










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.17 no.3 Ciudad de México dic. 2007
Artículos
Variación espacial de la comunidad de medusas (Cnidaria) del sur del Golfo de México, durante el otoño de 1999
Spatial variation of medusan community (Cnidaria) in the Southern Gulf of Mexico, during autumn 1999
Lucio Loman-Ramos1, Uriel Ordóñez-López1 y Lourdes Segura-Puertas2
1 Instituto Politécnico Nacional, Centro de Investigación y Estudios Avanzados, Unidad Mérida. Km 6, carretera antigua a Progreso, A.P. 73, Mérida, Yucatán, México. 97310. lucioloman@prodigy.net.mx
2 Universidad Nacional Autónoma de México, Instituto de Ciencias del Mar y Limnología, Unidad Académica Puerto Morelos. Apdo. Postal 1152, Cancún, Quintana Roo, México. 77501.
Recibido: 8 de noviembre de 2006
Aceptado: 3 de octubre de 2007
Resumen
Se analizó la variación espacial de la comunidad de medusas durante el otoño de 1999 en la región sur del Golfo de México. El material planctónico fue recolectado en 69 estaciones mediante arrastres superficiales utilizando una red tipo Bongo. Se determinaron 38 especies de medusas (35 hidromedusas, dos escifomedusas y una cubomedusa). Liriope tetraphylla fue la especie dominante al representar más del 78% de la fauna recolectada. Otras especies importantes por su abundancia fueron: Cunina octonaria, Clytia folleata, Aglaura hemistoma, Solmundella bitentaculata, Pegantha triloba y Octophialucium medium. Seis de las especies determinadas representaron nuevos registros geográficos para la región sur del Golfo de México. La distribución de las especies más abundantes indicó una concentración de organismos en las aguas someras del Banco de Campeche sin embargo, algunas como Nausithoe punctata y Euphysora gracilis, mostraron sus mayores abundancias sobre la estrecha plataforma continental de los estados de Veracruz y Tabasco. La especie dominante actuó como estabilizadora de la estructura de la comunidad aunque las especies de menor abundancia denotaron mejor los cambios de la comunidad en las distintas zonas oceanográficas. En este sentido, la variación espacial de los componentes estructurales de la comunidad durante el otoño, estuvo relacionada con la circulación oceánica imperante sobre la plataforma continental y la disponibilidad de alimento.
Palabras clave: Medusas, distribución, composición, sur del Golfo de México.
Abstract
The spatial variation of the jellyfish community during fall of 1999 in the southern region of the Gulf of Mexico was analyzed. Plankton material was collected through superficial Bongo-type net tows at 69 sites. Thirty-eight species of jellyfish (35 hydromedusae, two scyphomedusae and one cubomedusa) were found. Liriope tetraphylla was the dominant species, representing 78% of the collected fauna. Other important species due to their abundance were Cunina octonaria, Clytia folleata, Aglaura hemistoma, Solmundella bitentaculata, Pegantha triloba and Octophialucium medium. Six of the identified species are new records for the southern region of the Gulf of Mexico. The distribution of the most abundant species indicated a great concentration of organisms in the shallow waters of Campeche Bank. However, others like Nausithoe punctata and Euphysora gracilis showed their greatest abundances across the stretch continental shelf of the States of Veracruz and Tabasco. The dominant species acted as a community structure stabilizer, but less abundant species better denoted abundance changes in the community in particular oceanographic zones. In this sense, the spatial variation of the community structural components in the autumn season was related to the main oceanographic currents in the continental shelf and food availability.
Key words: Medusae, distribution, composition, southern Gulf of Mexico.
Introducción
Las medusas, al igual que el resto de los cnidarios, conforman un grupo exitoso debido a que son abundantes en una gama amplia de ambientes acuáticos, habitando en todas las latitudes; conjuntamente, las medusas son depredadores planctófagos que operan en los niveles tróficos superiores de los ecosistemas marinos en un amplio intervalo de productividad (Mills, 1995). Dada su extraordinaria capacidad depredadora, las medusas pueden competir por el alimento con otros carnívoros y devorar a organismos del zooplancton como copépodos, quetognatos, larváceos, cladóceros, huevos, larvas de peces (Alvariño, 1985), así como una gran variedad de organismos del meroplancton y aún a otras medusas (Purcell, 2003). Aunque las medusas habitan grandes áreas oceánicas y costeras, la mayoría de las especies tiene patrones bien definidos de distribución y abundancia, muchos de los cuales muestran una marcada similitud con las masas de agua (Vannucci, 1957); de esta manera, algunas especies han sido consideradas como indicadores biológicos de condiciones oceanográficas (Ramírez, 1977).
Las medusas ocurren con regular frecuencia en diversas regiones del Golfo de México, como lo han manifestado Phillips (1972) y Correia (1992), observándose en muchas ocasiones, incrementos masivos de sus poblaciones (Alvariño, 1985). Las aguas productivas en el sur del Golfo de México, propician una alta riqueza de la fauna planctónica, conformada por alrededor de 15 grandes grupos de zoopláncteres, incluidas las hidromedusas (De la Cruz, 1972). Los estudios de medusas en las regiones tropicales de los oceános Pacífico y Atlántico se han centrado en aquellos procesos físicos dados por las características oceanográficas y la variabilidad espacial y temporal (Ramírez & Zamponi, 1981; Segura-Puertas, 1984;1992; 2003; Zamponi et al. 1990; Segura-Puertas & Ordoñez-López, 1994; Ramos & Segura-Puertas, 2004). Los trabajos publicados sobre las medusas en el Golfo de México y áreas adyacentes son escasos pero detallados en cuanto a su composición, abundancia y distribución, entre ellos destacan los de Kramp (1959), Phillips (1972), Gómez-Aguirre (1986), Zamponi y Suárez (1991), Correia (1992), Segura-Puertas (1992), Segura-Puertas y Ordóñez-López (1994), Suárez-Morales et al. (1999; 2002), Segura-Puertas (2003), Segura-Puertas et al. (2003) y Ramos y Segura-Puertas (2004).
De lo anterior, se desprende la necesidad de contar con un marco robusto sobre la dinámica oceanográfica en el sur del Golfo de México como parte fundamental e importante para explicar los cambios en la estructura del zooplancton gelatinoso. Así, el escaso conocimiento en la dinámica espacial de las medusas y sus relaciones con las masas de agua y otros organismos ecológicamente importantes como son los copépodos, los quetognatos y las larvas de peces entre otros, dada la abundancia de los primeros así como por el eslabon que conforman entre diversos niveles tróficos, justifican este estudio, que tiene como propósito fundamental determinar la variación espacial de los componentes estructurales de la comunidad de medusas en el sur del Golfo de México durante el otoño de 1999, región influenciada por las características oceanográficas locales (temperatura, salinidad, densidad, corrientes y giros ciclónicos), mientras que su abundancia podría estar en relación a la disponibilidad del alimento potencial (biomasa zooplanctónica).
Materiales y métodos
El área de estudio se localiza en el sur del Golfo de México y está comprendida entre los 18° 28' 52" y los 20° 50' 39" N y los 91° 15' 43" y 94° 49' 59" W. Esta zona se encuentra delimitada por un polígono que cubre el área costera desde Punta Zapotitlán en el Estado de Veracruz hasta las costas de Yucatán frente a la Laguna de Celestún (Fig. 1). A nivel oceánico, el sur del golfo presenta una capa de mezcla superficial que comprende los primeros 100 ó 150 m de profundidad y la circulación oceánica está influenciada estacionalmente por el viento, la intrusión de la Corriente de Lazo y por las características físicas de las masas de agua. Particularmente, el último tercio del año se carateriza por la presencia de una circulación ciclónica que determina el patrón de circulación de las masas de agua sobre el Banco de Campeche (Monreal-Gómez & Salas de León, 1990).
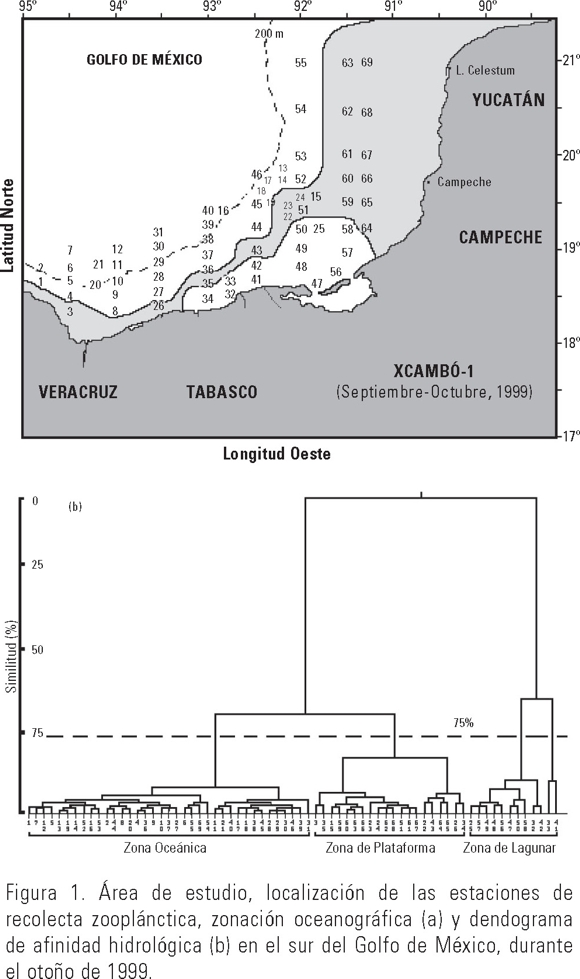
La recolección de las medusas se efectuó durante la campaña oceanográfica XCAMBÓ-1, del 29 septiembre al 12 octubre de 1999. Para tal efecto se realizó un derrotero de 69 estaciones a bordo del B/O "Justo Sierra" (Fig. 1). En cada sitio se efectuaron arrastres superficiales de 10 min con una red tipo Bongo y mallas filtrantes de 500 μm, en cada boca se colocaron flujómetros para la determinación del volumen de agua filtrada. La temperatura y salinidad fueron determinados con una sonda CTD. El material recolectado fue fijado en formalina al 10% en agua de mar y neutralizada con borato de sodio (Harris et al., 2000). La biomasa zooplanctónica se obtuvo mediante la técnica de peso húmedo, expresándose los valores en g/1000 m3. Las medusas fueron separadas, cuantificadas y determinadas a nivel de especie con la ayuda de literatura especializada de la región y zonas adyacentes (Mayer, 1910; Kramp, 1959, 1961; Segura-Puertas, 1984; Boltovskoy, 1999). A los datos hidrológicos se les aplicó un análisis de clasificación por aglomeración a través del índice de Bray-Curtis (IBC) con el fin de identificar grupos de estaciones y sometidas al análisis de varianza (ANOVA) de una vía (p < 0.05; Zar, 1988) para conocer si existían diferencias respecto a una misma variable. Posteriormente, con el objeto de evaluar y jerarquizar la importancia de las especies en la estructura de la comunidad se obtuvieron los descriptores como la riqueza específica (S), la diversidad (Shannon-Weiner H'), equidad (J) y dominancia a través del índice de valor de importancia (IVI) (McCune & Grace, 2002), para la estimación de estos descriptores se recurrió al paquete estadístico ANACOM (De la Cruz-Agüero, 1993).
Resultados
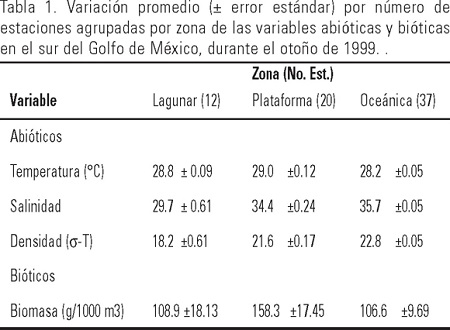
De manera general, el análisis de los principales parámetros hidrológicos denotó una marcada zonación hidrológica en el área estudiada. La temperatura superficial del agua varió de 27.6 a 29.9 °C, con un promedio para toda la campaña de 28.5 ± 0.5 °C. Los valores de temperatura más bajos se registraron hacia la zona oceánica y la plataforma continental de Veracruz y Tabasco, con un claro decremento frente a las desembocadura sde los Ríos Coatzacoalcos, Ver. y Dos Bocas, Tabasco. Los valores más altos (>29 °C) se localizaron por lo general, hacia las zonas someras del Banco de Campeche. La salinidad superficial osciló entre 23.7 y 36.4 con un valor medio de 35.3 ± 1.59. Los valores más altos se localizaron por lo general, hacia la zona oceánica y plataforma continental externa del área investigada, con un descenso hacia la desembocadura del Río Coatzacoalcos, Veracruz y de los sistemas fluviales Grijalva-Usumacinta y San Pedro-San Pablo en Tabasco, así como frente a la Laguna de Términos, Campeche.
La productividad planctónica indicó una fluctuación de la biomasa de 21.3 a 343.6 g/1000 m3, con una media de 119.3 ± 8.5 g/1000 m3. La biomasa mostró una distribución heterogénea, en donde las concentraciones mayores se localizaron sobre la plataforma continental externa de Veracruz, al noroeste de la Laguna de Términos y al oeste de Celestún (Fig. 2).
El análisis de clasificación con base en los datos hidrológicos permitió identificar tres grupos de estaciones de acuerdo al dendograma de afinidad (Fig. 1b) denominadas como: 1) zona de influencia lagunar (12 estaciones), 2) zona de plataforma (20 estaciones) y 3) zona oceánica (37 estaciones; Fig. 1a). La primera se caracterizó por presentar valores bajos de temperatura y salinidad; la segunda, por elevados valores de temperatura y salinidad coincidiendo con valores altos de biomasa zooplanctónica mientras que la tercera, se caracterizó por presentar salinidades y temperaturas intermedias con los valores de productividad planctónica también intermedios. Con la aplicación de un ANOVA de una vía (p < 0.05), se obtuvieron diferencias significativas entre las tres zonas hidrológicas (Tabla 1).
Medusas. Se recolectaron un total de 11,572 ejemplares de medusas que equivalen a una densidad promedio de 8,667 org./100 m3. Las densidades fluctuaron de 1 a 2,601 individuos entre estaciones con una media total de 125 ± 51 org./100 m3. Las densidades más altas se registraron en las aguas someras del Banco de Campeche, particularmente al noroeste del puerto de Campeche, mientras que las aguas sobre la plataforma continental de Veracruz y Tabasco mantuvieron bajas concentraciones (Fig. 2).
La fauna de medusas estuvo representada por 37 especies más una operacional (Obelia spp.), pertenecientes a 23 Familias y siete órdenes. La lista de especies con su autoridad nomenclatural se muestra en la Tabla 2. Las hidromedusas fueron el grupo más abundante representado por el 98.36%, las escifomedusas aparecieron con el 1.53% y las cubomedusas con el 0.11% del total de la fauna recolectada. Del total de especies determinadas, el 66% son formas neríticas y el 34% oceánicas. La mayoría de las especies ocurrieron en el estrato epipelágico, a excepción de Aglantha elata que fue considerada como batipelágica.
El Orden Conica incluyó el mayor número de Familias (siete). Otros órdenes importantes fueron Filifera (cinco) y Proboscoida (cinco). La Familia Geryoniidae fue la que aportó el mayor número de individuos a la comunidad con un 47.7%. Le siguieron, en orden decreciente, la Familia Campanulariidae con 9.2% y la Familia Cuninidae con 7.7% de los organismos recolectados. Estas tres Familias comprendieron en conjunto el 64.6% de los individuos capturados durante esta campaña oceanográfica. La Familia Campanulariidae fue la que presentó la mayor riqueza con seis especies, seguida de las Familias Bougainvillidae y Corymorphidae, ambas con tres especies cada una. Las Familias Pandeidae, Eirenidae, Cuninidae, Solmarisidae, Geryoniidae, y Rhopalonematidae estuvieron representadas sólo por dos especies cada una. Liriope tetraphylla fue la especie más abundante con 8,872 individuos (78.6%), seguida de Cunina octonaria con 819 (6.8%), Clytia folleata con 671 (4.7%), Aglaura hemistoma con 306 (2.6 %), Solmundella bitentaculata con 202 (1.6%), Nausithoe punctata con 160 (1.2%), Octophialucium medium con 106 individuos (1.0%) y Pegantha triloba con 122 (0.9%) y, que en conjunto representaron el 97.5 % de la abundancia total de los organismos recolectados (Tabla 2).
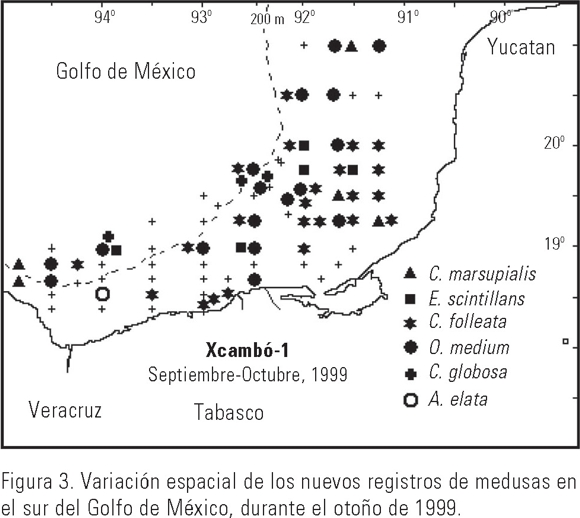
Las medusas estuvieron presentes en prácticamente toda el área de estudio. Las mayores densidades se registraron, por lo general, en las aguas de la plataforma continental sobre el Banco de Campeche, decreciendo en abundancia hacia la zona de influencia lagunar y oceánica. Liriope tretraphylla fue la especie dominante al representar el 47.7 % del IVI (Tabla 2). Las elevadas concentraciones de esta especie se observaron en las aguas someras (< 50 m de profundidad) del Banco de Campeche, decreciendo en número hacia aguas más profundas (> 50 m) (Fig. 2). Otras especies con un patrón de distribución similar fueron: Amphinema dinema, Merga violacea, Proboscidactyla ornata, Aequorea macrodactyla, Eutima gracilis, Eutonina scintillans, Laodicea undulata, Eucheilota duodecimalis, Octophialucium medium, Orchistoma pileus, Clytia discoida, C. folleata, S. bitentaculata, Cunina globosa, C. octonaria, Pengantha triloba, Aglaura hemistoma, Clytia hemisphaerica, C. simplex, Obelia spp. y Carybdea marsupialis. Por el contrario, especies como Bougainvillia muscus, Bougainvillia carolinensis, Cytaeis tetrastyla, Halitiara formosa, Vannuccia forbesii, Euphysora gracilis, Pegantha martagon, Aglantha elata, Nausithoe punctata y Pelagia noctiluca (Fig. 2), mostraron sus mayores abundancias sobre la estrecha plataforma continental de los estados de Veracruz y Tabasco. Hay que resaltar la presencia de importantes concentraciones de Lizzia alvarinoae, Corymorpha nutans y Geryonia proboscidalis en las aguas de la plataforma media y externa del Banco de Campeche. Dipurena ophiogaster se registró por lo general en las aguas costeras y Zanclea costata, en las aguas más profundas en el sur del Golfo de México. El único ejemplar recolectado de A. elata, considerada del medio batipelágico, se presentó cerca del talud continental, frente a la desembocadura del Río Coatzacoalcos, Veracruz. La mayoría de las especies identificadas en este estudio, han sido registradas previamente en aguas tropicales e incluso templadas, considerándolas por lo general como especies circumtropicales. Sin embargo, seis de ellas, Eutonina scintillans, O. medium, C. folleata, C. globosa, A. elata y C. marsupialis, constituyen nuevos registros para el área de estudio (Fig. 3).
La contribución de las especies varió de 0.1 a 2.5 con una diversidad total para el periodo de estudio de 1.42. La mayor diversidad se localizó por lo general, sobre el talud y la plataforma continental media del Banco de Campeche, y la menor, hacia las aguas costeras de Tabasco y Veracruz, observándose una importante concentración de especies al oeste de la desembocadura del sistema fluvial Grijalva-Usumacinta. El grado de homogeneidad existente en las abundancias relativas de las especies registradas, mostró una oscilación de 0.1 a 0.9, con un valor total de 0.2. Los valores más altos de equidad se localizaron sobre la plataforma de Veracruz y en general sobre el área oceánica, con algunos valores importantes en el Banco de Campeche.
En general, el IVI mostró que la especie dominante fue Liriope tetraphylla, al comprender más del 47.7%. Otras especies importantes por su abundancia y frecuencia fueron: C. octonaria, C. folleata, N. punctata, A. hemistoma, O. medium, S. bitentaculata, C. hemisphaerica, E. gracilis, P. triloba, B. muscus, A. dinema y Z. costata, que en conjunto con la especie dominante, sumaron el 87%; las 25 especies restantes representaron sólo el 13%. L. tetraphylla fue invariablemente la especie dominante en las tres zonas investigadas: influencia Lagunar con 50.7%, Plataforma con 49.9% y Oceánica con 39.3%. También se observó una contribución considerable de C. octonaria en la zona de influencia Lagunar (11.93%) y de Plataforma (9.09%), al igual que C. folleata para esta última zona (8.49%) y de N. punctata (14.55%) en la zona Oceánica (Tabla 2).
Discusión
Durante el otoño de 1999, las aguas oceánicas en el sur del Golfo de México, se caracterizaron por presentar densidades y salinidades mayores de 22.3 y 35.0 respectivamente, valores que coincidieron con los registrados por De la Lanza-Espino (1991) para el mismo periodo; estos valores de salinidad se encuentran dentro del intervalo de 34.0 a 36.5 considerado normal en el océano tropical abierto (De la Lanza-Espino, 2001). En contraste, las aguas cálidas y de salinidades elevadas registradas en el Banco de Campeche, sugieren un proceso importante de evaporación en esta región; así, las aguas someras del banco en su lento avance hacia el noreste durante esta época, se calientan más rápidamente, generando una mayor evaporación y concentración de sales, trayendo como consecuencia una mayor densidad del agua como lo han sugerido Capurro y Reid (1972), y sólo se registraron importantes descensos en la salinidad frente a la desembocadura de los principales sistemas fluvio-lagunares (De la Lanza-Espino, 2001).
Los núcleos de mayor biomasa zooplanctónica se localizaron sobre el Banco de Campeche (304. 2 y 312.4 g/1000 m3), lo que sugiere una importante actividad reproductiva de los organismos, así como el desplazamiento de dichos cúmulos por efecto de las corrientes oceánicas en esta época y a la propia movilidad de los organismos. Al respecto Houde y Chitty (1976), han sugerido una productividad similar en la plataforma de Florida para esta misma época, en tanto que Harris et al. (2000), han mencionado que las estructuras de locomoción del plancton gelatinoso y otros zooplancteres son tan eficientes, que les permiten condicionar su propio movimiento dentro de las masas de agua en un intervalo de escalas acorde al tamaño de los individuos.
La conformación de zonas oceanográficas con diferentes características abióticas y bióticas en el sur del Golfo de México, ha sido observada de manera similar por Flores-Coto et al. (2000a; 2000b), quienes han señalado que la distribución de la abundancia del ictioplancton está asociada a los procesos físicos de mezcla, que son generados por los vientos y por el patrón general de circulación debido a la influencia de las descargas de las aguas continentales que actúan a mesoescala. Asimismo, la zonación presupone la conformación de distintas comunidades de zooplancteres; al respecto Orozco-Zúñiga (2005), ha registrado diferencias en la composición del zooplancton que integra cada zona; este autor también ha señalado que durante su estudio, el grupo de los copépodos fue el que sostuvo la mayor productividad secundaria en el sur del Golfo de México, con importantes contribuciones de quetognatos, sergéstidos y larvas de decápodos.
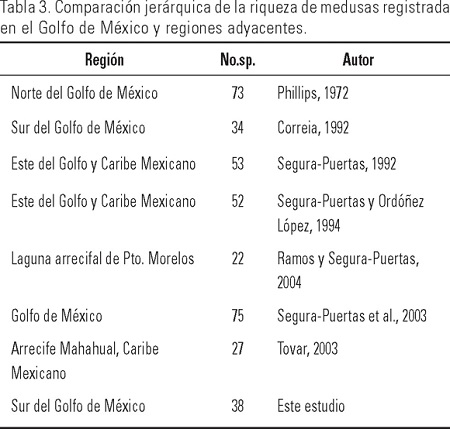
La riqueza de medusas registrada en el presente estudio (35 hidromedusas, dos escifomedusas y una cubomedusa), denota cierta similitud pero también diferencias si la comparamos con otros trabajos realizados en la región Atlántico occidental (Tabla 3). El número de especies fue similar al del trabajo de Correia (1992) para la misma zona (34 hidromedusas, una escifomedusa), sin embargo fue menor en la región norte del golfo y Caribe mexicano (48 hidromedusas, siete escifomedusas), en tanto que para la región norte del Golfo de México, Phillips (1972) registró 73 especies (56 hidromedusas, 16 escifomedusas y una cubomedusa). Estas diferencias pueden atribuirse en primera instancia, a una intensidad y periodicidad de muestreo diferentes, como se puede observar en los estudios de Phillips (1972), Segura-Puertas (1992; 2003) y Segura-Puertas y Ordóñez-López (1994), cuyos resultados derivaron de un mayor número de campañas oceanográficas cubriendo áreas (mayor número de hábitats) y periodos más extensos; o bien a una exhaustiva recopilación de trabajos con el objeto de integrar una lista de especies de medusas en el Golfo y Caribe mexicanos, como el trabajo realizado recientemente por Segura-Puertas et al. (2003). En este mismo sentido cabe descatar que el trabajo de Phillips (1972) efectuó un mayor número de recolectas en la zona costera y de influencia de las descargas de ríos en constraste con el medio oceánico, en tanto que en el estudio de Correia (1992) el esfuerzo de colecta se centro en el medio oceánico y el trabajo de Segura-Puertas y Ordóñez-López (1994) abarcó un mayor número de estaciones sobre la plataforma continental y zona de afloramiento de aguas profundas.
Los patrones de distribución de Liriope tetraphylla y de otras 21 especies de medusas, se caracterizaron por presentar sus mayores abundancias en aguas someras del Banco de Campeche, donde se registró una mayor temperatura, salinidad y biomasa zooplanctónica; por el contrario N. punctata y nueve especies más, mostraron sus mayores abundancias sobre la estrecha plataforma continental de los estados de Veracruz y Tabasco. Esta distribución de las medusas presupone una importante relación con las características hidrológicas y la productividad planctónica local. Al respecto, Gasca et al. (1995) y Orozco-Zúñiga (2005), han señalado que la mayor productividad secundaria zooplanctónica en el sur del Golfo de México, se localiza por lo general sobre el Banco de Campeche y frente a la desembocadura de los principales sistemas costeros durante el otoño y que esa disponibilidad trófica es capaz de mantener númerosas poblaciones de zooplancteres, incluyendo al componente gelatinoso (Gasca et al.,1995). Asimismo, durante esta época las cálidas aguas superficiales podrían contribuir a un mejor crecimiento y sobrevivencia de las medusas.
En este sentido, Zamponi et al. (1990), han observado que en las zonas costeras de elevada productividad planctónica y en aguas poco profundas, las hidromedusas (Cunina, Aglaura y Liriope) y algunas escifomedusas (Nausithoe), pueden incrementar sus poblaciones considerablemente, debido a que sus aguas cálidas promueven un rápido crecimiento, aprovechando los nichos disponibles que favorecen la presencia constante de especies altamente dinámicas como las que constituyen estos grupos. Por tal motivo, una disponibilidad abundante de alimento, la ausencia de competidores y depredadores, aunado a las aguas cálidas que promueven un rápido crecimiento, trae consigo un incremento exponencial en la biomasa de las medusas. Recientemente, el incremento poblacional de algunas especies de medusas y de zooplancteres ha sido relacionado con los recientes cambios del medio oceánico a nivel global (Brodeur et al., 2002). Asimismo, Bohannan y Lenski (2000) han señalado que la productividad de un ecosistema puede afectar de manera directa la importancia relativa de la competencia y la depredación, incidiendo en la estructura de la comunidad y en la abundancia de las especies. De acuerdo con Mills (1995), en años recientes, debido a los cambios globales asociados a la actividad antropogénica, algunos ecosistemas poco productivos que estaban caracterizados por el escaso número de organismos del fitoplancton, de copépodos pequeños y de otras especies de zooplancteres, han incrementado notablemente su productividad, lo que ha generado un aumento considerable en las poblaciones de medusas y ctenóforos.
Además, el patrón de circulación local es otro factor que podría influir sobre la distribución de las especies. En este sentido, Monreal-Gómez y Salas de León (1990), han señalado que en el sur del Golfo de México durante el otoño, la circulación geostrófica conforma un giro ciclónico, por lo que los materiales orgánicos e inorgánicos pueden ser transportados hacia el oriente. Esta circulación dominante, podría generar frentes de masa de agua con características oceánicas y transportar la fauna de esta zona a las áreas más cercanas a la costa; asimismo, el giro ciclónico en el Banco de Campeche, podría funcionar como un mecanismo de retención del zooplancton gelatinoso. Al respecto, Gaughan y Fletcher (1997), han señalado este mismo efecto sobre el macrozooplancton carnívoro en la corriente de Leeuwin, en las aguas costeras del suroeste de Australia.
Por otro lado, la presencia del Aglantha elata sobre el talud continental, presupone la existencia de un afloramiento de aguas profundas al borde del talud.
La medusofauna del sur del Golfo de México estuvo caracterizada por la co-ocurrencia de formas neríticas (66%) y oceánicas (34%), cuyas abundancias estuvieron relacionadas con las condiciones oceanográficas del sur del golfo. Lo anterior se reflejó en el área de estudio al registrar una mayor riqueza y diversidad en la zona oceánica (30 y 2.6) y una menor, en la zona de influencia lagunar (23 y 1.9). Al respecto Livingston (1984), ha señalado que la diversidad del zooplancton en sistemas costeros es típicamente baja; este rasgo ha sido asociado con la alta variabilidad del sistema en términos de salinidad y temperatura en comparación del ambiente más estable que representa el medio oceánico. Esto tiene como consecuencia que las especies mejor adaptadas al medio sean ecológicamente dominantes; lo anterior quedó demostrado al encontrar un grado de repartición de la dominancia de 0.42 en la zona de influencia lagunar, indicando la dominancia de una sola especie (L. tetraphyla) con más del 45% de IVI. Por el contrario, un valor de 0.6 supone una menor dominancia y mayor repartición de recursos entre más de una especie (L. tetraphyla, N. punctata, C. octonaria y O. medium) en la zona oceánica.
Las diferencias en la riqueza de medusas entre las tres zonas oceanográficas consideradas en este estudio, presuponen la existencia de diferentes comunidades en las cuales cada grupo está integrado por especies con afinidades ecológicas similares que, por su transporte, aprovechan mejor las condiciones favorables del medio para su desarrollo. En este sentido, el valor intermedio obtenido de riqueza (14) registrado en el Banco de Campeche, deriva de especies holoplanctónicas que no dependen de una fase polipoide sésil además de poseer altas tasas de reproducción asexual como es el caso de los géneros Liriope, Cunina y Aglaura, especies del ámbito oceánico que, al encontrar condiciones más favorables, incrementan su reproducción y por ende, su número. Así, una mayor disponibilidad de recursos alimentarios sobre la plataforma del Banco, favorece una mayor abundancia de especies tanto neríticas como oceánicas. Los elevados valores de diversidad y riqueza registrados en la zona oceánica, con respecto a los obtenidos en la zona de influencia lagunar y de plataforma, manifiestan definitivamente la influencia de un medio estable y de pocas perturbaciones para la comunidad de medusas. Los bajos valores de diversidad observados sobre el Banco de Campeche, en áreas afectadas por el enriquecimiento mediante afloramientos de origen anticiclónico (giros), y la cercanía con afluentes epicontinentales, incide en una menor complejidad estructural de la comunidad de medusas con la clara dominancia de una especie, L. tetraphylla, una especie euritérmica y eurihalina. En el medio con influencia lagunar, dominado por especies meroplanctónicas, se registró el valor más bajo de riqueza, no así el de diversidad, un valor intermedio entre las zonas oceánica y la de plataforma.
Además, Zamponi et al. (1990), han señalado que las especies dominantes registradas en la Bahía de la Ascensión, en el Caribe Mexicano, presentaron características eurihalinas y euritérmicas, por lo que son estas especies las que llegan a dominar diversas áreas geográficas. Finalmente, aún cuando L. tetraphylla fue la especie dominante en las tres zonas, las especies con una menor abundancia (especies meroplanctónicas) marcaron mejor la variabilidad de la comunidad en el área estudiada. Los cambios espaciales en el medio oceanográfico superficial, la disponibilidad del alimento, así como la biología de las especies, fueron los principales factores que afectaron la distribución de los diferentes componentes estructurales de la comunidad de medusas en el sur del Golfo de México.
Agradecimientos
Los autores agradecen a la tripulación del B/O Justo Sierra, así como a Laura Celis y Alma Almaral del Laboratorio de Plancton de la Unidad Académica Puerto Morelos del Instituto de Ciencias del Mar y Limnología de la UNAM por las facilidades otorgadas para la corroboración de algunas especies de medusas. Asimismo, se agradece a Margarita Ornelas por su apoyo en la separación de muestras. El presente trabajo forma parte del trabajo de investigación de maestría del primer autor en el CINVESTAV Unidad Mérida.
Referencias
ALVARIÑO, A. 1985. Predation in the plankton realm: mainly with reference to fish larvae. Investigaciones Marinas, 2(1): 1-22. [ Links ]
BOHANNAN, B. J. M. & R. E. LENSKI. 2000. The relative importance of competition and predation varies with productivity in a model community. American Naturalist 156: 329-340. [ Links ]
BOLTOVSKOY, D. 1999. South Atlantic Zooplankton. Vol.I-II, Backhuys Publishers, Leiden, The Netherlands. 1707 p. [ Links ]
BRODEUR, R., H. SUGISAKI & G.L. HUNT. 2002. Increases in jellyfish biomass in the Bering Sea: implications for the ecosystem. Marine Ecology Progress Series 233: 83-103. [ Links ]
CAPURRO, F. L. & J. REID. 1972. Contributions on the physical oceanography of the Gulf of Mexico. Vol. 2. Texas A. & M. University, Texas. 288 p. [ Links ]
CORREIA, V. K. 1992. Medusas del Golfo de México y litoral norte/noreste de Brasil (Ocurrencia y distribución). Tesis de Doctorado en Ciencias (Biología), Facultad de Ciencias, UNAM, México. 284 p. [ Links ]
DE LA CRUZ, A. 1972. Zooplancton de la región sureste del Golfo de México. Ciencias Biológicas, Serie 4(24): 1-54 [ Links ]
DE LA CRUZ, AGÜERO, G. 1993. ANACOM. Sistema para el análisis de comunidades en computadoras personales. In: V Congreso Latinoamericano sobre Ciencias del Mar, La Paz B. C S., México. [ Links ]
DE LA LANZA-ESPINO, G. (ED.). 1991. Oceanografía de los Mares Mexicanos. AGT Editor, S.A., México. 569 p. [ Links ]
DE LA LANZA-ESPINO, G. 2001. Características Físico-Químicas de los Mares de México. Temas Selectos de Geografia de México. Instituto de Geografía, UNAM, México. 149 p. [ Links ]
FLORES-COTO, C., F. FLORES-HERNÁNDEZ, F. ZAVALA-GARCÍA, V. ARENAS-FUENTES, A. MONREAL-GÓMEZ & D. SALAS DE LEÓN. 2000a. Oceanic and neritic ichthyoplankton at the edge of the continental shelf in the southern Gulf of Mexico. Gulf and Caribbean Research 12: 31-35. [ Links ]
FLORES-COTO, C., R. MARTÍNEZ-GUTIÉRREZ, M. GONZÁLEZ-FÉLIX, L. SANVICENTE-AÑORVE & F. ZAVALA-GARCÍA. 2000b. Annual variation of ichthyoplankton assemblages in neritic waters of the southern Gulf of Mexico. Caribbean Journal of Science 36(3-4): 233-243. [ Links ]
GASCA, R., E. SUÁREZ-MORALES & I. CASTELLANOS. 1995. Biomasas zoopláncticas en aguas superficiales del Golfo de México durante el verano e invierno de 1991. Caribbean Journal of Science 31(1-2): 128-140. [ Links ]
GAUGHAN, D. J. & W. J. FLETCHER. 1997. Effects of the Leeuwin Current on the distribution of carnivorus macrozooplankton in the shelf waters off Southern Western Australia. Estuarine Coastal and Shelf Science 45(1): 88-97. [ Links ]
GÓMEZ-AGUIRRE, S. 1986. Ocurrencia de Chiropsalmus quadrumanus (Schyphozoa: Cubmedusae) en la Laguna de Términos. Anales del Instituto de Biolología, UNAM, Serie Zoología 57(2): 227-234. [ Links ]
HARRIS, R. P., P. H.WIEBE, J. LENZ, H. R. KJOLDAL & M. HUNTLEY. 2000. Zooplankton Methodology Manual. Academic Press, London. 684 p. [ Links ]
HOUDE, D. E. & N. CHITTY. 1976. Seasonal abundance and distribution of zooplankton, fish eggs and fish larvae in the Eastern Gulf of Mexico, 1972-74. NOAA Technical Reports NMFS SSRF-701. 18 p. [ Links ]
KRAMP, P. L. 1959. The Hydromedusae of the Atlantic Ocean and adjacent waters. Dana Reports 46: 1-283. [ Links ]
KRAMP, P. L. 1961. Synopsis of the Medusae of the World. Journal of Marine Biology 40: 1-469. [ Links ]
LIVINGSTON, R. J. 1984. The ecology of the Apalachicola Bay System: an estuarine profile. U. S. Fish and Wildlife Service Office. Biological Services FWS/OBS. Tallahassee (USA), pp. 82-105. [ Links ]
MAYER, A. G. 1910. Medusae of the World. Hydromedusae, Vol. I. Carnegie foundation, Washington, D.C. 208 p. [ Links ]
MCCUNE, B. & J. B. GRACE. 2002. Analysis of Ecological Communities. MjM Software Design. Gleneden Beach, Oregon. 300 p. [ Links ]
MILLS, C. E. 1995. Medusae, Siphonophores, and Ctenophores as planktivorus predators in changing global ecosystem. ICES Journal of Marine Science 52(3-4): 575-581. [ Links ]
MONREAL-GÓMEZ, M. A. & D. A. SALAS DE LEÓN. 1990. Simulación de la circulación en la Bahía de Campeche. Geofísica Internacional 29: 101-111. [ Links ]
OROZCO-ZÚÑIGA, M. E. 2005. Análisis de la variación espacial de la comunidad del zooplancton y su relación con las características oceanográficas en el sur del Golfo de México (otoño de 1999). Tesis Profesional. Facultad de Ciencias, UNAM, México. 77 p. [ Links ]
PHILLIPS, P. J. 1972. The Pelagic Cnidaria of the Gulf of Mexico: zoo-geography, ecology and systematics. Ph.D. Thesis, Texas A. & M. University, USA. 212 p. [ Links ]
PURCELL, E. J. 2003. Predation on zooplankton by large jellyfish, Aurelia aurita, Cyanea capillata and Aequorea aequorea, in Prince William Sound, Alaska. Marine Ecology Progress Series 246: 137-152. [ Links ]
RAMÍREZ, F. C. 1977. Planktonic Indicators of Argentine Shelf and Adjacent Areas. Proceedings of the Symposium of Warm Water Zoopankton. Special Publication. UNESCO/NIO. UK., pp. 65-68. [ Links ]
RAMÍREZ, F. C. & M. O. ZAMPONI. 1981. Hydromedusae. In: Boltovskoy, D. (Eds.). Atlas del Zooplanton del Atlántico Sudoccidental y Métodos de Trabajo con el Zooplancton Marino. Publicaciones Especiales del Instituto Nacional de Investigaciones en Desarrollo Pesquero. Mar de la Plata, Argentina, pp. 443-469. [ Links ]
RAMOS, G. & L. SEGURA-PUERTAS. 2004. Seasonal occurrence of reef-related medusae (Cnidaria) in the western Caribbean Sea. Gulf and Caribbean Research 16: 1-9. [ Links ]
SEGURA-PUERTAS, L. 1984. Morfología, sistemática y zoogeografía de las medusas (Cnidaria: Hydrozoa y Scyphozoa) del Pacífico Tropical Oriental. Anales del Instituto de Ciencias del Mar y Limnología, UNAM 8: 1-320. [ Links ]
SEGURA-PUERTAS, L. 1992. Medusae (Cnidaria) from the Yucatán Shelf and Mexican Caribbean. Bulletin of Marine Science 51: 353-359. [ Links ]
SEGURA-PUERTAS, L. 2003. Las medusas (Cnidaria) del Caribe mexicano. In: Barreiro-Guemes, M. T., M. E. Meave del Castillo, M. Signoret-Poillon & M. G.Figueroa-Torres (Eds.). Planctología Mexicana. Sociedad Mexicana de Planctología, A. C., México, pp. 213-228. [ Links ]
SEGURA-PUERTAS, L. & U. ORDÓÑEZ-LÓPEZ. 1994. Análisis de la comunidad de medusas (Cnidaria) de la región Oriental del Banco de Campeche y Caribe Mexicano. Caribbean Journal of Science 30(1-2): 104-115. [ Links ]
SEGURA-PUERTAS, L., E. SUÁREZ-MORALES & L. CELIS. 2003. A cheklist of the Medusae (Hydrozoa, Scyphozoa and Cubozoa) of Mexico. Zootaxa 194: 1-15. [ Links ]
SUÁREZ-MORALES, E., R. GASCA & L. SEGURA-PUERTAS. 1999. Medusae (Cnidaria) assemblages off the Caribbean coast of Mexico. Journal of Coastal Research 15: 140-147. [ Links ]
SUÁREZ-MORALES, E., R. GASCA, L. SEGURA-PUERTAS & D.C. BIGGS. 2002. Planktonic cnidarians in a cold-core ring in the Gulf of Mexico. Anales del Instituto de Biología, UNAM, Serie Zoología 73(1): 19-36. [ Links ]
TOVAR, J. E. 2003. Medusas (Cnidaria) del arrecife Mahahual, Quintana Roo, México. Tesis de Maestría, ECOSUR, Chetumal, México. 47 p. [ Links ]
VANNUCCI, M. 1957. On Brazilian Hydromedusae an their distribution in relation to different water masses. Boletim do Instituto Oceanográfico, Universidade de Sâo Paulo 8: 23-109. [ Links ]
ZAMPONI, M. O. & E. SUÁREZ. 1991. Algunas hidromedusas del mar Caribe mexicano, con la descripción de Tetratoporpa siankaanensis gen. et sp. nov. (Narcomedusae: Aeginidae). Spheniscus 9: 41-46. [ Links ]
ZAMPONI, M. O., E. SUÁREZ-MORALES & R. GASCA-SERRANO. 1990. Hidromedusas (Coelenterata:Hydrozoa) y Escifomedusas (Coelenterata: Scyphozoa) de la Bahía de la Ascensión, Reserva de la Biosfera de Sian Ka'an. In: Navarro, D.L. & J.G. Robinson (Eds.). Diversidad Biológica en la Reserva de la biosfera de Sian Ka'an, Quintana Roo México. Centro de Investigaciones de Quintana Roo/Program of Studies in Tropical Conservation, University of Florida, pp. 99-107. [ Links ]
ZAR, J. H. 1988. Biostatistical Analysis. Second edition. Prentice-Hall, Inc, USA. 718 p. [ Links ]

