










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.17 no.1 Ciudad de México abr. 2007
Artículos
Changes in the crustacean community of a tropical rocky intertidal shore: is there a pattern?
Cambios en la comunidad tropical de crustáceos en la zona rocosa intermareal: ¿hay un patrón?
Carmen Hernández Alvarez and Fernando Alvarez
Colección Nacional de Crustáceos, Instituto de Biología, Universidad Nacional Autónoma de México, Apartado Postal 70-153, México 04510, D.F., México. E-mail: cha30mx@yahoo.com.mx
Recibido: 20 de junio de 2006
Aceptado: 12 de enero de 2007
Abstract
A tropical rocky intertidal community in Montepio, southern Veracruz, Mexico, was sampled throughout a year to determine the extent of the changes in species composition and abundance. The study focused on the crustacean community, of which 49 species were identified. The community was characterized by a high species replacement rate, with 38.7% of the species appearing only once, 10% appeared twice, 16.3% were present in three different samples, and the remaining 34.7% were collected in 4 or more monthly samplings. Species diversity (H') varied constantly, while evenness remained relatively high and constant. An ordination analysis indicates that the crustacean community was dominated by the occurrence of rare species (47%). A cluster analysis, based on the Bray-Curtis similarity coefficient, shows that only a maximum of two consecutive samplings can have more than 50% similarity, reflecting the constant changes that occur in this community. When compared to the other numerically important phyla in the community, crustacean densities ranked third after annelids and mollusks. With such highly variable species composition and the absence of defined seasonal patterns, we propose that non-local processes, such as the strong winter winds ("northers") and a long season of tropical storms, are acting upon the community preventing the establishment of species for long periods of time and maintaining the biodiversity.
Key words: Rocky intertidal shore, community, Crustacea, species replacement, Gulf of Mexico.
Resumen
La comunidad de la zona intermareal rocosa tropical de Montepío, al sur de Veracruz, México, se muestreó por un año para determinar los cambios en la composición de especies y su abundancia. El estudio se enfocó a la comunidad de crustáceos, de la cual se identificaron 49 especies, ésta se caracterizó por un alto reemplazo de especies, con 38.7% de las especies que sólo se presentaron una vez, 10% presentes dos veces, 16.3% se capturaron en tres muestreos y el restante 34.7% fue recolectado en cuatro o más meses. La diversidad de especies (H') varió constantemente, mientras que la equitatividad se mantuvo con valores relativamente altos y constantes. Un análisis de ordenación indicó que la comunidad de crustáceos fue dominada por la presencia de especies raras (47%). El análisis de agrupamiento, basado en el coeficiente de similitud de Bray-Curtis, mostró que sólo un máximo de dos muestreos consecutivos pueden tener más del 50% de similitud, reflejando los constantes cambios que existen en esta comunidad. Cuando se comparó con otros phyla numéricamente importantes en la comunidad, la densidad de crustáceos se ubicó en tercer sitio, después de los anélidos y moluscos. Con una composición de especies tan variable y la ausencia de patrones estacionales definidos, se propone que procesos no locales, como los nortes y una larga temporada de tormentas tropicales, actúan sobre la comunidad evitando el establecimiento de especies por períodos largos de tiempo y manteniendo la diversidad.
Palabras clave: Zona intermareal rocosa, comunidad, Crustacea, reemplazo de especies, Golfo de México.
Introduction
Rocky intertidal communities have been intensely studied for a long time due to their accessibility and richness of natural resources. However, in spite of the long history of ecological studies conducted in this ecosystem, many questions related to the spatio-temporal structuring of the communities remain unexplored. Recent studies have identified important gaps in our knowledge about this ecosystem, including the use and relative importance of specific microhabitats (McKindsey & Bourget, 2001; Thompson et al., 2002), methodological approaches to categorize species richness at a regional scale (Benkendorff & Davies, 2002), and the importance of short term temporal variation (Olabarria & Chapman, 2002).
The number of species and variability in occurrence of this fauna is determined by their ability to colonize and withstand the changing conditions of this habitat at a local scale and their capacity to maintain interspecific interactions (Lewis, 1964; Connell, 1972; Stephenson & Stephenson, 1972; Little & Kitching, 1996; Raffaelli & Hawkins, 1996). In many cases, rocky intertidal habitats are affected by the prevailing hydrographic conditions, specially in areas influenced by upwellings (Menge et al., 1997; Menge, 2000).
Rocky shores are nursery and feeding areas for a number of species of crustaceans, fishes, and other vertebrates, which are interconnected with adjacent terrestrial and marine ecosystems (Rangeley & Kramer, 1995; Bradshaw et al., 1999; Burrows et al., 1999; Thompson et al., 2000). The populations inhabiting the rocky intertidal have been considered as open due to larval transport and recruitment from separate populations (Gaines & Roughgarden, 1985; Underwood & Fairweather, 1989; Menge, 1991; Small & Goslling, 2001). This "openness" produces a variable recruitment which combined with diverse abiotic factors create a spatio-temporal mosaic from the local to the regional scale (Lewis, 1976; Bowman & Lewis, 1977; Underwood et al., 1983; Hartnoll & Hawkins, 1985; Johnson et al., 1998; Underwood, 1999; Jenkins et al., 2001). The variability found in populations inhabiting the rocky intertidal may be due to different abiotic factors such as tidal regime, temperature, red tides, changes in sea level, storms, wave action, and fisheries (Crisp, 1964; Southgate et al., 1984; Underwood & Fairweather, 1989; Bustamante & Branch, 1996; Denny & Paine, 1998).
In the Mexican portion of the Gulf of Mexico, rocky intertidal communities are spatially limited, occurring mainly as: a) limestone formations, in the northwestern section of the Yucatan Peninsula; b) coral reefs, in Isla Lobos, the Port of Veracruz, Antón Lizardo, and Cayo Arcas; c) lava flows, present in Los Tuxtlas region; and d) man-made structures, ever more prominent such as docks, jetties and oil rigs (Britton & Morton, 1988; Hernández, 2002). The rest of the coastline (>80%), mainly formed by sand bars, is under the influence of enormous sediment loads, discharged by a number of important rivers. Under these circumstances, the remaining rocky intertidal habitat is both spatially restricted and always influenced to some extent by freshwater, nutrients and sediments from the coastal runoff.
The present study examines the variation in the intertidal community composition in Montepio, southern Veracruz, Mexico, established along an ancient lava flow that extends 60 m from the shore to a depth of 1.5 m. This study represents the first report on the composition and dynamics of the crustacean assemblage inhabiting a rocky intertidal shore in the southwestern Gulf of Mexico. A monthly sampling program was designed to survey the community at a short temporal scale. We hypothesized that strong changes in the community composition would be detected since the region is constantly under the influence of extreme weather conditions, namely: the intense winter winds ("northers") from November to February, high temperatures and risk of desiccation at low tide during the dry season from March to May, and the tropical storms typical of the rainy season from June to October. We analyzed the community focusing on the crustacean species, but considered other abundant groups (mollusks, annelids, sipunculids, and echinoderms) for a general comparison.
Materials and methods
Eleven monthly samplings were conducted on the lava flow at Montepio, from February 1996 to February 1997. No samples were obtained in October and December due to bad weather caused by the "northers". Water temperature and salinity were recorded in every sampling. The sampling procedure consisted in obtaining pieces of rubble formed by encrusting, tubiculous, and coral forming organisms. Three replicates were obtained at depths ranging between 0.5 and 1 m, and placed in bags individually underwater to minimize the loss of organisms. The rocks were weighed and their volume estimated in a graduated bucket. Each rock was fragmented until all the organisms present were obtained. Crustaceans, mollusks, echinoderms, and sponges were preserved in 70% Ethanol (EtOH), while polychaetes were preserved in 4% formalin. In a previous sampling, 10 rock samples weighing between 1.6 and 6.7 kg were analyzed to determine the optimum sample size. The maximum number of crustacean species (18) was obtained in rocks weighing between 3.5 and 4.1 kg, a weight that was adopted for the whole sampling program (Hernández, 2002). It was further estimated that 4 kg rocks had a volume of 1.2 to 2.2 liters.
All crustaceans were identified to species using appropriate keys for each group as follows: for amphipods, Barnard (1969), Bousfield (1973), Barnard & Barnard (1983); for isopods, Menzies & Kruczynski (1983) and Kensley & Schotte (1989); and for decapods, Rathbun (1930), Provenzano (1959), Gosner (1971), Chace (1972), Gore & Abele (1976), Dardeau (1984), Williams (1984), and Abele & Kim (1986). Abundance, density (number of organisms/ liter of sample, org/L), and biomass (grams of wet weight/liter of sample, g/L), were obtained for all species. All organisms were deposited in the National Crustacean Collection, Instituto de Biología, UNAM. Non-crustaceans were classified to phylum, and their densities and biomasses obtained.
The community analysis consisted in estimating species richness by taxonomic group and season and Shannon-Wiener's diversity index (H') and Pielou's evenness (J), to analyze temporal variation in species composition. A one way analysis of variance and Tukey's HSD tests were used to test, among the monthly samples, for significant differences in species richness, density, and biomass of crustaceans. The Olsmtead-Tukey ordination technique was used to obtain the relative importance with respect to abundance of each species in the community. This analysis was performed with the total sample density values (Steele & Torrie, 1992). A cluster analysis based on the Bray-Curtis similarity coefficient was performed; square root-transformed abundance data were used.
Results
Throughout the sampling period water temperature varied between 22 and 36ºC ( = 27.1ºC) in February and July, respectively, and salinity between 30 and 38 ( = 34.4) in July and April, respectively.
= 27.1ºC) in February and July, respectively, and salinity between 30 and 38 ( = 34.4) in July and April, respectively.
The whole invertebrate community at Montepio changed considerably at the phylum level. The five more abundant phyla were Annelida, Mollusca, Crustacea, Sipunculida, and Echinodermata. Three distinct density peaks were recorded: the first one occurred in April, with a major contribution of annelids with 149 org/L; the second peak appeared in July, with the mollusks contributing with 147 org/L; and the third peak, recorded in September, had two main contributions by the mollusks and annelids with 156 and 206 org/L, respectively (Fig. 1). A significant feature of this pattern is that the maximum crustacean density, obtained in June with 109 org/L, does not represent a density maximum for the whole community. The density for the whole community, considering these five phyla, ranged from a minimum of 111 org/L in February to a maximum of 482 org/L in September.
Species richness
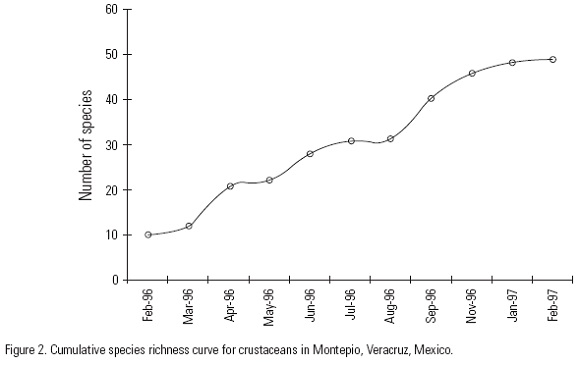
The total sample was composed of 3657 crustaceans with a biomass of 677.7 g, representing 4 orders, 16 families, 31 genera, and 49 species (Table 1). The shape of the cumulative species richness curve suggests that still more species can be found in this microhabitat (Fig. 2). The families with more species in the total sample were: Porcellanidae (7), Alpheidae (7), Gammaridae (6), Hyalidae (5), Menippidae (4), and Corophiidae (4). The 10 remaining families contributed with three or less species each (Table 1). Species richness varied significantly throughout the study, from an average of 3 species in March to 15 in September (Anova, F = 3.34, P = 0.0087) (Fig. 3). A multiple comparison indicates that September is significantly different from the two months poorest in species March and May (Tukey HSD, P = 0.008 and P = 0.043, respectively; Fig. 3).
Diversity
Species diversity (H') varied constantly, with a tendency to increase, from 0.4 in February 1996 to 1.1 in January 1997 (Fig. 4). Evenness remained relatively high and constant, except for two periods, February and June 1996. In February 1996 only 9 species were collected, with the hermit crab Clibanarius antillensis Stimpson, 1862, numerically dominating the community (n = 244). Interestingly, this is the highest number of specimens collected for any individual species in one monthly sample. However, C. antillensis was only present in this period, disappearing for the rest of the year. In June 1996, an undescribed species of amphipod, Elasmopus sp 1, and the anomuran crab Neopisosoma angustifrons (Benedict, 1901), were the most abundant with 113 and 109 individuals, respectively; while the remaining 17 species present in this sample had an average of 12 individuals.
The species turnover pattern shows that a maximum of 58% of the total species present in two consecutive samplings are shared (January-February), while the two most dissimilar consecutive samplings (February-March) only shared 27% of the species. From another angle, 19 (38.7%) of the 49 species recorded were collected only once, five (10.2%) species appeared twice, eight (16.3%) species were present in three different samples, and the remaining 17 (34.7%) species were collected in 4 or more monthly samplings. Only two species, the menippid crab Eriphia gonagra (Fabricius, 1781) and the grapsid Pachygrapsus transversus (Gibbes, 1850) were present in all the samples.
Density
Regarding monthly crustacean density, although it varied considerably among samples, from a minimum average of 25 org/L in March to 109 org/L in June, no significant differences were detected (Fig. 5). Mean density values are low from February to May, peaking in June, and remaining above 50 org/L the rest of the period. Although the biomass increases considerably from February (5 g/L) to July (80 g/L), decreasing seven months later to comparable initial levels (February, 9 g/L), no significant differences were obtained either (Fig. 6). The biomass peak obtained in July is due to the presence of the barnacle Tetraclita floridana Pilsbry, 1916, and the crabs N. angustifrons and P. transversus, which appeared in large numbers, besides being relatively large organisms.
Community analysis
The Olmstead-Tukey association technique shows that, for the complete study period, 47% of the species were rare, 33% dominant, 16% indicator, and 4% common in terms of density (Table 1). Considerable variation, not presented here, was observed as to what species were in each of the four categories every month. This result is consistent with the high species turnover rate that characterizes this community.
The cluster analysis shows that the monthly samples cannot be associated in any defined general pattern (Fig. 7). Since the Bray-Curtis similarity coefficient considers the abundance of each species in relation to the total sample, it reflects up to some extent changes in community composition. In that sense, the abrupt changes in species composition from one sample to the next one produces in Montepio a different number of groups depending on the level of similarity. Between 60 and 80% similarity only four groups consisting of two months each can be recognized: April and May, characterized by low densities; January and February 1997, with high species richness and intermediate densities; July and August share intermediate values for both species richness and density; and June and September combine intermediate to high species richness and density.
Discussion
A clear controversy emerges between those studies that find that non-local processes, such as changing environmental conditions and recruitment intensity (Roughgarden et al., 1988; Hutchinson & Williams, 2001), rather than local processes or within the community interactions, such as competition for microhabitats (Minton & Gochfeld, 2001; Neil, 2001), are the main factors structuring intertidal communities and maintaining biodiversity. In this study, the very high species replacement rate (Gore et al., 1978; Domínguez, 2006), which may be preventing the establishment of long-term interspecific interactions, suggests that non-local processes such as storm mediated stress and stochastic recruitment of species, are key factors shaping the community. Similarly, Bertness & Leonard (1997) have already summarized these ideas for intertidal communities recognizing that at low levels of physical stress predation pressure may be high, becoming rare at high levels of physical stress.
The discontinuous nature of the rocky intertidal habitat along the coast of the southwestern Gulf of Mexico may also be contributing to the high species replacement pattern observed. The recruitment of different species throughout the study may be reflecting the absence of large source populations for most species that could predictably produce propagules to recolonize disturbed areas. A likely explanation is that most of the recorded species follow a "fugitive species" strategy, settling in newly opened spaces, and growing and reproducing rapidly before the next disturbance modifies the abiotic conditions and they disappear from that rocky shore.
The analysis of the obtained Results is based on species diversity, abundance and density of each species, and their patterns of occurrence. The percentage of rare species (47%), which were present in three or less samplings with very low densities, suggests that recruitment intensity is low for a high proportion of species in this group. In contrast however, six of the 19 species that occurred only once throughout the study, were present in large numbers most probably as a result of high recruitment rates (Robles, 1997). This ample variation in recruitment patterns may also be preventing the establishment of interspecific interactions such as competition and predation, precluding also the presence of a keystone predator (Robles et al., 1995; Robles, 1997).
Two crustacean species that were present in all samples and that could have been part of a significant community structuring interaction as predators, were the crabs Eriphia gonagra and Pachygrapsus transversus. Both have extensive geographical ranges and reproduce virtually all year-round, being the commonest species at every site where they are recorded (Williams, 1984; Cuesta & Schubart, 1998). Both species could be considered omnivorous, feeding on algae, or preying upon a wide variety of organisms (Gore et al., 1978). Eriphia gonagra had medium to low densities, being abundant only in March (26 org/L), the most species-poor month; whereas P. transversus was relatively abundant (25 - 37 org/L) from July to January, a period characterized by high species richness and variable densities. Thus, P. transversus could be a key species, with a significant role in the shaping of the community as the most important invertebrate predator.
The constant variation detected in species composition in this study contrasts with other Results from rocky intertidal habitats in subtropical and temperate areas where community composition is stable and predictable. Gore et al. (1978) described as stable and predictable the decapod species composition associated with a sabellariid worm reef in Florida. Based on stomach contents, they argued that the constant supply of food provided by the sabellariid worms was the main factor that maintained the community without noticeable changes. Other studies have identified predictable temperature regimes (Thomson & Lehner, 1976), predictable recruitment rates and population growth of keystone species (Flores & Negreiros-Fransozo, 1999), and the strength of certain predator-prey interactions (Farrell, 1988), as the factors that prevent communities from undergoing constant changes.
Regarding the species richness estimates for this ecosystem, it is clear that with such high species replacement rate and a large percentage of rare species, only continuous samplings for more than a year would start to produce reliable Results. The cumulative species curve presented here indicates that more species are still to be added. Supporting this interpretation, diversity (H') levels do not clearly increase with time in this community, suggesting that a typical succession process is not taking place here. Rather, diversity seems to completely depend on the random recruitment of species, which is taking place continuously throughout the year. The constant presence of disturbances in tropical intertidal ecosystems, rather than predictable seasonal changes, has been found to be responsible for the maintenance of high diversity in other tropical systems (Ben-Eliahu et al., 1988).
Crustaceans are usually conceived as a numerically dominant group in coastal habitats; however, our Results show that annelids and mollusks have higher densities than crustaceans. A salient point is that the maximum density obtained for crustaceans does not coincide with the maximum for the whole community. When analyzing community structure in tropical rocky intertidal sites, the difficulties imposed by the taxonomy of each group and the countless possible interactions among all the species often promote that the analysis be restricted to a small set of taxa. However, as shown with the Results presented herein, interpretations about community composition can be seriously biased, overestimating the importance of the studied groups.
Acknowledgements
We thank J. C. Molinero, R. Robles, J. Calderón, J. L. Bortolini and J. L. Villalobos for their help in the field and laboratory work; and the Estación Los Tuxtlas, Instituto de Biología, Universidad Nacional Autónoma de México, for their logistic support. Drs. E. Escobar-Briones and L. A. Soto contributed with many stimulating Discussions. The first author was supported by a Consejo Nacional de Ciencia y Tecnología scholarship through the doctoral program at UNAM, (schoolarship #95881)
References
ABELE, L. G. & W. KIM, 1986. An illustrated guide to the marine decapod crustaceans of Florida. Department of Environmental Regulation, Florida State University 8(1): 1-760. [ Links ]
BARNARD, J. L., 1969. The families and genera of marine gammaridean Amphipoda. United States National Museum. Washington 271: 1-535. [ Links ]
BARNARD, J. L. & C. M. BARNARD, 1983. Freshwater amphipoda of the world. Hayfield Associates, Virginia, 830 p. [ Links ]
BEN-ELIAHU, M. N., U. N. SAFRIEL & S. BEN-TUVIA, 1988. Environmental stability is low where polychaete species diversity is high: quantifying tropical vs temperate within-habitat features. Oikos 52: 255-273. [ Links ]
BENEDICT, J. E., 1901. The anomuran collections made by Fisho Hawk Expedition to Puerto Rico. Bulletin of the U.S. Fish Commission 20(2): 129-148 [ Links ]
BENKENDORFF, K. & A. R. DAVIES, 2002. Identifying hotspots of molluscan species richness on rocky intertidal reef. Biodiversity and Conservation 11: 1959-1973. [ Links ]
BERTNESS, M. D. & G. H. LEONARD, 1997. The role of positive interactions in communities: lessons from intertidal habitats. Ecolog 78: 1976-1989. [ Links ]
BOUSFIELD, E. L., 1973. Shallow-Water Gammaridean Amphipoda of New England. Comstock Publishing Associates, London, 312 p. [ Links ]
BOWMAN, R. S. & J. R. LEWIS, 1977. Annual fluctuations in the recruitment of Patella vulgata L. Journal of the Marine Biological Association of the United Kingdom 57: 793-815. [ Links ]
BRADSHAW, C. J. A., C. M. THOMPSON, L. S. DAVIS & C. LALAS, 1999. Pup density related to terrestrial habitat use by New Zealand fur seals. Canadian Journal of Zoology 77: 1579-1586. [ Links ]
BRITTON, J. C. & B. MORTON, 1988. Shore Ecology of the Gulf of Mexico. University of Texas Press, Austin. 387 p. [ Links ]
BURROWS, M. T., K. KAWAI & R. N. HUGHES, 1999. Foraging by mobile predators on a rocky shore: underwater TV observations of movements of blennies Liphophrys pholis and crabs Carcinus maenas. Marine Ecology Progress Series 187: 237-250. [ Links ]
BUSTAMANTE, R. H. & G. M. BRANCH, 1996. Large scale patterns and trophic structure of southern African rocky shores: The roles of geographic variation and wave exposure. Journal of Biogeography 23: 339-351. [ Links ]
CHACE, F. A., JR., 1972. The shrimp of the Smithsonian-Bredin Caribbean Expedition with a summary of the West Indian shallow-water species (Crustacea: Decapoda: Natantia). Smithsonian Contributions to Zoology 98: 1-179. [ Links ]
CONNELL, J. H., 1972. Community interactions on marine rocky intertidal shores. Annual Review of Ecology and Systematics 3: 169-192. [ Links ]
CRISP, D. J., 1964. The effects of the severe winter of 1962-1963 on marine life in Britain. Journal of Animal Ecology 33: 165-210. [ Links ]
CUESTA, J. A. & C. D. SCHUBART, 1998. Morphological and molecular differentiation between three allopatric populations of the littoral crab Pachygrapsus transversus (Gibbes, 1850) (Brachyura: Grapsidae). Journal of Natural History 32: 1499-1508. [ Links ]
DARDEAU, M. R., 1984. Synalpheus shrimps (Crustacea: Decapoda: Alpheidae). I The Gambarelloidae group with a description of a new species. Memoirs of the Hourglass Cruises 7(2): 1-125. [ Links ]
DENNY, M. W. & R. T. PAINE, 1998. Celestial mechanics, sea-Ievel changes, and intertidal ecology. Biological Bulletin 194: 108-115. [ Links ]
DOMÍNGUEZ, D., 2006. Biodiversidad de la criptofauna coralina (Crustacea) de isla Peña, playa Careyeros e islas Marietas, Nayarit, México, México. M.Sc. Thesis, Universidad Nacional Autónoma de México, 80 p. [ Links ]
.FLORES, A. A. V. & M. L. NEGREIROS-FRANSOZO, 1999. On the population biology of the mottled shore crab Pachygrapsus transversus (Gibbes, 1850) (Brachyura, Grapsidae) in a subtropical area. Bulletin of Marine Science 65: 59-73. [ Links ]
GAINES, S. D. & J. ROUGHGARDEN, 1985. Larval settlement rate, a leading determinant of structure in sessile marine species. Nature 360: 579-580. [ Links ]
GIBBES, L. R. 1850. On the carcinological collections of the United States. Proceedings of the American Association for the Advancement of Science 2: 167-201. [ Links ]
GORE, R. H. & L. G. ABELE, 1976. Shallow water porcelain crabs from the Pacific coast of Panama and adjacent Caribbean waters (Crustacea: Anomura: Porcellanidae). Smithsonian Contributions to Zoology 237: 1-759. [ Links ]
GORE, R. H., L. E. SCOTTO & L. J. BECKER, 1978. Community composition, stabillity, and trophic partitioning in decapod crustaceans inhabiting some subtropical sabellarid worm reefs. Bulletin of Marine Science 28: 221-248. [ Links ]
GOSNER, K., 1971. Guide to Identification of Marine and Estuarine Invertebrates. Wiley-Interscience, John Wiley & Sons, Inc., New York, 693 p. [ Links ]
HARTNOLL, R. G. & S. J. HAWKINS, 1985. Patchiness and fluctuations on moderately exposed rocky shores. Ophelia 24: 53-63. [ Links ]
HERNÁNDEZ, M C., 2002. Variabilidad estacional de la comunidad de crustáceos de la facie rocosa intermareal en Montepío, Veracruz, México. M.Sc. Tesis de Maestría, Universidad Nacional Autónoma de México, 70 p. [ Links ]
HUTCHINSON, N. & G. A. WILLIAMS, 2001. Spatio-temporal variation in recruitment on a seasonal, tropical rocky shore: The importance of local versus non-local processes. Marine Ecology Progress Series 215: 57-68. [ Links ]
JENKINS, S. R., F. ARENAS, J. ARRONTES, J. BUSSELL, J. CASTRO, R. A. COLEMAN, S. J. HAWKINS, S. KAY, B. MARTINEZ, J. OLIVEROS, M. F. ROBERTS, S. SOUSA, R. C. THOMPSON & R. G. HARTNOLL, 2001. European-scale analysis of seasonal variability in limpet grazing activity and microalgal abundance. Marine Ecology Progress Series 211: 193-203. [ Links ]
JOHNSON, M. P., M. T. BURROWS & S. J. HAWKINS, 1998. Individual based simulations of the direct and indirect effects of limpets on a rocky shore Fucus mosaic. Marine Ecology Progress Series 169: 179-188. [ Links ]
KENSLEY, B. & M. SCHOTTE, 1989. Guide to the Marine Isopod Crustaceans of the Caribbean. Smithsonian Institution Press. Washington, 293 p. [ Links ]
LEWIS, J. R., 1964. The Ecology of Rocky Shores. London, UK: English Universities Press. [ Links ]
LEWIS, J. R., 1976. Long-term ecological surveillance: practical realities in the rocky littoral. Oceanography and Marine Biology, Annual Review 14: 371-390. [ Links ]
LITTLE, C. & J. A. KITCHING, 1996. The Biology of Rocky Shores. Oxford, UK, Oxford University Press. [ Links ]
MCKINDSEY, C. W. & E. BOURGET, 2001. Diversity of a northern rocky intertidal community: The influence of body size and succession. Ecology 82: 3462-3478. [ Links ]
MENGE, B. A., 1991. Relative importance of recruitment and other causes of variation in rocky intertidal community structure. Journal of Experimental Marine Biology and Ecology 145: 69-100. [ Links ]
MENGE, B. A., 2000. Top-down and bottom-up community regulation in marine rocky intertidal habitats. Journal of Experimental Marine Biology and Ecology 250: 257-289. [ Links ]
MENGE, B. A., B. A. DALEY, P. A. WHEELER, E. DAHLHOFF, E. SANFORD & O. T. STRUB, 1997. Benthic-pelagic links and rocky intertidal communities: Bottom-up effects on top-down control? Proceedings of the National Academy of Sciences of the United States of America 94: 14530-14535. [ Links ]
MENZIES, R. J. & W. L. KRUCZYNSKI, 1983. Isopod Crustacea (exclusive of Epicaridea). Florida Departament of Natural Resources Marine Research Laboratory. VI(1): 1-126. [ Links ]
MINTON, D. & D. J. GOCHFELD, 2001. Is life on a tropical shore really so hard?: The role of abiotic factors in structuring a supralittoral molluscan assemblage. Journal of Shellfish Research 20: 77-483. [ Links ]
NEIL, K. M., 2001. Microhabitat segregation of co-existing gastropod species. Veliger 44: 294-300. [ Links ]
OLABARRIA, C. & M. G. CHAPMAN, 2002. Inconsistency in short-term temporal variability of microgastropods within and between two different intertidal habitats. Journal of Experimental Marine Biology and Ecology 269: 85-100. [ Links ] Provenzano, A. J., 1959. The shallow-water hermit crabs of Florida. Bulletin of Marine Science of the Gulf and Caribbean 9(4): 349-420. [ Links ]
RAFFAELLI, D. & S. HAWKINS, 1996. Intertidal Ecology. London, UK: Chapman and Hall, 372 p. [ Links ]
RANGELEY, R. W. & D. L. KRAMER, 1995. Tidal effects on habitat selection and aggregation by juvenile Pollock Pollachius virens in the rocky intertidal zone. Marine Ecology Progress Series 126: 19-29. [ Links ]
RATHBUN, M. J., 1930. The cancroid crabs of America of the families Euryalidae, Portunidae, Atelecyclidae, Cancridae and Xanthidae. Bulletin of the Smithsonian Institution, United States National Museum. Washington 152: 1-609. [ Links ]
ROBLES, C. D., 1997. Changing recruitment in constant species assemblages: Implications for predation theory in intertidal communities. Ecology 78: 1400-1414. [ Links ]
ROBLES, C. D., R. SHERWOOD-STEPHENS & M. ALVARADO, 1995. Responses of a key intertidal predator to varying recruitment of its prey. Ecology 76: 565-579. [ Links ]
ROUGHGARDEN, J., S. GAINES & H. POSSINGHAM, 1988. Recruitment dynamics in complex life cycles. Science 241: 1460-1466. [ Links ]
SMALL, M. P. & E. M. GOSLING, 2001. Population genetics of a snail species complex in the British Isles: Littorina saxatilis (Olivi), L. neglecta Bean and L. tenebrosa (Montagu), using SSCP analysis of Cytochrome-B gene fragments. Journal of Molluscan Studies 67: 69-80. [ Links ]
SOUTHGATE, T., K. WILSON, T. F. CROSS, & A. A. MYERS, 1984. Recolonization of a rocky shore in S.W. Ireland following a toxic bloom of the dinoflagellate Gyrodinium aureolum. Journal of the Marine Biological Association of the United Kingdom 64: 485-492. [ Links ]
STEELE, R. G. D. & J. H. TORRIE, 1992. Bioestadística: principios y procedimientos. McGraw-Hill, México, 622 p. [ Links ]
STEPHENSON, T. A. & A. STEPHENSON, 1972. Life Between the Tidemarks on Rocky Shores. San Francisco, USA: W.H. Freeman, 425 p. [ Links ]
THOMPSON, R. C., S. R. JENKINS, & J. A. BUSSELL, 2000. A method for recording predator-prey encounters between crabs and limpets using wax replicas. Journal of Marine Biological Association of the United Kingdom 80: 633-638. [ Links ]
THOMPSON, R. C., T. P. CROWE & S. J. HAWKINS, 2002. Rocky intertidal communities: past environmental changes, present status and predictions for the next 25 years. Environmental Conservation 29: 168-191. [ Links ]
THOMSON, D. A. & C. E. LEHNER, 1976. Resilience of a rocky intertidal fish community in a physically unstable environment. Journal of Experimental Marine Biology and Ecology 22: 1-29. [ Links ]
UNDERWOOD, A. J. 1999. History and recruitment in structure of intertidal assemblages on rocky shores: an introduction to problems for interpretation of natural change. In: M. Whitfield, J. Matthews & C. Reynolds (Eds.). Aquatic Life Cycle Strategies, Survival in a Variable Environment, pp. 79-96. Plymouth, UK. Marine Biological Association of the United Kingdom. [ Links ]
UNDERWOOD, A.J. & P.G. FAIRWEATHER, 1989. Supply-side ecology and benthic marine assemblages. Trends in Ecology and Evolution 4: 16-20. [ Links ]
UNDERWOOD, A. J., E. J. DENELY, & M. J. MORAN, 1983. Experimental analyses of the structure and dynamics of mid-shore rocky intertidal communities in New South Wales. Oecologia 56: 202-219. [ Links ]
WILLIAMS, A. B., 1984. Shrimps, lobsters, and crabs of the Atlantic coast of the eastern United States, Maine to Florida. Smithsonian Institution Press: 550 p. [ Links ]

