ibarra, Arturo Nava Torales y Javier Cruz Hernández quienes participaron en los muestreos, trabajo de laboratorio y separación de los ejemplares vivos para el procedimiento de experimentación. Al PIFI-IPN por el apoyo económico a los estudiantes de posgrado. Los fondos para llevar a cabo este trabajo provinieron de la Coordinación General de Posgrado e Investigación del IPN a través del apoyo asignado al proyecto 20020337.
Referencias
ANRAKU, M. 1964. Influence of the Cape Cod Canal on the hydrography and on the copepods of Buzzards Bay and Cape Cod Bay, Massachusetts. II. Respiration and feeding. Limnology and Oceanography 9: 195-206. [ Links ]
BÅMSTEDT, U., D. J. GIFFORD, X. IRIGOIEN, A. ATKINSON & M. ROMAN. 2000. Feeding. In: R. P. Harris, P. H. Wiebe, J. Lenz, H. R. Skjoldal & M. Huntley (Eds) ICES Zooplankton Methodology Manual. Academic Press, Londres, pp. 297-399. [ Links ]
CALBET, A. & M. L. LANDRY. 2004. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnology and Oceanography 49(1): 51-57. [ Links ]
CONOVER, R. J. 1956. Oceanography of Long Island Sound 1952-1954. VI. Biology of Acartia clausi and A. tonsa. Bulletin of Bingham Oceanography Collection 15: 156-233. [ Links ]
CONOVER, R. J. & M. E. HUNTLEY. 1980. General rules of grazing in pelagic ecosystems. In: P.G. Falkowsky (Ed.). Primary Productivity. Plenum Press, New York, pp. 461-484. [ Links ]
CRUZ-AYALA, M. B. 1996. Variación espacio-temporal de la ficoflora y su abundancia relativa en la Bahía de La Paz, B. C .S. México. Tesis de Maestría. CICIMAR, IPN, La Paz, B. C. S. 90 p. [ Links ]
ESPINOZA, J. & H. RODRÍGUEZ. 1987. Seasonal phenology and reciprocal transplantation of Sargassum sinicola Setchell et Gardner in the southern Gulf of California. Journal of Experimental Marine Biology Ecology 110: 183-195. [ Links ]
FIEDLER, P. C. 1982. Zooplankton avoidance and reduced grazing responses to Gymnodinium splendens (Dinophyceae). Limnolology and Oceanography 27: 961-965. [ Links ]
FROST, B. W. 1972. Effect of size and concentrations of food particles on the feeding behavior of the marine planktonic copepod Calanus pacificus. Limnology and Oceanography. 17: 805-815. [ Links ]
GALLEGOS, CH. 1989. Microzooplankton grazing on phytoplankton in the Rhode River, Maryland; nonlinear feeding kinetics. Marine Ecology Progress Series 57: 23-33. [ Links ]
GARCÍA-PÁMANES, J. 1989. Variación día a día de la tasa de pastoreo zooplanctónico frente a Baja California México. Tesis de Maestría CICESE, Ensenada, B. C. 76 p. [ Links ]
GARCÍA-PÁMANES, J. R. LARA-LARA & G. GAXIOLA-CASTRO. 1991. Daily zooplankton filtration rates off Baja California. Estuarine, Coastal and Shelf Science 32: 503-510. [ Links ]
HUNTLEY, M. E. 1982. Yellow water in La Jolla Bay, California, July 1980. 2. Suppression of zooplankton grazing. Journal Experimental Marine Biology Ecology 63: 81-91. [ Links ]
JEFFREY, S. W. & G. F. HUMPHREY. 1975. New spectrophotometric equations for determining chlorophylls a, b, c1, and c2 in higher plants, algae and natural phytoplankton. Biochemical Physiology Pflanz 167: 191-194. [ Links ]
JEPPESEN, E., M. SØNDERGAARD, J. P. JENSEN, E. MORTENSEN & O. SORTJÆR. 1996. Fish-induced changes in zooplankton grazing on phytoplankton and bacterioplankton: a long-term study in shallow hypertrophic Lake Søbigaard Journal of Plankton Research 18(9): 1605-1625. [ Links ]
LANDRY, M. R. & R. P. HASSET 1982. Estimating the grazing impact of marine micro-zooplankton. Marine Biology 67: 283-288. [ Links ]
LARA-LARA, J. R. & E. MATUS-HERNÁNDEZ. 1997. Tasas diarias de pastoreo del macrozooplancton en la costa de Baja California. Ciencias Marinas 23(1): 71-81. [ Links ]
MARÍN, V., M. E. HUNTLEY & B. W. FROST. 1986. Measuring feeding rates of pelagic herbivores: analysis of experimental design and methods. Marine Biology 93: 49-58. [ Links ]
MARTÍNEZ-LÓPEZ, A, R. CERVANTES-DUARTE, A. REYES-SALINAS & J. E. VALDEZ-HOLGUÍN. 2001. Cambio estacional de clorofila a en la Bahía de La Paz, B.C.S., México. Hidrobiológica 11(1): 45-52. [ Links ]
OBESO-NIEBLAS, M., A. R. JIMÉNEZ-ILLESCAS & S. TROYO-DIÉGUEZ. 1993. Modelación de la marea en la Bahía de La Paz, B. C. S. Investigaciones Marinas CICIMAR 18 (1): 13-22. [ Links ]
PAFFENHOFFER, G. A. 1971. Grazing and ingestion rates of nauplii, copepodids and adults of planktonic copepod Calanus helgolandicus. Marine Biology 11: 286-298. [ Links ]
ROBLES-GIL-MESTRE, S. 1998. El clima de la ciudad de La Paz. Tesis de Maestría. UNAM. México, D. F. 233 p. [ Links ]
TURNER, J. T., R. R. HOPCROFT, J. A. LINCOLN, C. S. HUESTIS, P. A. TESTER & J. C. ROFF. 1998. Zooplankton feeding ecology: grazing by marine copepods and cladocerans upon phytoplankton and cianobacteria from Kingston Harbour, Jamaica. Marine Ecology 19 (3): 195-208. [ Links ]
WANG, R. & R. J. CONOVER. 1986. Dynamics of gut pigment in the copepod Temora longicornis and the determination of in situ grazing rates. Limnology and Oceanography 31: 867-877. [ Links ]

