










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.16 no.2 Ciudad de México ago. 2006
Caracterización estacional de las condiciones físico-químicas y de productividad primaria fitoplanctónica de dos lagunas costeras tropicales del estado de Chiapas, México
Seasonal characterization of the physicochemical conditions of two tropical coastal lagoons of the state of Chiapas, Mexico
Francisco José Gutiérrez Mendieta, Francisco Varona-Cordero y Francisco Contreras Espinosa
Laboratorio de Ecosistemas Costeros, Departamento de Hidrobiología, DCBS. Universidad Autónoma Metropolitana-Iztapalapa. Av. San Rafael Atlixco No. 186 Col. Vicentina. Apdo Postal 55-535. C.P. 09340. México D. F. e-mail: fgm@xanum.uam.mx
Recibido: 8 de marzo de 2005
Aceptado: 9 de enero de 2006
RESUMEN
El objetivo de este trabajo fue el de evaluar en diferentes meses las principales variables ambientales, así como la biomasa y productividad primaria fitoplanctónica en dos lagunas costeras del estado de Chiapas, así como determinar sus posibles diferencias entre las dos épocas climáticas características de la región. El estudio abarcó varios ciclos anuales (1990-1991; 1997-2003). Los resultados muestran que las descargas de los principales ríos que desembocan en las lagunas determinan el establecimiento en las lagunas de características ambientales distintas para las dos principales épocas climáticas, secas y lluvias. Durante secas, la disminución de la precipitación y como consecuencia la baja influencia fluvial, aunada al intercambio mareal, genera en la laguna altas temperaturas y salinidades, así como concentraciones elevadas de nutrientes, que favorecen una alta biomasa y productividad primaria fitoplanctónica. En el caso de la época de lluvias, el alto aporte fluvial disminuye la salinidad e introduce nutrientes al sistema que son utilizados por el fitoplancton, favoreciendo también altas biomasas y productividades; sin embargo, debido al bajo tiempo de residencia, muchos de estos materiales son acarreados fuera del sistema hacia la zona costera adyacente.
Palabras clave: Caracterización estacional, multianual, laguna costera, Chiapas.
ABSTRACT
The objective of this study was to evaluate in several months, the main environmental variables and the phytoplankton biomass and production of two coastal lagoons in the state of Chiapas, and to determine the possible differences between the two main climatic seasons. The study was carried out during several yearly cycles (1990-1991; 1997-2003). The results show that the discharge of the main rivers flowing into these lagoons determines the establishment of different environmental characteristics during the main climatic seasons, dry and wet. During the dry season, the limited fluvial input, together with the tidal influence, generates stable environmental conditions, characterized by high temperatures and salinities, and high nutrient concentrations which favor high phytoplankton biomass and production. On the contrary, during the wet season, river input diminish salinities and increases nutrients which are used by the phytoplankton, promoting high biomasses and productions. However, due to the low residence time, many of this materials are exported out of the system to the adjacent coastal zone.
Key words: Seasonal characterization, multiannual, coastal lagoon, Chiapas.
INTRODUCCIÓN
Las lagunas costeras y estuarios se ubican entre los ecosistemas de mayor productividad, la cual depende de muchos factores, entre los que se encuentran la disponibilidad de luz y nutrientes para la comunidad fitoplanctónica. Estos factores a su vez, se ven modificados por los aportes fluviales y los intercambios mareales, los cuales alternarán su dominancia en función de las principales épocas climáticas que se presentan a lo largo del año.
La limitación de la penetración de la luz por la turbidez en ambientes ricos en nutrientes es uno de los factores que más se cita como un mecanismo regulador de la producción fitoplanctónica (Cloern, 1996; Santamaría del Ángel et al., 1996; Gameiro et al., 2004). La resuspensión de los sedimentos por la marea y los aportes de éstos por los ríos son factores importantes que modifican la concentración de materiales en suspensión y, por lo tanto, la profundidad de la zona fótica en la columna de agua.
La formación y acumulación de la biomasa fotosintética en ambientes estuarinos, depende no solo de la productividad del fitoplancton, sino también de otros factores como la herbivoría por parte del zooplancton y la exportación, resuspensión y depositación de sedimentos. La relación que existe entre el flujo de los ríos y la acumulación de la biomasa fitoplanctónica es compleja. Por ejemplo, aportes fluviales importantes pueden estimular a la producción primaria, al introducir nutrientes al sistema; sin embargo, sólo se podrá dar el desarrollo de un florecimiento de fitoplancton cuando la tasa de acumulación exceda a la perdidas, esto es, bajo condiciones de escasos aportes fluviales y largos tiempos de residencia como los que se presentan durante la temporada de secas (Gameiro et al., 2004); siendo lo contrario durante la época de lluvias (altos aportes fluviales y bajos tiempos de residencia).
A pesar de lo anterior y de la abundante información relacionada con el estudio de estos factores en zonas costeras templadas, poco se ha analizado en regiones costeras tropicales, en las que se reconoce la presencia de dos épocas climáticas características (secas y lluvias) (Alongi, 1998), así como el hecho de que la influencia más episódica de los ríos (Eyre & Balls,1999), genera una estructura de salinidad más dinámica y una mayor variabilidad en los aportes de nutrientes. Por lo anterior se hace indispensable efectuar estudios enfocados a establecer las condiciones físicas y químicas que caracterizan a estos ambientes durante las temporadas de secas y lluvias a fin de contribuir a su conocimiento ecológico y productivo. Asimismo, el conocimiento de las características y variaciones naturales de estas lagunas es necesario si posteriormente se quieren documentar los cambios que se presenten en estos ecosistemas como consecuencia de las actividades humanas.
A diferencia de otras latitudes, en México existen pocos ambientes estuarinos que hayan sido muestreados de manera continua (p. ej. Estero Punta Banda BCS, Lara-Lara & Alvarez-Borrego, 1975; Martínez & Garate, 1997; Laguna de Celestún, Herrera-Silveira, 1999). El caso de las lagunas costeras de Chantuto-Panzacola y Carretas-Pereyra en la costa chiapaneca no es la excepción, ya que existe poca información disponible previa a 1990 (Castañeda & Contreras, 2004). Estas lagunas son ambientes muy importantes de los que dependen una gran cantidad de recursos biológicos y comunidades humanas, además de que forman parte de la Reserva de la Biosfera de "La Encrucijada" (SEMARNAP, 1999). Estos ecosistemas están sujetos cada vez más a mayores presiones como consecuencia de las actividades humanas, por lo que se hace indispensable conocer adecuadamente sus características físicas y químicas. Con base en lo anterior los objetivos del presente trabajo fueron: a) caracterizar estacionalmente las variables ambientales y de productividad primaria fitoplanctónica (PPF) de los sistemas lagunares de Carretas-Pereyra y Chantuto-Panzacola con base en los muestreos efectuados durante los periodos de 1990-91 y 1997-2003; b) determinar las diferencias entre las variables y la PPF para las principales épocas climáticas; c) analizar el grado de variabilidad interanual de las características ambientales y de PPF, para sustentar la importancia y necesidad de la realización de monitoreos a largo plazo, que permitan una mejor comprensión del funcionamiento de las lagunas costeras mexicanas.
MATERIALES Y MÉTODOS
Área de estudio. El sistema lagunar-estuarino Carretas-Pereyra (C-P) está situado en los 93° 06' y 93° 15' de longitud oeste y entre los 15° 23' y 15° 32' de latitud norte. El sistema está conformado por varias lagunas someras que llevan el nombre de Pereyra, Carretas, Bobo y Buenavista, un cordón estuarino conocido como el Palmarcito y una boca de comunicación permanente con el mar (Fig. 1A), con una extensión de cuerpos lagunares de 3,696 ha (SEMARNAP, 1999). En este complejo lagunar desembocan de norte a sur cinco ríos principales: Pijijiapan, Echegaray, Margaritas, Bobo y Progreso.
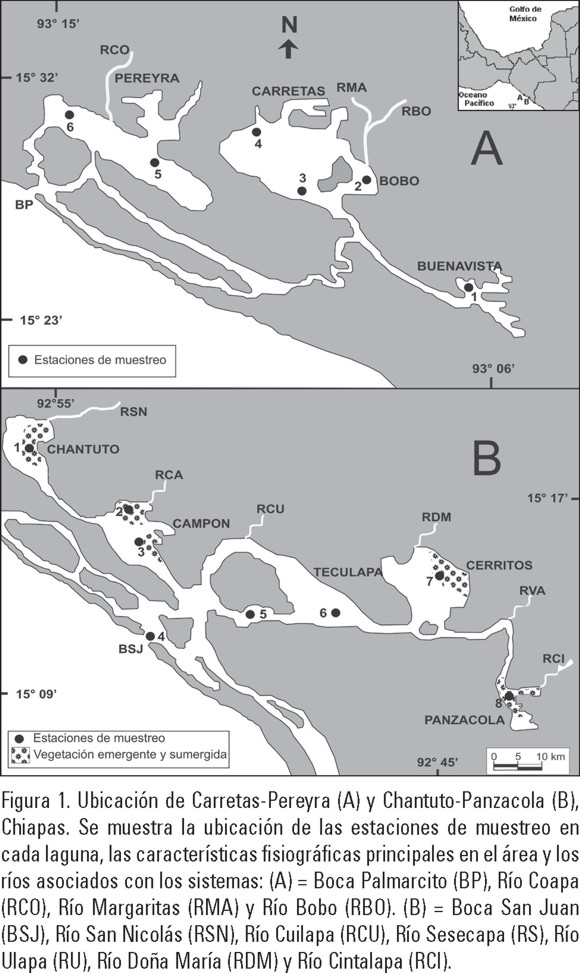
El sistema lagunar Chantuto-Panzacola (CH-P) se localiza en los 92° 45' y 92° 55' de longitud oeste y los 15° 09' y 15° 17' de latitud norte y está conformado por cinco lagunas: Chantuto, Campón, Teculapa, Cerritos y Panzacola con una extensión total de los cuerpos de agua de 18 000 ha. Presenta una boca de comunicación con el mar conocida como San Juan y un largo cordón estuarino paralelo a la barrera arenosa llamado El Hueyate (Fig. 1B). En este ecosistema desembocan seis ríos principales: San Nicolás (Payucal), Ulapa, Cacaluta, Doña María, Cintalapa y Vado Ancho.
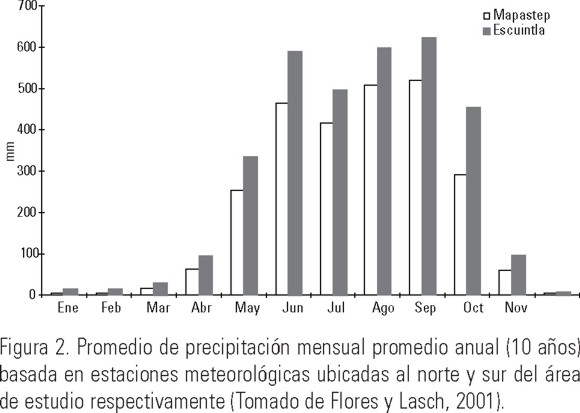
El régimen pluvial en la zona está determinado por la humedad proveniente del Océano Pacífico. El estiaje está definido entre los meses de noviembre a abril con menos del 10% de la tasa anual de precipitación en el período, mientras que las lluvias se presentan entre mayo y octubre. La precipitación anual promedio es de 2,310 mm, de los cuales el 95% del total se distribuye de mayo a octubre (Fig. 2). La marea es de tipo mixto semidiurno con una amplitud de 1.25 m en la Boca de Palmarcito y una hora de bajamar y pleamar casi simultánea para toda la zona (Márquez et al,. 2006).
Procedimiento de campo. Desde el año de 1990 se estableció en cada sistema una red de estaciones (9 en CP y 10 en CHP; Fig.1 A y B) las cuales fueron muestreadas (4-6 veces por año) con la finalidad de establecer la variación estacional de las principales variables físico-químicas a lo largo de los ciclos anuales. Se seleccionaron las diferentes zonas de cada sistema tomando en cuenta la influencia de ríos, mar y áreas con diferente vegetación (manglares y vegetación emergente y sumergida). Debido a lo somero de las lagunas (promedio de 1.15 m), en cada estación se tomaron las muestras a 1 m o menos de profundidad.
La temperatura se determinó con un termómetro de cubeta con una precisión de ± 0.1° C; la transparencia con un disco de Secchi y el espesor de la capa eufótica (Zeu) se obtuvo con base en la profundidad del disco de Secchi (Zsd) mediante la fórmula:
Zeu = - Zsd ln(0.01)/1.44 (Kirk, 1994).
El oxígeno disuelto se determinó por medio del método de Winkler (Strickland & Parsons, 1972); la salinidad se cuantificó empleando un refractómetro marca American Optical con una precisión de ±1.0 ups. A partir del año de 2003, la temperatura, la salinidad, el oxígeno disuelto y la turbidez se evaluaron con una sonda marca Horiba modelo U-22, calibrada al inicio de cada muestreo.
Para la cuantificación de nutrientes (NH4+, NO3-, NO2-, PO43- y P tot) se obtuvieron muestras de agua empleando una botella van Dorn. Las muestras se filtraron en el campo con filtros GF/F, y se mantuvieron a baja temperatura (4°C) hasta su procesamiento en el laboratorio. Paralelamente se colectaron y filtraron en la lancha entre 100-250 ml de agua para cuantificar la clorofila a como estimador de la biomasa fitoplanctónica.
Para la evaluación de la productividad primaria se utilizó el método de la botella clara y oscura (Gaarder & Gran, 1927), siguiendo las sugerencias propuestas por Brower y Zar (1981), manteniendo las botellas en incubación en un recipiente de acrílico transparente de 30 L a bordo de la lancha y cubierto con una malla que reduce la luz aproximadamente 20% con el fin de evitar la fotoinhibición de la fotosíntesis y simular las condiciones naturales de la laguna. Las muestras fueron incubadas durante tres horas, basados en experiencias previas que han mostrado que este tiempo es suficiente para registrar cambios detectables en la producción de oxígeno en las muestras (Varona-Cordero & Gutiérrez, 2003). Regularmente se efectuaron recambios del agua en el incubador con el fin de evitar el sobrecalentamiento debido a la insolación. La descarga de los ríos (m3/s) se obtuvo de los datos proporcionados por la CNA (1999).
Procedimiento de laboratorio. Los nutrientes se analizaron siguiendo las técnicas propuestas por Strickland y Parsons (1972). El índice N:P (inorgánico) se calculó dividiendo las formas de nitrógeno inorgánicas entre los ortofosfatos (Redfield, 1958).
La cuantificación de la clorofila a se realizó mediante la técnica de SCOR-UNESCO (1980), extrayendo con acetona al 90%, en la oscuridad durante 24 horas. Para la cuantificación se empleó un espectrofotómetro marca Beckman® Du-65.
Análisis estadísticos. La variabilidad temporal de los diferentes factores físico-químicos (transformados log N+1 y sin transformar) se analizó mediante pruebas de hipótesis para determinar las diferencias significativas entre los meses y años de muestreo e identificar los meses característicos de cada época climática.
Adicionalmente, para determinar la significancia de tales diferencias, se ejecutaron pruebas de hipótesis dependiendo del tipo de distribución (no paramétrico Kruskall-Wallis, así como un análisis de varianza de una vía ANOVA, siendo el tiempo el factor) con un nivel de significancia de α <0.05 (Zar, 1997).
RESULTADOS
Sistema Chantuto-Panzacola. El sistema CH-P presentó una profundidad promedio de 1.11 m; sin embargo, a pesar de la influencia que los ríos pueden tener sobre la profundidad de la laguna, no existieron diferencias entre las dos épocas climáticas. Los registros de profundidad de la laguna mostraron que el sistema ha perdido una parte importante de su volumen, ya que en 1990 se determinó una profundidad promedio de 1.44 m, la cual diminuyó a 1.03 m hasta el año de 2003. Resalta de manera importante el período de 2000-2002 en el que la profundidad promedio fue menor a los 0.90 m.
La salinidad fue la variable que indicó claramente la diferencia entre las dos épocas climáticas típicas de la zona (secas y lluvias). El promedio anual de salinidad fue de 14.3 ups, presentándose diferencias significativas en el promedio que fue de 21.6 ups en secas y 9.73 ups en lluvias.
Debido al incremento en la entrada de materia orgánica por parte de los ríos en la temporada de lluvias (Torres et al., 2006), la concentración de oxígeno fue baja (2.9 ml/L ); esta concentración de oxígeno, se vio reflejada en el porcentaje de saturación, que presentó un intervalo de 52-80 % sin diferencias importantes entre las dos épocas climáticas (Tabla 1).
El efecto del océano adyacente y el de los ríos influyó también en el pH de la laguna, por ejemplo en la época de secas se incrementaron los valores, disminuyendo durante las lluvias. Sin embargo, no se presentaron diferencias significativas entre ambas épocas.
En lo que se refiere a las concentraciones de nutrientes, en este sistema lagunar la forma nitrogenada más abundante fueron los nitratos, con una concentración promedio de 5.76 μM; en términos porcentuales entre un 40% y 70% del nitrógeno total inorgánico (NH+4 + N0-3 + NO-2) fue aportado por este compuesto. El amonio fue la segunda forma en abundancia, con una concentración en general menor a 5.0 μM. La concentración promedio de ortofosfatos fue 4.57 μM, sin existir diferencias significativas entre las dos épocas climáticas.
Se determinó una biomasa fitoplanctónica de 20.85 μg/L, no existiendo diferencias significativas en la concentración de este pigmento entre las dos épocas climáticas. Los valores de la producción primaria fitoplanctónica fueron de 166 mg C/m3/hr, en promedio presentando valores similares entre ambas épocas.
Sistema Carretas-Pereyra. El sistema C-P en promedio fue más somero (0.78 m) que CH-P; existiendo diferencias significativas entre las épocas de secas y lluvias con respecto a la profundidad (0.2 m).
En términos de la salinidad, este sistema fue comparativamente más marino que el de CH-P, con un promedio anual de salinidad de 16.2 ups. Asimismo, presentó diferencias significativas entre secas y lluvias (21.4 y 12.7 ups), respectivamente (Tabla 1). Al igual que en CH-P las temperaturas fueron muy elevadas en las dos épocas del año, existiendo diferencias mínimas pero significativas entre la época de secas (31.2 °C) y lluvias (32.3 °C).
La concentración de oxígeno fue más elevada que en CH-P (4.0 ml/l), con un intervalo del porcentaje de saturación de 56-113%. En lo que se refiera a los valores de pH se presentaron diferencias significativas entre secas (7.8) y lluvias (7.6).
El amonio en este sistema presentó una concentración promedio de 3.97 μM; siendo los nitratos, al igual que en CH-P, la primera forma nitrogenada en abundancia, con una concentración promedio de 9.07 μM. La concentración promedio de fósforo, fue más elevada en C-P (7.46 μM) que en CH-P, existiendo diferencias significativas entre las dos épocas climáticas (5.12 y 9.09 μM en secas y lluvias, respectivamente), mostrando el efecto por parte de los ríos sobre este nutriente.
En el sistema C-P se registró una mayor biomasa fitoplanctónica (36.7 μg/L) que en CH-P y no existieron diferencias significativas entre las dos épocas climáticas. Como resultado de lo anterior, la producción primaria fitoplanctónica fue más alta (194 mg C/m3/hr), aunque con valores similares en ambas épocas.
DISCUSIÓN
Comportamiento estacional. El descenso de la profundidad en el período 2000-2002, se puede atribuir a dos fenómenos importantes que se registraron durante este lapso de tiempo. Por un lado, durante 1998 se registraron unas de las mayores precipitaciones en la zona (1200 mm en cinco días), como consecuencia, la mayor parte de los ríos se salieron de su cauce y una gran cantidad de sedimentos fue transportada desde la cuenca y depositada en el fondo de las lagunas. Por otro lado, posterior al evento de 1998, hubo un periodo de sequía (Galván et al., 2006), lo cual, aunado a los sedimentos depositados en las lagunas, condujeron a esta disminución de la profundidad. A partir de ese momento, los sistemas han tenido un comportamiento cíclico alternando condiciones de baja profundidad (0.40 m) en secas y alta (1.0 m) en lluvias. En el caso de sistemas someros como los aquí estudiados, la disminución de profundidad puede representar la pérdida de más de un 15 % del volumen lagunar, lo cual puede tener consecuencias importantes en el funcionamiento y permanencia del sistema.
Aunque la temperatura presentó diferencias significativas en C-P entre las épocas de secas y lluvias, en general el intervalo interanual en ambas lagunas fue estrecho (30.7 ± 2.06 y 31.8 ± 1.74; promedio y desviación estándar para CH-P y C-P, respectivamente), cercano al máximo ambiental y característico de zonas tropicales (Alongi,1998). Las temperaturas fueron elevadas en las dos épocas del año, estando siempre por arriba de los 30 °C en promedio, esta situación se puede explicar por la existencia de dos máximos de temperatura ambiental que corresponden al doble paso del sol por el cenit, de los cuales el primero se presenta en mayo y el segundo en agosto (Cardoso, 1979), correspondiendo cada uno de estos a las épocas de secas y lluvias, respectivamente.
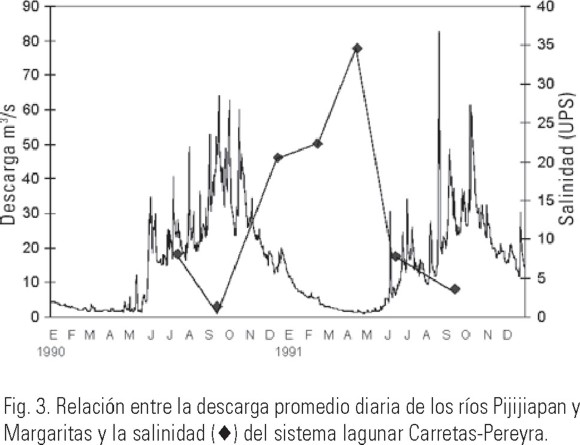
Por el contrario, la salinidad mostró una clara diferencia entre cada una de las épocas climáticas, indicando la influencia que las entradas de agua dulce, provenientes de los ríos y la lluvia, tienen sobre el sistema (Tabla 1). En lo que se refiere al patrón de distribución de la salinidad, diversos estudios (Alongi, 1998; Troccoli et al., 2004) han demostrado que uno de los principales factores que afectan su distribución es la entrada de agua dulce proveniente de los ríos y la precipitación. Además del efecto que tienen sobre la salinidad (Fig. 3), las descargas fluviales también ocasionan que disminuya la transparencia, especialmente en las desembocaduras de los ríos (Varona-Cordero & Gutiérrez, 2003), mientras que durante la época de secas, la transparencia fue homogénea en la totalidad de las lagunas (Fig. 4).
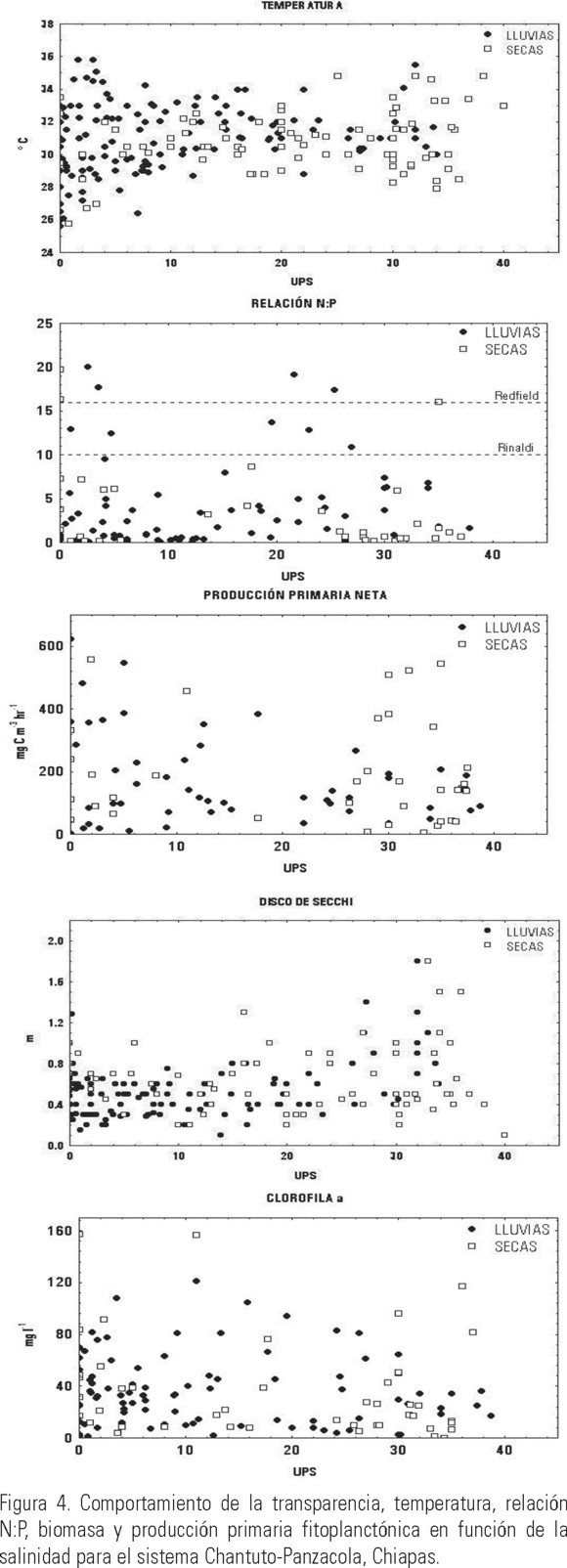
En el caso de la transparencia (disco de Secchi), la mayor penetración de la luz estuvo asociada con altas salinidades, especialmente durante la época de secas en CH-P (Fig. 4). Por el contrario, las menores transparencias están asociadas a bajas salinidades, denotando la entrada de agua proveniente de los ríos la cual de manera general acarrea sedimentos y materiales en suspensión, limitando el paso de la luz en la columna de agua.
La concentración promedio anual y por época climática (Tabla 1) del oxígeno disuelto indica que ambos sistemas presentaron una deficiencia importante de este gas, principalmente a bajas salinidades, pudiendo estar asociada con una alta demanda biológica de oxígeno (DBO) debido a un mayor contenido de material orgánico aportado por los ríos (Eyre, 1998). Por ejemplo, Torres et al. (2006), reportaron para ambos sistemas una mayor concentración de carbono orgánico en sedimentos durante la época de lluvias que en la de secas, aunque las diferencias no fueron significativas.
Con respecto al pH, se determinaron condiciones ácidas asociadas con bajas salinidades y por lo tanto relacionadas con las entradas de agua dulce, mientras que las características básicas reflejaron la influencia del océano sobre la laguna, especialmente durante la época de secas en ambos sistemas.
La influencia de los ríos sobre las lagunas también se observó en las concentraciones de nutrientes (amonio, nitratos + nitritos y fosfatos), las cuales de manera general fueron superiores en la temporada de lluvias (> aporte fluvial) (Varona-Cordero & Gutiérrez, 2003), y disminuyeron a altas salinidades durante la época de secas (Figs. 4 y 5).
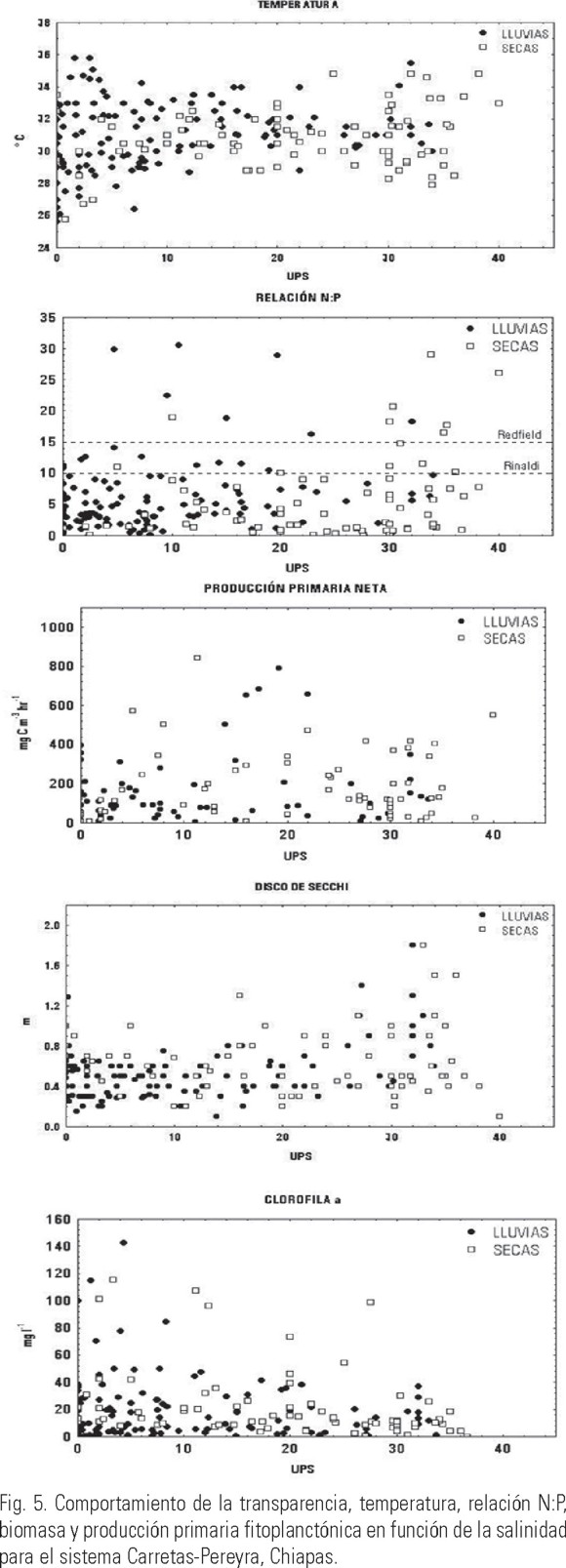
La relación N:P, indicó de manera general para ambos sistemas una deficiencia relativa de nitrógeno para los productores primarios (Redfield, 1958); inclusive, el promedio anual y por época climática mostró en ambos sistemas valores promedio menores a 6.0, lo que indica una fuerte limitación por dicho nutriente. Por el contrario, en ocasiones puntuales este índice sugiere una limitación por parte del fósforo hacia la comunidad fitoplanctónica. Esta fluctuación entre limitación por nitrógeno y fósforo, ha sido documentada en otros sistemas costeros tanto templados como tropicales (Alongi, 1998; Eyre & Balls, 1999).
El comportamiento estacional de la biomasa fitoplanctónica (clorofila a) se relacionó con las concentraciones de nutrientes, ya que en ambos sistemas las mayores biomasas estuvieron asociadas a bajas salinidades y altas concentraciones de nutrientes (Figs. 4 y 5). Sin embargo, la producción primaria generada por esta biomasa presentó un comportamiento diferente en cada sistema. En el caso de C-P, la distribución en las épocas climáticas fue de forma unimodal, es decir, con producciones altas en salinidades intermedias y menores a altas y bajas salinidades (Fig. 4). Esta distribución sugiere que en las zonas cercanas a las desembocaduras de los ríos con baja transparencia y salinidad, y altas concentraciones de nutrientes, la producción fitoplanctónica estuvo limitada por la baja penetración de la luz (Cloern, 1996; Gameiro et al., 2004). En las zonas de salinidad intermedia, la turbidez disminuyó, y las concentraciones de nutrientes favorecieron altas biomasas y productividades. En la zona de influencia marina, la concentración de nutrientes fue menor, disminuyendo por lo tanto la productividad. En el caso del sistema CH-P, esta distribución no es tan evidente, registrándose altas productividades tanto en las épocas secas como de lluvias; no obstante hay una tendencia de que las mayores productividades se presentaron con bajas salinidades y altas concentraciones de nutrientes. En el caso de los dos sistemas estudiados, los resultados confirman la influencia de las dos épocas climáticas sobre los sistemas lagunares. Contreras (2001), estableció que en lagunas costeras mexicanas, durante esta época se alcanzan altas biomasas fitoplanctónicas como efecto directo del aumento en los nutrientes, pero éstas son efímeras por los bajos tiempos de residencia del agua dentro de la laguna y su rápida exportación hacia el mar (Eyre, 1998, Lucas et al., 1999).
Se ha establecido para diversos sistemas estuarinos tropicales, caracterizados por presentar épocas de secas y lluvias, la influencia que tienen los aportes fluviales en el comportamiento hidrológico (Eyre, 1998; Eyre & Balls, 1999; Herrera-Silveira, 1994; Herrera-Silveira et al., 1999).
El caso de los sistemas costeros de Chiapas no es la excepción, los ríos que desembocan a estos sistemas lagunares, tienen un efecto fundamental sobre el comportamiento hidrológico, productivo y en general sobre todo el ecosistema. Estos son ríos de un cauce corto (< 50 km) a partir de su punto de origen (SEMARNAP, 1999). A lo largo de este cauce, pasan de aguas transparentes, altas pendientes y velocidades y lechos rocosos en la parte alta de la cuenca, a cauces con un alto contenido de sedimentos finos y baja pendiente en donde el cauce principal se pierde en pequeños arroyos de carácter dendrítico conformado por corrientes de cuarto orden (Flores & Lasch, 2001). Es en esta última zona en donde adquieren muchas de las características finales antes de depositar sus aguas en las lagunas (Galván & Márquez, 2006). El flujo es altamente variable y la mayor parte del material es transportado durante períodos de descarga cortos, dividiendo estacionalmente a las lagunas en las dos épocas características. Eyre y Balls (1999), establecieron para estuarios tropicales australianos, que durante la época de secas, se acumula en la cuenca mucho del material que será transportado por los ríos en la época de lluvias cuando se presentan las grandes avenidas (80-90% de los nutrientes y sedimentos suspendidos en los sistemas costeros). Esto se reflejara a su vez en las concentraciones de nutrientes en las partes oligohalinas de las lagunas, favoreciendo altas productividades fitoplanctónicas de manera similar a lo establecido en estuarios turbios de otras latitudes (Kocum et al., 2002). Por lo anterior, los extremos temporales que se presentan en la descarga de los ríos, ejercen un control marcado en la manera en que los materiales originados en tierra firme pasan por las lagunas y son exportados a la zona costera adyacente. Como ya se mencionó, los sistemas han perdido una parte importante de su volumen (Márquez et al., 2006).
Los resultados de este estudio, confirman la presencia de este comportamiento estacional para ambos sistemas en los que la semejanza geomorfológica de las dos lagunas (Fig. 1), la cercanía entre ellas (± 40 km) y la respuesta similar determinada por el efecto de las descargas fluviales, ocasionan que las fluctuaciones de los diferentes factores físicos y químicos fueran similares en ambos sistemas.
En conclusión, durante la época de secas hubo una limitada descarga fluvial y una elevada influencia marina, presentando los sistemas características eurihalinas, principalmente en las zonas más cercanas a las bocas de comunicación y partes intermedias de la laguna (Varona-Cordero, 2004). La mezcla de la columna de agua por la marea y el viento, favorecieron una resuspensión de los sedimentos, los cuales redujeron la transparencia de la columna de agua, incrementando las concentraciones de nutrientes (Monbet, 1992). Estas condiciones, aunadas al largo tiempo de residencia del agua (26 días; Varona-Cordero, 2004), le confieren a los sistemas una estabilidad ambiental que dio como resultado una alta biomasa y producción primaria fitoplanctónica, aunque es importante mencionar que la mayor parte de los valores de la relación N:P se encontraron dentro del intervalo de 5 a 10, el cual se puede interpretar como indistinto, esto es, que no representan una limitación real para la comunidad fitoplanctónica (Figs. 4 y 5; Rinaldi et al., 1992).
Durante la época de lluvias, por el contrario, se presentó una elevada influencia de los ríos sobre las lagunas, disminuyendo la salinidad (características oligohalinas), principalmente en las zonas cercanas a las descargas e incrementando la concentración de nutrientes. Este aumento favoreció altas biomasas y productividades en la columna de agua, pero su permanencia dentro del sistema puede ser efímera debido a los bajos tiempos de residencia del agua dentro de la laguna (2.31 días; Varona-Cordero, 2004) y su rápida exportación hacia el mar.
El presente análisis muestra la importancia de la realización de monitoreos a largo plazo lo cual permite una mejor caracterización de los dos sistemas lagunares.
AGRADECIMIENTOS
El presente trabajo fue apoyado parcialmente por el CONACYT bajo el convenio 400200-5-4282PT y por la Universidad Autónoma Metropolitana-Iztapalapa. Se agradecen también las sugerencias y comentarios recibidos por parte de la M. en C. Rocío Torres Alvarado y tres revisores anónimos, las cuales contribuyeron a mejorar la calidad del manuscrito.
REFERENCIAS
ALONGI, D.M. 1998. Coastal Ecosystem Processes. CRC Press. Boca Ratón. USA 419 p. [ Links ]
BROWER, J. E., & J. H. ZAR. 1981. Field and Laboratory methods for general ecology. WM.C Brown Company. 194 p. U.S.A. [ Links ]
CARDOSO, C.M.D. 1979. El clima de Chiapas y Tabasco. Instituto de Geografía. UNAM. México. 99 p. [ Links ]
CASTAÑEDA, L. 0. & F. CONTRERAS. 2004. Bibliografía comentada sobre los ecosistemas costeros mexicanos. Vol. II: Litoral del Pacífico. CONABIO, UAM-I CDELM. México. 495p. [ Links ]
CLOERN, J. E. 1996. Phytoplankton bloom dynamics in coastal ecosystems: a review with some general lessons from sustained investigation of San Francisco Bay, California. Revue of Geophysics 34: 127-168. [ Links ]
CNA, IMTA. COMISIÓN NACIONAL DEL AGUA. INSTITUTO MEXICANO DE TECNOLOGÍA DEL AGUA.1999. Extractor Rápido de Información Climátológica (ERIC) CD. Datos climatológicos. México. [ Links ]
CONTRERAS E. F. 2001. Caracterización de lagunas costeras mexicanas a través de variables ecológicas seleccionadas. Tesis de Doctorado. Universidad Autónoma Metropolitana Unidad Iztapalapa. División de Ciencias Biológicas y de la Salud. México. 95 p. [ Links ]
EYRE, R., 1998. Transport, retention and transformation of material in Australian Estuaries. Estuaries 21: 540-551. [ Links ]
EYRE, R. & P. BALLS. 1999. A comparative study of nutrient behaviour along the salinity gradient of tropical and temparate estuaries. Estuaries 22(2): 313-326. [ Links ]
FLORES, M. A. R., & C. L. LASCH. 2001. Herramientas de planificación para la conservación de sitios. Estudio caso: cuenca del Río Coapa, Municipio de Pijijiapan, Chiapas, México. SEMARNAT. México. 60 p. [ Links ]
GAARDER, T. & H. H. GRAN. 1927. Investigations of the production of plankton in the Oslo Fjord. Rapport. Conseil International sur le Exploration de la Mer. 42: 3-31. [ Links ]
GALVÁN, M. A. F. & A. Z. MÁRQUEZ. 2006. Descripción biofísica de la cuenca del Río Coapa, Chiapas. Hidrobiológica 16(2): En prensa. [ Links ]
GAMEIRO, C. P. CARTAXANA, M.T. CABRITA & V. BROTAS. 2004. Variability in chlorophyll and phytoplankton composition in an estuarine system. Hydrobiologia 525: 113-124. [ Links ]
HERRERA-SILVEIRA, J. A., 1994. Spatial heterogeneity and seasonal patterns in a tropical coastal lagoon. Journal of Coastal Research 10(3): 738-746 [ Links ]
HERRERA-SILVEIRA, J. A., M. B. MARTÍN & V. DÍAZ-ARCE. 1999. Variaciones del fitoplancton en cuatro lagunas costeras de Yucatán. Revista de Biología Tropical 47 (Supl.1): 47-56. [ Links ]
KIRK, J. T.O. 1994. Light and photosynthesis in Aquatic Ecosystems. Cambridge. Univ. Press. U.S.A. 509 p. [ Links ]
KOCUM, E., D.B. NEDWELL & G:C. UNDERWOOD. 2002. Regulation of phytoplankton primary production along a hypernutrified estuary. Marine Ecology Progress Series 231:13-22 [ Links ]
LARA-LARA, J. R. & S. ÁLVAREZ-BORREGO. 1975. Ciclo anual de clorofilas y producción orgánica primaria en Bahía San Quintín, B. C. Ciencias Marinas 2(1): 77-96. [ Links ]
LUCAS L. V., J.R. KOSEFF, S.G. MONISMITH, J.E. CLOERN & J.K. THOMPSON. 1999. Processes governing phytoplankton blooms in estuaries II: the role of horizontal transport. Marine Ecology Progress Series 187: 17-30. [ Links ]
MÁRQUEZ, G. A. Z., A. PÉREZ R. & L. G. CALVA B. 2006. Consideraciones batimétricas del sistema lagunar Carretas-Pereyra, Chiapas, México. Hidrobiológica 16(3): En prensa. [ Links ]
MARTÍNEZ, L. A. & I. L. GARATE. 1997. Variación diurna de la materia orgánica particulada en una laguna costera del Golfo de California. Revista de Biología Tropical 45: 1421-1428. [ Links ]
MONBET, Y. 1992. Control of phytoplankton biomass in estuaries: a comparative analysis of microtidal and macrotidal estuaries. Estuaries 15: 56-571 [ Links ]
REDFIELD, A. C., 1958. The biological control of chemical factors in the environment. American Science 46: 205-221. [ Links ]
RINALDI, A. G., MONTANARI.A. GHETTI, C. R. & A. FERRARI. 1992. Eutrophy and dystrophy in the Goro Lagoon. In: R. A. Volenweider, R. Marchetti & R. Viviani. (Eds.) Marine Coastal Eutrophication. Elsevier. 457-470. [ Links ]
SANTAMARÍA DEL ÁNGEL, E., MILLÁN-NÚÑEZ R. & G. DE LA PEÑA-NETTEL. 1996. Efecto de la turbidez en la productividad primaria en dos estaciones en el área del delta del Río Colorado. Ciencias Marinas. 22: 483-493. [ Links ]
SCOR-UNESCO. 1980. Determination of chlorophyll in seawater. UNESCO Technical Papers in Marine Science. France. 35 p. [ Links ]
SEMARNAP. 1999. Programa de Manejo. Reserva de la Biósfera "La Encrucijada. México. Instituto Nacional de Ecología- Secretaría el Medio Ambiente, Recursos Naturales y Pesca. México. 184 p. [ Links ]
SOLÓRZANO, L., 1969. Determination of ammonia in natural water by phenol-hypochlorite method. Limnology and Oceanography 14: 799-801. [ Links ]
STRICKLAND, J. & T. R. PARSONS. 1972. A practical handbook of seawater analysis. Fisheries Research Board of Canada. Canada. 310 p. [ Links ]
TORRES-ALVARADO, M.R., F.J. FERNÁNDEZ, I. A. BARRIGA S., & F. RAMÍREZ V. 2006. Dinámica de las bacterias anaeróbicas en las fases terminales de la mineralización de la materia orgánica en el sedimento de los ecosistemas Carretas-Pereyra y Chantuto-Panzacola. Hidrobiológica (16) 2: En prensa. [ Links ]
TROCCOLI,L.G., J.A. HERRERA-SILVEIRA & F.A. COMIN. 2004. Structural variations of phytoplankton in coastal seas of Yucatan, Mexico. Hydrobiologia 519: 85-102 [ Links ]
VARONA-CORDERO, F. 2004. Estudio comparativo de los cambios en la composición, distribución y abundancia de la comunidad fitoplanctónica de dos lagunas Costeras (Carretas-Pereyra y Chantuto-Panzacola) y su relación con las variaciones de los parámetros físicos y químicos. Tesis de Maestría. Universidad Autónoma Metropolitana-Iztapalapa, México. 127 p. [ Links ]
VARONA-CORDERO, F. & F. GUTIÉRREZ M. 2003. Estudio multivariado de la fluctuación espacio-temporal de la comunidad fitoplanctónica en dos lagunas costeras del estado de Chiapas. Hidrobiológica 13(3): 177-194. [ Links ]
ZAR, J.H. 1998. Biostatistical Analysis. 4th Ed. Prentice Hall, Englewoods California. U.S.A. 718 p. [ Links ]

