










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.13 no.1 Ciudad de México abr. 2003
Article
Ceratium balechii sp. nov. (Dinophyceae: Gonyaulacales) from the Mexican Pacific
Ceratium balechii sp. nov. (Dinophyceae: Gonyaulacales) del Pacífico Mexicano
Ma. Esther Meave del Castillo1, Ma. Eugenia Zamudio Resendiz1, Yuri B. Okolodkov1 and Isaías H. Salgado Ugarte2,3
1Dpto. de Hidrobiología. Universidad Autónoma Metropolitana-Iztapalapa; Av. San Rafael Atlixco 186. Col. Vicentina México, D.F. 09340. México. mem@xanum.uam.mx
2Dpto. de Biología. Universidad Autónoma Metropolitana-Iztapalapa; Av. San Rafael Atlixco 186. Col. Vicentina México, D.F. 09340. México.
3Carrera de Biología. Facultad de Estudios Superiores, Zaragoza, UNAM; Guelatao 66, Ejército de Oriente, Iztapalapa, 09230, D.F. México.
Recibido: 5 de noviembre de 2002.
Aceptado: 10 de marzo de 2003.
Abstract
We propose a new species Ceratium balechii sp. nov., a bloom-producing dinoflagellate in the Mexican tropical Pacific, and often by mistake referred to as C. dens. In order to know its morphological variability, a morphometric study was perfomed, on the basis of 584 cells from 48 populations collected in 11 coastal localities of the Mexican tropical Pacific. Sixteen morphological characters (14 quantitative and 2 qualitative) were studied in each cell, and another 6 characters, relevant for establishing allometric relationships, were taken into account. To perform the statistical analysis, four different cell shapes were distinguished based on their appearance. A discriminant analysis distinguished two forms that were also correlated with water temperature and salinity. Two forms are proposed, C. balechii f. balechii and C. balechii f. longum. The latter occurred in water with temperature higher than 26ºC and salinity less than 32. The species is subjected to cyclomorphosis and intense synchronized autotomy in both antapical horns. The new species is compared with other morphologically similar species and intraspecific taxa (C. dens, C. dens var. reflexa, C. tripos "var. benguela", C. tripos var. dalmaticum, C. tripos var. ponticum, C. californiense, C. ehrenbergii and C. porrectum). The presence of C. dens in the Mexican Pacific is heavily questioned.
Key words: Dinophyceae, Ceratium, new species, morphometry, tropical Pacific.
Resumen
Se describe el dinoflagelado Ceratium balechii sp. nov., taxón que comúnmente produce florecimientos en el Pacífico tropical Mexicano y que ha sido nombrado erróneamente como C. dens. Se realizó un análisis morfométrico para conocer la variación de la especie, incluyendo datos de 584 organismos pertenecientes a 48 poblaciones recolectadas en 11 localidades costeras del Pacífico tropical Mexicano. En cada organismo se midieron 16 caracteres morfológicos (14 cuantitativos y 2 cualitativos), y se consideraron otros 6 caracteres con significado alométrico. Para realizar el análisis estadístico los organismos fueron clasificados en cuatro grupos por su apariencia. Basándose en un análisis discriminante se reconocieron dos formas, cuya ocurrencia se correlacionó con la temperatura y salinidad del agua. Todo ello permitió describir dos formas taxonómicas: C. balechii f. balechii y C. balechii f. longum. Esta última predominando en agua con temperatura mayor de 26°C y salinidades menores a 32. Se compara a la nueva especie con taxones morfológicamente similares (C. dens, C. dens var. reflexa, C. tripos "var. benguela", C. tripos var. dalmaticum, C. tripos var. ponticum, C. californiense, C. ehrenbergii y C. porrectum). Se descarta la ocurrencia de C. dens en el Pacífico Mexicano.
Palabras clave: Dinophyceae, Ceratium, especie nueva, morfometría, Pacífico tropical.
Introduction
The genus Ceratium Schrank is one of the most important marine phytoplankton components, including around 80 "species" not considering other 30 dubious species (Sournia, 1986). For more than a century, Ceratium thecal tabulation has been known as a constant character (Kofoid, 1907a) with the formula 4', 5", 4-5c, 2+s, 5"', 2"" (Steidinger and Tangen, 1996), noting that all marine species have 4 cingular plates, whereas freshwater species have 5 (Sournia, 1986). That explains why thecal tabulation was ignored by taxonomists. In contrast, cell shape and size (mainly length and transdiameter), horn size (especially anthapical), and thecal ornamentation, have been used to distinguish between species. Other allomeric relations have been considered, such as the ratio between the lengths of the antapical horns (left and right), cell total length divided by the transdiameter (Steidinger and Tangen, 1996), and the ratio between the transdiameter and the length of the antapical horns (López, 1966). Based on the same morphometric characters, Ostenfeld (1903) and Jörgensen (1911) divided the genus into four subgenera, and their classification has been conserved over time.
This paper is dedicated to one species known in Latin American literature as Ceratium dens Ostenfeld et Schmidt (Balech, 1988; Cortés-Altamirano and Luna-Soria, 1998; Pech-Pacheco et al., 1999), C. tripos var. ponticum Jörgensen (Licea et al., 1995) or C. divaricatum Kofoid (Hernández-Becerril, 1989; in Cortes-Altamirano and Núñez-Pasten, 2000). It has been reported from the Gulf of California to Acapulco and Zihuatanejo in Guerrero, Mexico (Cortés-Altamirano and Hernández-Becerril, 1998), and was also cited as a bloom-producing species in the coastal waters of Mazatlán (Cortés-Altamirano and Núñez-Pasten, 2000). C. dens is reported as a part of the coastal planktonic flora of Ecuador (http://www.inocar.mil.ec/boletin/ebiologia.html) and from the coastal waters of Chile (Muñoz and Avaria, 1980). Trying to understand why different names have been assigned to organisms with practically the same morphology, we decided to search for the species in question retrospectively, and we understood that it was a confusion derived from Balech's (1988) monograph. C. dens described from the material from the Indian Ocean, has so peculiar morphological characters that Jörgensen (1911) opened a particular section (Densa section) to which he ascribed the species. The figures of "C. dens" from the SW Atlantic (neritic waters from southern Brazil) and from the Pacific coast of California presented by Balech (1988) are very similar to the specimens that are abundant in the Mexican Pacific and at the same time are different from the figures of C. dens by other authors (Jörgensen, 1911; Sournia, 1967; Taylor, 1976). Balech (1988) acknowledged the differences but he erroneously considered that other authors had drawn them inverted, because in C. dens the antapical left horn is the shortest one.
With the revision we made it was confirmed that only Wood (1954) had published an inverted image of the species.
C. dens was also considered a stage in the life cycle of C. reflexum Cleve (Konovalova, 1988). On the other hand, the authors who worked in the Mexican Pacific have contributed to the confusion by referring to it as C. tripos var. ponticum (Cortés-Altamirano and Hernández-Becerril, 1998; Licea et al., 1995), a species that, in our opinion, resembles a little the taxon in question. Steemann Nielsen (1934), López (1955, 1966), Dowidar (1972) and Yarranton (1967) carried out morphometric studies to clarify the taxonomy of Ceratium species and its varieties, because of the ample morphological variation that is known to occur in several Ceratium species. Seasonal variations in cell shape, maximum cell diameter and length of the horns (especially antapical) have been studied in some marine species, morphological variation being often correlated with temperature (Karsten, 1907; Steemann Nielsen, 1934; López, 1966).
The objective of the present study was to answer the following questions:
- Does C. dens really inhabit the Mexican Pacific?
- Is the taxon from the Mexican Pacific, referred to as C. dens or C. tripos var. ponticum, a new species?
- Which is the range of the morphological variation of the species in question?
- Could Ceratium with short antapical horns be an intraspecific taxon of C. tripos?
- Are the populations of the studied Ceratium species with short horns represent a stage or stages in the life cycle of a known species?
Material and methods
To answer the previously posed questions an exhaustive revision of the old literature was made in relation with C. dens and C. tripos, as well as a morphometric study of the species. Using a Carl Zeiss conventional microscope, we reviewed the material form 53 samples from 18 different localities of the Mexican tropical Pacific, collected from November 1998 to April 2001 (Fig. 1). This material corresponded to liquid samples collected with a net and fixed with formaldehyde, from the collection of the Laboratory of Marine and Brackishwater Phytoplankton, Universidad Autónoma Metropolitana - Iztapalapa. The specimens were digitally photographed and drawings were made with the aid of a camera lucida. Each sample was considered a population, from which several specimens were studied in detail. When the species was scarce, several water mounts were scanned, and all the cells were analyzed up to a maximum of 55, when the species was abundant.
Study area. The study was carried out in the Mexican tropical Pacific that corresponds to the 40% of the Mexican Pacific. Several authors agree that this region starts in Cabo Corrientes, Jalisco (De la Lanza, 1991; Espinoza-Ávalos, 1993; Serviere-Zaragoza et al., 1993). The current pattern is very variable in the study region, because the anticyclonic gyres do not penetrate to the coastal zone (Wyrtky, 1965).
Within the water column, the superficial layer is characterized by a temperature practically constant throughout the year and a permanent and shallow thermocline (between the 10 m and 30 m depths). Despite the warm waters of the Mexican tropical Pacific are characterized by low density, the intense evaporation increases the superficial salinity and with it the density, resulting in convection that causes a stable mixed superficial layer. In September, the winds hold great humidity producing hurricanes in the region. Besides, because in this region the continental shelf is very narrow, the coastal region is affected by particular local processes, such as river discharges and wind patterns. In general, it is known that the Mexican tropical Pacific waters are oligotrophic, but Thomas and Dodson (1972) mention that the nitrates are a limiting factor for phytoplankton development in this region.
In contrast with the subtropical-temperate part of the Mexican Pacific, where the diatoms dominate the phytoplankton (Meave del Castillo et al., 2003), in the Mexican tropical Pacific, especially during the dry season, dinoflagellates are dominant.
Evaluating the morphological characters. In each specimen, 16 morphological characters were measured, 14 quantitative and 2 qualitative (Table 1, Fig. 2). In the tips of the right and left antapical horns three character states were considered: a) pointed, b) truncated and, c) rounded. To evaluate the characters we followed the schemes of measurements used by Jörgensen (1911), Nie (1936), Graham and Bronikovsky (1944), López (1966), Muñoz and Avaria (1980) and Figueroa (1990). We present the following measurements for the first time: the angles of the body outline, the angle of the direction of the horns (apical and antapical), the length of the cell body (without the horns) and the length of the apical horn.

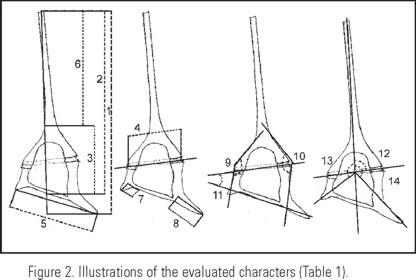
From the characters described above, 6 new quantitative characters were derived, that corresponded to ratios (Table 1, characters with numbers 17-22). It is worth mentioning that the total length (TL) of the cell, was obtained by tracing an axis parallel to the apical horn, considering its curvature and that it crossed the cingulum midpoint in dorsal or ventral view, forming a 90° angle with another axis that touched the lower point of the antapical horn. That way TL is affected by the length of the left apical and antapical horns as well as their direction.
Classification of cell shapes. The observed variation in "C. dens" specimens, all characterized by reduced antapical horns, especially the right one, was classified in four shapes, or morphotypes (Fig. 3a-p), named as follows:
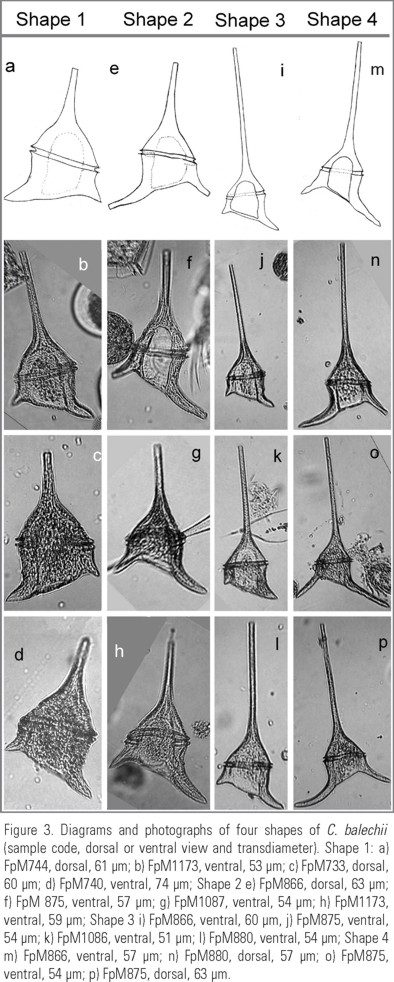
1. Short cell with short horns (Fig. 3a-d)
2. Short cell with long horns (Fig. 3e-h)
3. Long cell with short horns (Fig. 3i-l)
4. Long cell with long horns (Fig. 3m-p)
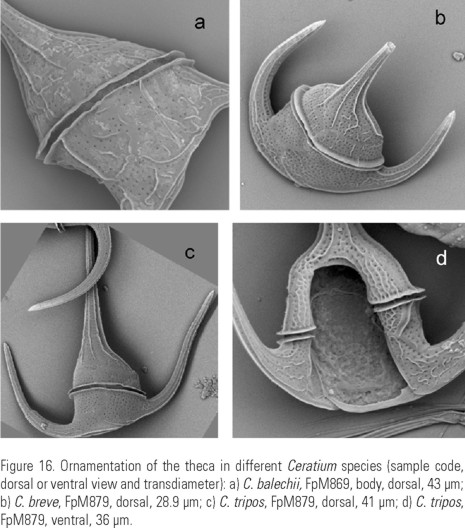
Also, to evaluate the intraspecific variation in the morphometric analysis, the measurements of 25 specimens of C. breve (Ostenfeld et Schmidt) Jörgensen were included as shape 5 (Fig. 16b).
Morphometric analysis. The data were analyzed in a matrix that included 619 specimens from 53 populations (including 5 of C. breve) of 11 localities. Table 2 presents the general information (frequency) of the samples (collection named FpM) that included populations used for the morphometric analysis. Only few of the 48 populations with "C. dens" had more than 20 individuals. Because of that and with the aim of having equivalent units in the number of specimens, all populations collected at different stations in the same sampling area were grouped. As a result only 14 groups of populations were considered in the analysis (table 2). Supposedly, this procedure did not affect the analysis, given the passive transport of organisms within a sampling area.
The 22 characters evaluated were subject to an exploratory analysis to recognize general patterns of dispersion and distribution as well as to detect possible outliers. Boxplots (including the median, quartiles and extreme values) for each character were made to classified them by cell shape and by population (Emerson and Strenio, 1983; Salgado-Ugarte, 1992).
Smoothed histograms (kernel density estimators) were used to observe in detail the values' distribution and were applied to all the observations to recognize multimodality, where each mode represents a potential shape (Scott, 1992; Salgado-Ugarte et al., 1993; Salgado-Ugarte, 2002).
To combine the smoothed histograms and the box diagrams, violin plots were used (Hintze and Nelson, 1998). These diagrams suggested the statistic comparison of shapes. This comparison was carried out using ANOVA (ANalysis Of VAriance) with one-way classification using the shape as a factor. In case of rejecting the hypothesis of homocedasticity (Bartlett or Levene criteria), non-parametrical Kruskal-Wallis tests were applied (Zar, 1999).
To verify the relationships between the characters, matrix scatterplots were made accompanied in each case by a correlation matrix (Salgado-Ugarte, 1992), without using the data of C. breve. The correlation was accepted when the test was significant (p <0.05).
To evaluate the differences between the characteristics of the different forms of "C. dens", a discriminant step-by-step forward analysis was performed, analyzing λ Wilks and considering the quantitative variables in its entity, although using only the individuals with complete data, and performing the analysis with 560 specimens.
Additionally, using the data on the relative abundance, the geographic distribution of the cell shapes was analyzed. To evaluate if the shapes correspond to genetic varieties or change along with the environment (pleiomorphism), temperature and salinity superficial values from the 10 m depth were plotted against the shapes, using only data on the cells collected with horizontal haules. In this analysis, short body cells (1+2) were compared against long cells (3+4), as well as cells with short antapical horns (1+3) against cells with long horns (2+4).
Results
Morphometry. Only 6 characters turned out to be constant in "C. dens": Body Length (BL), Transdiameter (T), Angle of the Apical Horn (AAH) Angle of the Right Antapical Horn (RHA), Angle of the Left Antapical Horn (LHA), and the ratio between the transdiameter and the body length (T/BL). The Violin plots showed in Fig. 6 that in spite of the morphological variation of "C. dens", the pattern of this variation was similar within the species and different from C. breve.
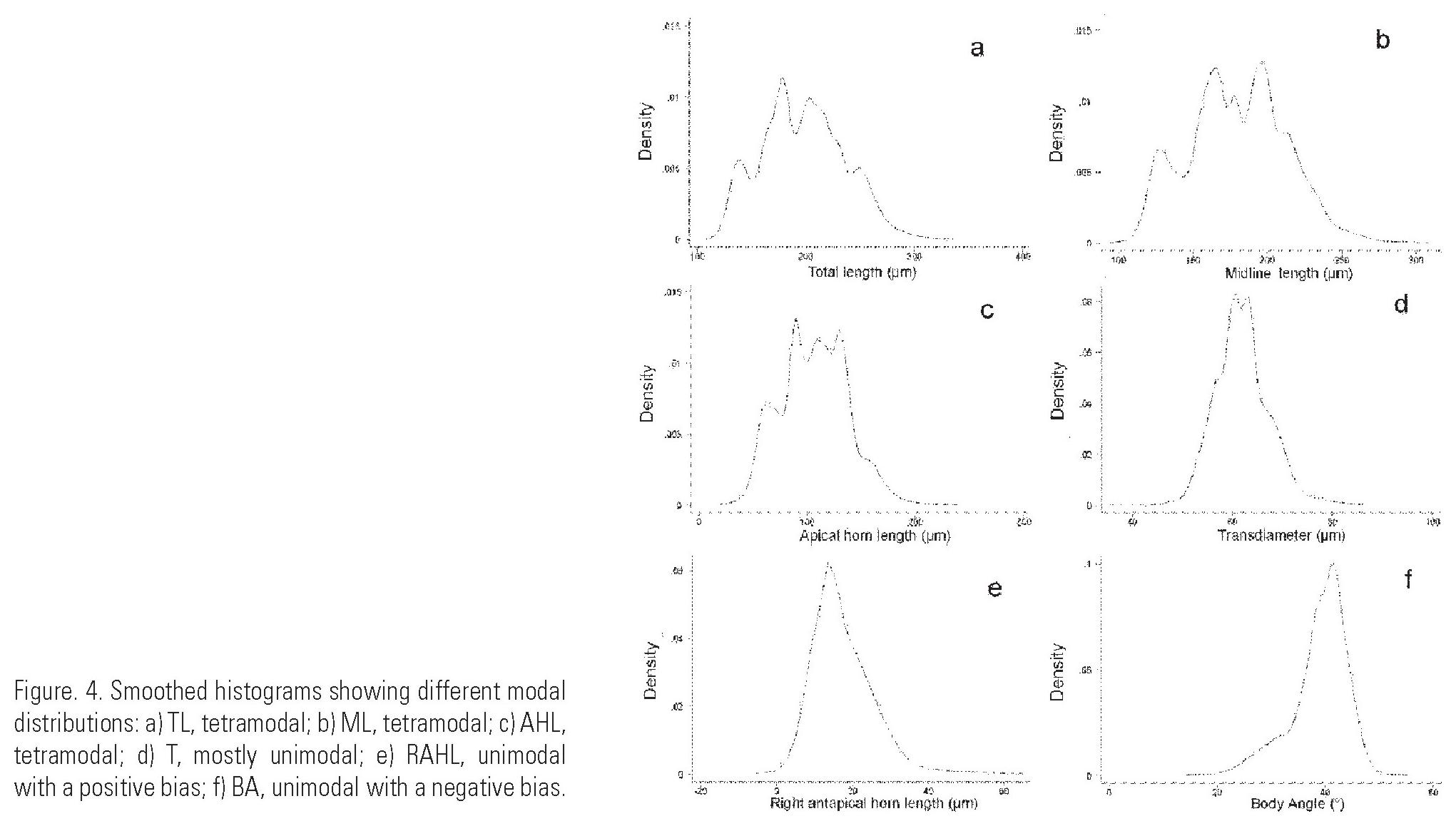
Statistical description of the characters. Several of the statistical plots showed long tails (Fig. 4a-f), which probably means that they are extraordinary cases. Characters had different distributions: Total Length (TL), Midline Length (ML) and Apical Horn Length (AHL) had a multimodal behavior (tetra- or pentamodal, Fig. 4a-c) that may correspond to the shapes. T, BL, Distance Between Antapical Horns (DH), Left Antapical Horn Length (LAHL) and LHA had a predominantly unimodal behavior (Fig. 4d). The rest of the characters, the Right Antapical Horn Length (RAHL), Angle of the Right Outline (RO), Angle of the Left Outline (LO), Angle at the distal part of the body (BA), AAH and RHA had an unimodal behavior biased to the right or to the left (Fig. 4e, f). For RAHL the bias may show a group of organisms with longer horns that do not have enough numbers to form a separate mode. Although is not shown in LAHL plot, individuals with longer horns are more frequent and are represented as an accesorial mode. The distribution of most of the angles was deviated towards values below the mode.
Characters' correlations. Several morphological characters are correlated in "C. dens", some as closely as TL and AHL (r=0.94, Fig. 5a) as well as the length of the antapical horns (r=0.92, Fig. 5c) and the distance between the antapical horns (DH) and the length of both horns LAHL (r=0.82, Fig. 5c) and RAHL (r=0.78, Fig. 5c). T did not correlate significantly with any of the horn lengths but it did with BL (r=0.52, Fig. 5b). AHL had a slight but significant correlation with both antapical horns (LAHL r=0.35, RAHL R=0.25). The right and left outline correlated slightly but significantly with one another (r=0.21). LO and RO were weakly but significantly correlated with BA (r=0.09) and no correlation was found between the LO and LAHL, and between RO and RAHL.
Statistical comparison of the cell shapes. Only the character TL was different for all shapes as showed in the boxplots inside the violin diagrams (Fig. 6a). Other characters distributed the shapes in two groups, i.e. LM and AHL separated shapes 1 and 2 vs. 3 and 4 (Fig. 6b, f, c, d). In contrast, the shapes 1 and 3 were grouped by the characters DAH and RAHL and LAHL (shapes with short horns vs. shapes with long horns, Fig. 6e, g, h.) Eight characters had a constant value independently on which shapes were evaluated: BL, T, RO, LO, AAH, BA, LHA and RHA. This fact was very important in assigning the species status to the taxon. Furthermore, 5 of those constant characters (BL, T, AAH, RA and LHA; Fig. 6a-n) differentiated C. breve from "C. dens".
Comparison of the median. Chi square was used to compare TL, ML, AHL and T. All shapes had a significant difference in total length (Bonferroni, p<0.01) which could mean that all four shapes are different taxa. In contrast, the comparison that used ML and AHL, grouped together the shapes with short cell vs. long cell (Bonferroni, p<0.05). Non-parametric statistics were used when comparing the shapes using RAHL, LAHL, DAH, RO, LO and AAH. When comparing the shapes using RAHL, there were significant differences between shapes with short antapical horns vs. long antapical horns. To discriminate between the three hypotheses posed by the results of the statistical analysis, a discriminant analysis was carried out, as well as analyzing the frequency, geographical distribution and the relationship of the shapes with environmental parameters (temperature and salinity).
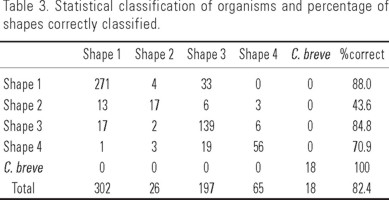
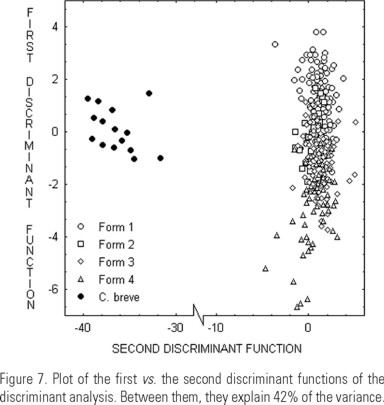
Discriminant analysis. The discriminant characters were RAHL, RHA, LHA, TL, DAH, LAHL and T. The specimens assigned to each of the five shapes were correctly placed within a range of 44-100%. Table 3 shows that it is easier to confuse shape 1 with shape 2, and shape 3 with shape 4. All shapes of "C. dens" are distributed in a gradient with shapes 1 and 4 in the extremes (Fig. 7). Both results were important when evaluating the taxonomical status of the species.
Geographical distribution and the species relationships with environmental factors. The most frequent organisms were those with short horns (Table 3/3). Based on the results of the discriminant and frequency analysis, shapes 1 and 2 were combined together and named short cell body; shapes 3 and 4 together were named long cell body. Figure 8 shows that long cells are more frequent in the southern Mexican tropical Pacific, from Acapulco to Huatulco.
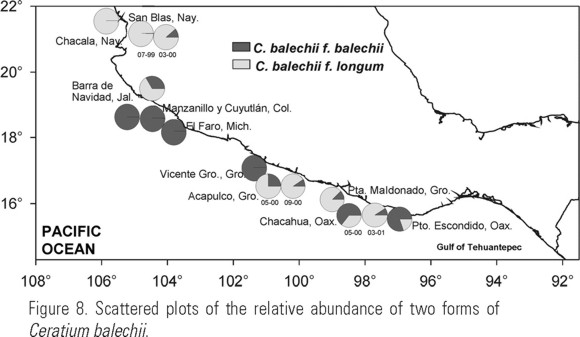
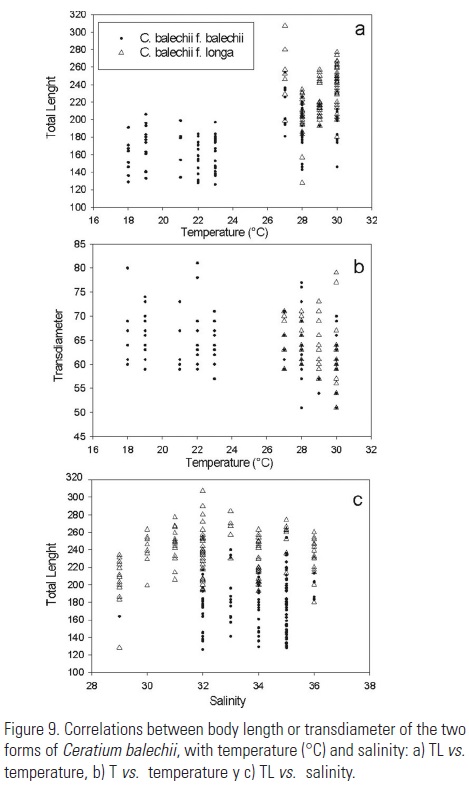
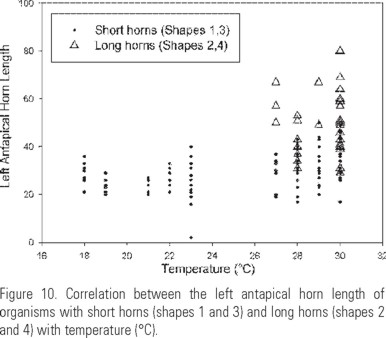
Long cells were found in places with temperature above 26ºC with a salinity below 32 (Fig. 9a, b) while short cells predominated below 24ºC with a salinity above 32 (Fig. 9c). The distribution of long left antapical horn organisms was correlated with temperatures above 26ºC (Fig. 10).
Taxonomy
Ceratium balechii Meave, Okolodkov et Zamudio sp. nov.
Ceratium dens auct. non Ostenfeld et Schmidt: Cortés-Altamirano and Hernández-Becerril, 1998: 68, fig. 23; Pech-Pacheco et al., 1999: fig. 1, 3a, 7; Cortés-Altamirano and Núñez-Pasten, 2000: 309, fig. 3; C. tripos var. ponticum auct. non Jörgensen: Licea et al., 1995: 49, pl. 5, fig. 6a- (b)c.
Diagnosis: Cellula quinquangularis asymmetrice, 125-254 µm longitudine; spina apicalis 38-145 µm longitudine; spinae antapicales duae, reductae, distinctae longitudine directioneque; spina dextra spina sinestra semper reductiora, recta et lateraliter-postice posita; spina sinestra recta vel leviter curvata, plerumque lateraliter, numquam antice extensa.
Type locality: Mexican Pacific, El Faro, Michoacán. Neritic sample, collected at 2 nm from the coast (18°19'58"N, 103°32'23"W), net sample, 54 µm mesh size, vertical haul, 70 m depth. Collection number: FpM744. Collecting date: April 9, 2000.
Collectors: Meave and Zamudio.
Habitat: Neritic.
Distribution: Widely distributed in the Mexican Pacific from the California Gulf to Puerto Escondido, Oaxaca. Bloom-forming species in Mazatlán, Sinaloa; Manzanillo, Colima; Cuyutlán, Colima; El Faro, Michoacán.
Etymology: Named after Dr. Enrique Balech, a prominent dinoflagellate researcher who closely described the new species (Balech, 1988).
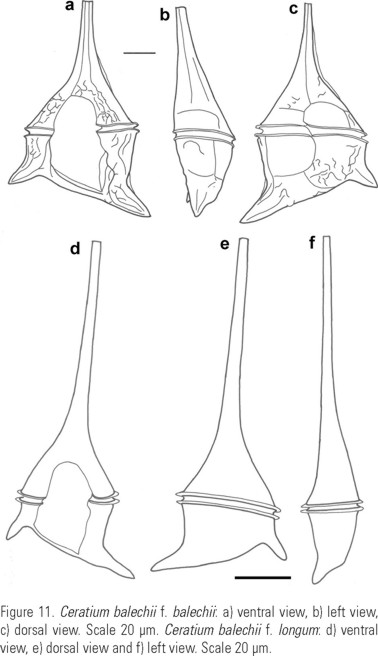
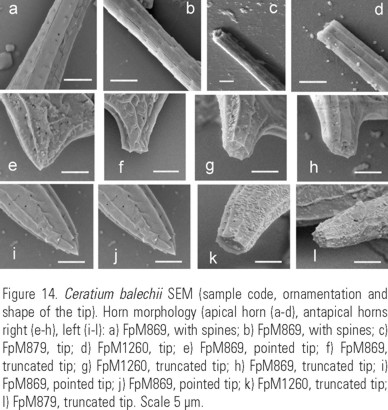
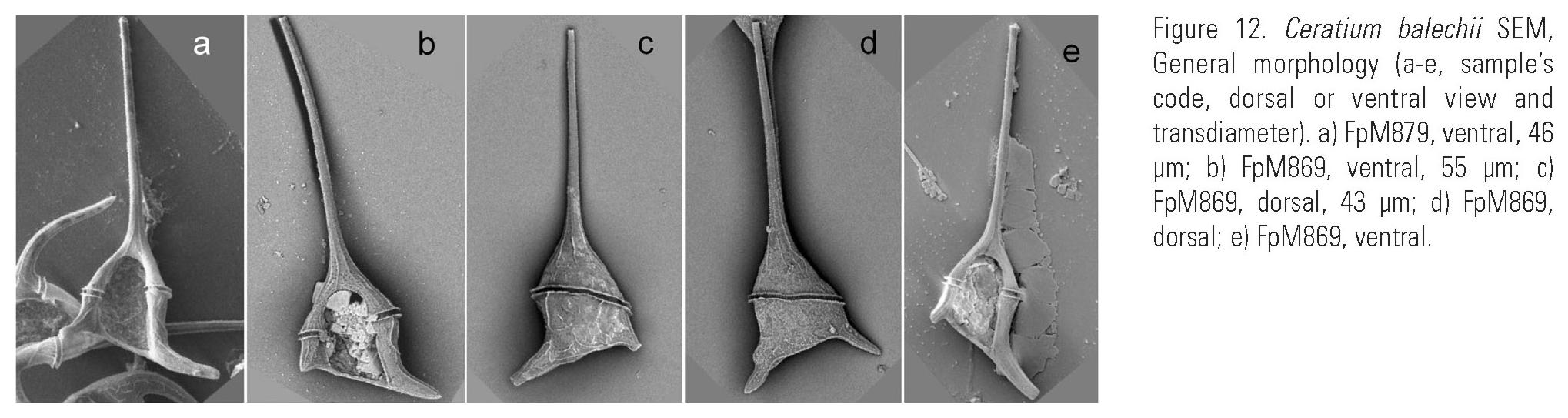
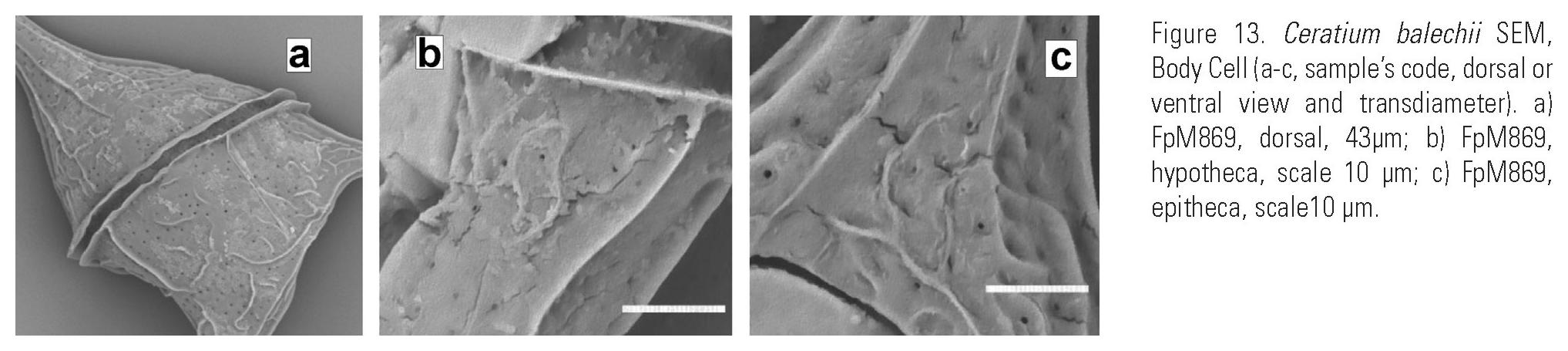
Description: Pentagonal asymmetric cell, with hypotheca tilted towards the left (Fig. 12a-b, (c, d) e). Apical horn truncated and open (Fig. 14c-d). Both antapical horns reduced and different in length and direction, with pointed tips (Fig. 14e, i-j) or truncated but always closed (Fig. 14f-h,k-l). The right antapical horn always shorter than the left one, straight and in a lateral-posterior direction (Fig. 11a, d; 12a-c). The left horn straight or slightly curved, usually more lateral and never directed towards the anterior part of the cell (Fig. 11a, b, c, d, e; 12b, e). All horns with equidistant ribs and small spines (Fig. 14a-l). The margins of the epitheca may be convex (Fig. 3c; 13a), straight (Fig. 3m-n) or the left one convex and the right one slightly concave (Fig. 12b). In ventral or dorsal view the margin of the hypotheca tilted towards the left horn. In dorsal view, the cingulum is almost perpendicular to the left horn (Fig. 11c, d, e; 12c-d; 13a). Punctate theca with spread pores (Fig. 13b-c; 16a).
Dimensions: Total length 125-254 µm average 180 µm. Length of the apical horn 38-145 µm, average 91 µm. Length of the body 50-127 µm, average 73 µm. Transdiameter 51-81 µm, average 63 µm. Length of the apical horn 38-145 µm, average 91 µm. Length of the left antapical horn 2-56 µm, average 33 µm. Length of the right antapical horn 1-40 µm, average 18 µm. Distance between the tips of the antapical horns 40-129 µm. Right outline angle 117°-163°, average 145°. Left outline angle 122°-156° average 140°. BA: 18°-51°, average 40°. LAHL/RAHL: 0.1 - 23, average 2.1; T/RAHL: 2-47, average 4.3.
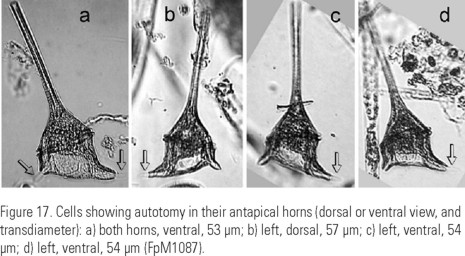
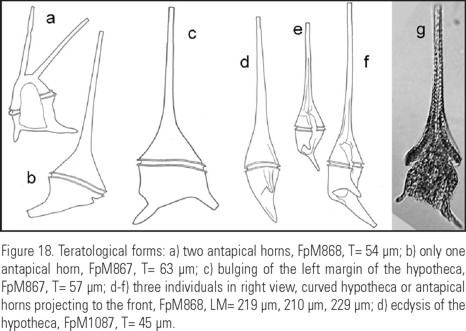
The species presents a marked autotomy, mostly in the antapical horns, the shapes with very reduced horns being more frequent, sometimes looking as stumps. The regeneration of the antapical horns may be evident. (Fig 17a-d).
Ceratium balechii f. longum Zamudio et Meave
Diagnosis: Forma cum characteribus speciei, sed longior (128-312 µm) et cum spinis apicalibus longior (95-217 um).
Type locality: Mexican Pacific, Acapulco, Guerrero Neritic sample collected at 6 nm from the coast (16°46.36'N, 99° 55.83'W), net sample, 54 µm mesh size, vertical haul, 50 m depth. Collection number: FpM866. Collecting date: May 19, 2000.
Collectors: Meave and Zamudio.
Description: The general morphology of the body, the theca outline and the characteristics of the antapical horns are similar to the type variety. Total length 174 - 312 µm, average 234 µm; with a longer apical horn: 57 to 217 µm, average 136 µm. The length of the left antapical horn varies from 20 to 80 µm, and the distance between the antapical horns reaches 160 µm, average 100 µm. LAHL/RAHL: 0.3-4.4, average: 3.5. T/RAHL: 0.4 -8.2, average 3.0. In specimens with a longer left antapical horn it is common to observe constrictions along its length (Fig. 3m).
Discussion and conclusions
Taxonomy of the species. The new species is similar to: C. dens (Fig. 15a), C. dens var. reflexa Schmidt (Fig. 15b), C. tripos "var. benguela" (Fig. 15d), C. californiense Kofoid (Fig. 15f) y C. ehrenbergii Kofoid (Fig. 15g).
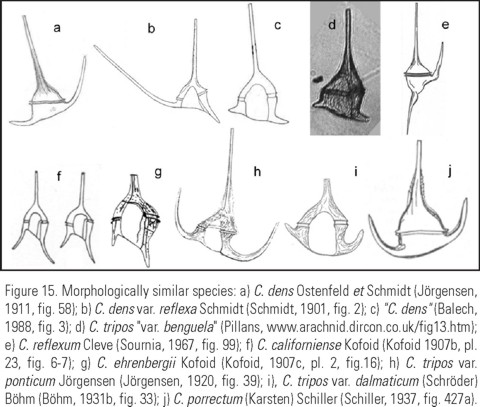
The original description of C. dens (Ostenfeld y Schmidt, 1901: 165-166; Fig. 16) says "body robust, left antapical horn very short, most commonly robust, straight or slightly curved, tooth-like". All these characters correspond the iconotypes that show specimens in dorsal view. The specimens described were collected in the Gulf of Aden, the Indian Ocean.
In later publications, different authors (Karsten, 1907; Jörgensen, 1911; Böhm, 1931a; Steemann Nielsen, 1934; Nie, 1936; Schiller, 1937; Wood, 1954; Sournia, 1967; Subrahmanyan, 1968), described and illustrated different specimens name as C. dens in agreement with the original description.
Schmidt (1901: 214, Fig. 2) described a new variety of C. dens var. reflexa, which in fact differs from the type variety only by the direction of the left antapical horn, bent downward. The validity of the var. reflexa is doubtful, because C. dens shows a wide variability in the direction of the left antapical horn (Böhm, 1931a: 15-16; Fig. 11a-(b, c, d), e).
Few authors described or drew C. dens differently. As it was mentioned in the introduction, Wood (1954: 284; Fig. 204) published an inverted image of a dorsal view of C. dens, although in his description he mentioned clearly that the left antapical horn is the shortest. Konovalova (1998: 139, 151-152; Pl. 29, Fig. 11) registered C. dens in the Sea of Japan, but her illustration looks as if she had used or copied a drawing made by Balech (1988: 291; Pl. 69, Fig. 5), that does not correspond at all to the description of C. dens, and pointed out that the specimen may have two kinds of antapical horns: 1) both antapical horns of the same length, with the right one slightly shorter, or 2) the right horn longer than the left one, in a ratio of 3.5:5 and with a pronounced anterior curvature. We believe that the first case is not C. dens.
It is worth noticing that in all the illustrations of C. dens that coincide with the original description, the cell body is more or less triangular with straight or slightly convex epithecal margins. Besides, the tips of the antapical horns are pointed or "closed". The straight or slightly convex hypothecal outline was considered characteristic of the species by Nie (1936), but is a feature common at least in C. tripos (Fig. 15c-d)y C. breve. (Fig. 15b).
It is interesting to notice that all authors who described their specimens, which corresponded to the original description of C. dens, were collected in the Indian Ocean (Ostenfeld and Schmidt, 1901; Karsten, 1907; Jörgensen, 1911; Steemann Nielsen, 1934, 1939; Wood, 1954; Sournia, 1967; Subrahmanyan, 1968) or in the West Pacific (Schmidt, 1901; Böhm, 1931a; Steemann Nielsen, 1934, 1939; Nie, 1936; Schiller, 1937; Wood, 1954). No specimens with the original characters of C. dens have been reported from the Atlantic Ocean.
C. californiense Kofoid (Kofoid, 1907b: 302-; Pl. 23, Fig. 6-9) and C. ehrenbergii Kofoid (Kofoid 1907c, pl. 2, Fig. 16), described from the tropical and subtropical Pacific (Kofoid, 1907b: 302; Pl. 23, Fig. 6-9 (7, 8)), have reduced antapical horns, especially the right one. But in both species, the antapical horns follow the same direction. The asymmetry in the direction of the antapical horns is considered here as a diagnostic character for C. balechii. Furthermore, both C. californiense and C. ehrenbergii differ from the new species in having a more rounded cell body and the hypothecal outline tilted towards the left antapical horn.
Balech (1988) designated specimens from the SW Atlantic under the name of C. dens. However, his specimens have the right antapical horn shorter than the left one (Fig. 15c), a feature that does not coincide with the original description of C. dens, but it is very similar to C. balechii. Noticing that the specimens presented variation in the length of the antapical horns, he completed the illustration using the material from the E Pacific (California coast) that presented longer horns. This resulted in the illustration of a cell with long symmetrical horns directed laterally that looks more like C. porrectum (Fig. 14j), (= C. divaricatum, according to Schiller, 1937).
Other publications that reported C. dens from the SW coasts of Africa and the Mexican Pacific, show illustrations very similar to C. balechii. Some specimens similar to C. balechii form the Mexican Pacific were designated as C. tripos "var. benguela" by D. Harbour, in a doctoral thesis, although it has not been formally published. This material was found in SW Africa both in net and bottle samples at a depth of 7 m, in areas with an intense upwelling of the Benguela Current, that seems to favor the development of the species (Julian Pillans, personal communication).
The illustrations by Cortés-Altamirano and Hernández-Becerril (1998), the photographs by Cortés-Altamirano and Núñez-Pasten (2000) and Pech-Pacheco et. al. (1999) look very similar to our material and different from C. dens.
Licea et. al. (1995) designated, described and illustrated very similar specimens in their atlas both as C. dens (Pl. 18, Fig. 2) and as C. tripos var. ponticum (Pl. 5, Fig. 6a- b, c), although their images and descriptions that do not correspond to Jörgensen's original description of C. tripos var. ponticum. Jörgensen (1920: 49, Fig. 38, 39) defines C. tripos var. ponticum (Fig. 15h) having a "squarrose right postical horn... and is often strongly bent in a ventral direction, and the widely spread posterior horns... the posterior horns moderately long, a little diverging at their tips, both in relation to each other and to the apical horn, which is usually rather long and often nearly parallel to the right posterior one". Böhm (1931b: 381, Fig. 34) pictures C. tripos var. ponticum, which has slender and rather long antapical horns, bent anteriorly. Clearly, this species has antapical horns more or less symmetrical and bent towards the apical horn. Why did Licea et. al. (1995) identify the Mexican specimens as C. tripos var. ponticum? Trying to find out where was the origin of the "ponticum" confusion, we found that López (1966: 332, Fig. 36) named an organism with short and with asymmetrical shape and direction of the tips of both antapical horns as C. tripos var. ponticum. Possibly, because this paper is cited in Licea et al.'s atlas, it could be one the sources of the confusion. To complicate matter, Cortés-Altamirano and Hernández-Becerril (1998: 68-69, Fig. 23a, b) considered in their description of C. dens, that C. tripos var. ponticum was its synonym. However, C. dens and C. tripos var. ponticum are completely different from each other and from C. balechii, too.
Other species similar to C. balechii is C. tripos var. dalmaticum (Fig. 15i), with antapical horns shorter and dissimilar (with the right one reduced), both strongly curved in a lateral or anterior-lateral direction.
Why do not we consider C. balechii as a new variety of C. tripos as some authors did it with the organisms found in Benguela current? To answer this question it is important to discuss the status of the taxonomy of C. tripos (Fig. 16c-d), described by Müller (1786). Pavillard (1931) wrote that delimiting C. tripos (O.F. Müller) Nitzsch is one of the most complex problems in the systematics of dinoflagellates. Sournia (1967) pointed out that C. tripos can be a subject of a separate monograph. After a thorough research about C. tripos, Balech (1988) concluded that it is the oldest species of the genus and it has a wide range, being distributed in both neritic and oceanic zones, including cold, temperate and warm waters, and presenting an extreme polymorphism. According to him, although there have been a large number of varieties, subspecies and forms named, all of them omit to name the type variety C. tripos var. tripos. Therefore, he assigned Figure 65, pl. IV, of Jörgensen (1911) as the iconotype of this variety, and composed the description of C. tripos var. tripos using the descriptions of the forms balticum, subsalsum and atlanticum. This is the description we used to base our decision about which variety should be considered as the type variety.
When deciding if our specimens corresponded or not to an infraespecific taxon of C. tripos, we considered important that C. balechii has a pentagonal body shape, while C. tripos var. tripos and all the rest of varieties and forms have a trapezoidal asymmetrical shape.
On the basis of the analysis of our material, we excluded the possibility that the specimens from the Mexican Pacific could be seasonal forms or autotomized cells from other species already known. There were no cells with the same characteristics of C. balechii with long antapical horns. The description of the new taxon does not correspond to any other species. Some cells from the Mexican Pacific, especially those collected around Acapulco, present one or two constrictions in the left antapical horn (Fig. 3m, o), which is indicative of the beginning of the autotomy, and of that the species really has reduced horns during its all life cycle.
Konovalova (1998) suggested that the specimens of C. dens might represent a stage on the C. reflexum life cycle (Fig. 15e) or other species closely related to it. C. balechii is not C. reflexum with broken or autotomized horns, because they have different directions. Besides, C. reflexum has a trapezoidal body shape, and C. balechii is pentagonal. C. reflexum was present in the samples of the Mexican Pacific, but was never abundant, that is why we discarded the possibility to consider the abundant specimens described as C. balechii as life-cycle stages of this species.
Taxonomy of the forms. When new species of Ceratium are identified or described, two phenomena must be taken into consideration: 1) the seasonal variation in morphology, when morphological changes happen very quickly, even within one generation life span, similar to the cyclomorphosis of zooplankton species , and 2) autotomy, i.e. a rapid alteration of the horn length by "self cutting" (Taylor, 1987). The term "autotomy" was first used by Kofoid (1908) and Lohmann (1908) and later by Peters (1934), and it has been observed in many marine species of Ceratium, especially in specimens with long horns belonging to the Macroceros and Amphiceratium sections.
Autotomy of the species. It is clear that C. balechii possesses a marked intrinsic autotomy, considering that it happens in both forms described (Fig. 17a-(b, c, d). Some results, such as smoothed histograms showing long tails, direct observations on horn regeneration and the more frequent occurrence of shapes 1 and 3 (with short horns) in comparison to shapes 2 and 4 (with long horns), show that in C. balechii autotomy happens quickly, in both the apical and antapical horns and is synchronized in the antapical horns. The synchronization is deduced from the fact that a high correlation between the antapical horns length was observed. Figure 18g shows a specimen with the hypotheca's ecdysis, a common phenomenon during Ceratium blooms with high cell density (Kofoid, 1908). In C. balechii, exuviation of the distal portions of the antapical horns and their regeneration also occur (Fig. 17a-d). The fact that the forms with long horns were found mainly in the southern Mexican tropical Pacific and were associated with temperatures higher than 26°C is in complete agreement with Kofoid's (1908) observations, who found that some species of Ceratium "self cutted" their horns when changing a habitat from temperate to cold water. In spite of tetramodal distribution of several characters in smoothed histograms, we did not consider the differences between the lengths of the horns to describe additional forms, because we assume that the long horns are labile.
Cyclomorphosis. Seasonal variation in body shape, the length and transdiameter and of the horns (especially the left antapical) have been studied in different species of Ceratium (Steemann Nielsen, 1934; Nordli, 1953; López, 1955, 1966; Böhm, 1976). Karsten (1907) was the first to observe that the Ceratium species with longer horns were present in the well-lit and warmer waters, suggesting that the length of the horns was an adaptive character. López (1966) confirmed that in warmer waters, the cells are thinner, with longer horns, altering their allometric ratios. In late summer and early fall, in Kattegat, Nielsen (1956) found that cells were shorter and with shorter antapical horns. These changes are well correlated with temperature. Steemann Neilsen (1934) also suggested that the temperature is the main factor in determining the shape of Ceratium cells, and he also considered the neritic or oceanic conditions to be important. Nordli's (1957) observations on cultivated dinoflagellates showed that shape changed with the amount of nutrients, water temperature and cell abundance.
In the Mexican tropical Pacific, C. balechii f. longum is dominant only in its southern part. At the beginning, it was suggested that it could be a (genetic) geographic variety, but the correlation found between the cell shapes and the water temperature allowed us to consider the characters pleiomorphic. Thus, we suggest that the geographic distribution of the forms is a result of the influence of the cold California Current in the northern part of the study area.
The temperature of the water affects its density. Taken this fact into account, López (1966) suggested that the shape of Ceratium cells may be directly affected not so much by the temperature but by the viscosity of the water. Our results agree with López' observations, because we also found a correlation between the dominant form and salinity (Fig. 9c), with shorter form (C. balechii f. balechii) dominating at salinities between 32 and 36.
Finally, we were able to recognize even teratological (aberrant) forms of C. balechii (Fig. 18a-(b, c) d), which is another argument in favor of being a well-defined species.
Conclusions
1. C. dens has not been found in the Mexican Pacific. The specimens named as C. dens collected from the Mexican Pacific have been misidentified.
2. In spite of the great morphological variation of C. balechii, it is discrete, which allowed us to distinguish intraspecific taxa.
3. C. balechii is a distinct species, because its antapical horns have different size and direction, compared with other species and intraspecific taxa, the right horn being shorter.
4. In C. balechii, three almost constant characters, the cell body length, the angle of direction of the apical horn and the angle of direction of the right antapical horn suggest that all the shapes found correspond to only one species.
5. The statistical differences in the total length, the length of the apical horn and the transdiameter, support the separation of C. balechii f. longum. This form may be considered a cyclomorphic stage; it dominates in warmer waters (both in warmer waters at a station and in warmer seasons) and lower salinities. Probably, this form is an adaptation to lower water densities.
6. C. balechii is a species with an intense autotomy that is synchronized in the antapical horns.
7. The specimens that closer resemble C. balechii are those illustrated by Balech (1988, Pl. 69, Fig. 3-4) as C. dens and those photographed and placed into the electronic report (http://www.inocar.mil.ec/boletin/ebiologia.html) and named informally as C. tripos "var. benguela" by Harbour. In both cases, the images differ from C. balechii in the direction of the antapical horns.
8. C. balechii is different from C. dens in that its right antapical horn is te shortest instead of the left one, as in C. dens.
9. Direction of the antapical horns in C. balechii, discard the possibility of its specimens belonging to C. porrectum (reported from the Mexican Pacific as its synonym C. divaricatum, according to Schiller, 1937).
10. C. balechii does not correspond to a life cycle stage, or to a teratologic form, or to autotomized specimens of other species.
11. C. balechii does not belong to C. tripos, because its body shape is pentagonal, while the latter has trapezoidal body shape.
12. C. balechii seems to be circumscribed to the Mexican Pacific, because, probably, neither Balech's specimens from the SW Atlantic, nor C. tripos "var. benguela" from the NW coast of Africa, clearly belong to C. balechii. There are some reports without images of "C. dens" in the coastal waters of Ecuador (http://www.inocar.mil.ec/boletin/ebiologia.html), Peru (Patricia Villanueva-Merino, personal communication) and the northern coasts of Chile (Muñoz and Avaria, 1980). If these reports confirm the presence of C. balechii in these regions, its distribution will include the East Equatorial and subtropical Pacific.
Aknowledgments
To Sandra Guerra Martínez of the UAM-I for some measurements of dinoflagellate cells and technical assistance. To Mona Hoppenrath at the Helgoland Biological Station of the Alfred Wegener Institute for Polar and Marine Research in Germany and Eduardo Morales at the Patrick Center for Environmental Research of the Philadelphia Academy of Natural Sciences for sending us literature from the late 19th and early 20th centuries. To Julian Pillans at the University of Newcastle-upon-Tyne, U.K., for information on material collected in the Benguela Current. To Lizeth Botes at the University of Cape Town, South Africa, for a photograph of "C. dens" from the coast of South Africa. To Richard Moe at the University Herbarium of the University of California at Berkeley, for the Latin translations of the diagnoses. To Patricia Castillo for the SEM photographs. To Eréndira Julieta Cohen-Fernández for improving the style and her critical comments. To Jesús Sánchez for his suggestion to use the discriminant analysis. This study was partially funded with a grant from CONABIO (Project S-151 "Diatomeas Planctónicas del Pacífico Tropical", 1998-2001) and by CONACyT (Project G37560-V "Biología y Filogenia Molecular de Dinoflagelados del Pacífico Mexicano", 2002-2006), and the support for the work of Yuri Okolodkov in the Universidad Autónoma Metropolitana - Iztapalapa (Cátedra Patrimonial de Excelencia CONACyT, 2001-2003).
References
BALECH, E. 1988. Los dinoflagelados del Atlántico Sudoccidental. Publ. Espec. Inst. Esp. Oceanogr. 1. Ministerio de Agricultura, Pesca y Alimentación, Madrid. 219 p., 88 pl. [ Links ]
BÖHM, A. VON. 1931a. Distribution and variability of Ceratium in the Northern and Western Pacific. Bernice P. Bishop Museum Bulletin, 87: 1-46, pl. 1. [ Links ]
BÖHM, A VON. 1931b. Die adriatischen Ceratien (Nach dem Materiale der österreichischen Adriaforschung in den Jahren 1911-1914). Botanische Archiv 31: 349-385. [ Links ]
BÖHM, A. VON. 1976. Morphologische Studien im Bereiche der Pyrrhophyta. Das Problem Form und Selektion. Bibliotheca Phycologica, 22. J. Cramer, Vaduz. 199 p. [ Links ]
CORTÉS-ALTAMIRANO, R. and D. U. HERNÁNDEZ-BECERRIL. 1998. Especies responsables y probables de mareas rojas. In: R. CORTÉS-ALTAMIRANO R. (Ed.). Las mareas rojas. ATG editor S.A., D.F., México, pp. 43-80. [ Links ]
CORTÉS-ALTAMIRANO, R. and R. LUNA-SORIA. 1998. Lista mundial de microalgas responsables de florecimientos, mareas rojas y tóxicas. In: CORTÉS A.R. (Ed.). Las mareas rojas. ATG editor S.A., D.F., México, pp. 141-153. [ Links ]
CORTÉS-ALTAMIRANO, R. and A. NÚÑEZ-PASTEN. 2000. Distribución y abundancia anual de Ceratium dens (Peridiniales: Ceratiaceae) en el Golfo de California, México. Revista de Biología Tropical 48 (2-3): 305-311. [ Links ]
DE LA LANZA, G. E. 1991. Oceanografía de los mares mexicanos. AGT Editor, México. 569 p. [ Links ]
DOWIDAR, N. M. 1972. Morphological variation in Ceratium egyptiacum in different habitats. Marine Biology 16: 138-149. [ Links ]
EMERSON, J. D. and J. STRENIO. 1983. Boxplots and batch comparison. In: D.C. HOAGLIN, F. MOSTELLER and J. W. TUKEY (Eds.) Understanding robust and exploratory data analysis. John Wiley & Sons, NY, pp. 58-96. [ Links ]
ESPINOZA-ÁVALOS, J. 1993. Macroalagas marinas del Golfo de California. In: S. I. SALAZAR-VALLEJO and N. E. GONZÁLEZ (Eds.) Biodiversidad marina y costera de México. CONABIO-CIQROO. Chetumal. 875 p. [ Links ]
FIGUEROA, M. G. 1990. Sistemática y distribución del género Ceratium Schrank 1793, en el sur del Golfo de México (Feb.-Dic. 1987). Tesis de Maestria. Facultad de Ciencias, UNAM, México. 51 p. [ Links ]
GRAHAM, H. W. and N. BRONIKOVSKY. 1944. The genus Ceratium in the Pacific and North Atlantic oceans. Scient. Results. Cruise VII. Carnegie Inst. Wash. Pub. 564 (5): 1-209. [ Links ]
HINTZE, J. L. and R. D. NELSON. 1998. Violin plots: a box plot-density trace synergism. The American Statistician 52 (2): 181-184. [ Links ]
JÖRGENSEN, E. VON. 1911. Die Ceratien. Eine kurze Monographie der Gattung Ceratium Schrank. Internationale Revue der gesamten Hydrobiologie und Hydrographie, 4. Biologische Supplement - Heft 1. 124 p., 10 Taf., 184 Fig. [ Links ]
JÖRGENSEN, E. VON. 1920. Mediterranean Ceratia. Report on the Danish Oceanographical Expeditions 1908-10 to the Mediterranean and Adjacent Seas. Vol. 2. Biology. J. 1: 1-110. [ Links ]
KARSTEN, G. 1907. Das Indische Phytoplankton. Nach dem Material der Deutschen Tiesee-Expedition 1898-1899. Deutsche Tiefsee-Expedition 1898-1899, 2: 1-328, Taf. 35-54. [ Links ]
KOFOID, C. A. 1907a. The plates of Ceratium with a note on the unity of the genus. Zool. Anz., 32, 7: 177-183. [ Links ]
KOFOID, C. A. 1907b. Dinoflagellata of the San Diego region, III. Descriptions of new species. Contributions from the Laboratory of the Marine Biological Association of San Diego. XII. University of California Publications, 3, 13: 299-340, pl. 22-33. [ Links ]
KOFOID, C. A. 1907c. New species of dinoflagellates. Reports of the Scientific Results of the Expedition to the Eastern Tropical Pacific from October, 1904, to March, 1905. IX. Bulletin of the Museum of Comparative Zoology at Harvard College, 50, 6: 163-207, pl. 1-18. [ Links ]
KOFOID, C. A. 1908. Exuviation, autotomy and regeneration in Ceratium. University of California Publications in Zoology, 4 (7): 387-393. [ Links ]
KONOVALOVA, G. V. 1998. Dinoflagellatae (Dinophyta) of the Far Eastern seas of Russia and adjacent waters of the Pacific Ocean. Dalnauka, Vladivostok. 300 p. (In Russian). [ Links ]
LICEA-DURAN, S., J. L. MORENO, H. SANTOYO and M. G. FIGUEROA. 1995. Dinoflageladas del Golfo de California. UBCS, SEP-FOMEX, México, 165 p. [ Links ]
LOHMANN, H., 1908. Untersuchungen zur Feststellung des vollstän digen Gehaltes des Meers an Plankton. Wissenschafen Meeresunters Ab. Kiel 10: 129-370. [ Links ]
LÓPEZ, J. 1955. Variación alométrica en Ceratium tripos. Investigación Pesquera, Barcelona, 2: 131-159. [ Links ]
LÓPEZ, J. 1966. Variación y regulación de la forma en el género Ceratium. Investigación Pesquera, Barcelona, 30: 325-427. [ Links ]
MEAVE DEL CASTILLO, M. E., M. E. ZAMUDIO RESÉNDIZ, J. A. AKÉ-CASTILLO, S. GUERRA-MARTÍNEZ and I. F. BARBOSA-LEDESMA. (2003). Biodiversidad de Diatomeas (Bacillariophyta) en la Columna de Agua del Pacífico Mexicano. In: BARREIRO, T., M. E. MEAVE DEL CASTILLO, G. FIGUEROA-TORRES and M. SIGNORET (Eds.). Planctología Mexicana. Sociedad Mexicana de Planctología, A.C. (SOMPAC), pp. 43-84. [ Links ]
MÜLLER, O. F. 1786. Animalcula infusoria fluviatilia et marina, que detexit, systematice descripsit et ad vivum delineari curavit. Opus hoc posthumum quod cum tabulis aeneis L. in lucem tradit vidua ejus nobilissima, cura Othonis Fabricii. Hauniae. 367 p., 50 pl. [ Links ]
MUÑOZ, S. P. and S. AVARIA. 1980. Estudio taxonómico de los dinoflagelados tecados de la Bahía de Valparaíso. I. Género Ceratium. Revista de Biología Marina 17: 1-43. 12 pl. [ Links ]
NIE, D. 1936. Dinoflagellata of the Hainan region. I. Ceratium. Contributions from the Biological Laboratory of the Science Society of China, Nanking, Zoological Series, 12, 3: 29-73. [ Links ]
NIELSEN, J. 1956. Temporary variations in certain marine Ceratia. Oikos, 7: 256-272. [ Links ]
NORDLI, E. 1953. Salinity and temperature as controlling factors for distribution and mass occurrence of Ceratia. Blyttia 11: 16-18. [ Links ]
OSTENFELD C. H. 1903. Phytoplankton from the sea around the Faroes. Botany of the Faroes, Copenhague 2: 558-611. [ Links ]
OSTENFELD C. H. and J. SCHMIDT. 1901. Plankton fra det Rode Hav og Adenbugten (Plankton from the Red Sea and the Gulf of Aden). Vidensk. Meddi. Dansk. Naturh. Foren. 25: 161-182. [ Links ]
PECH-PACHECO, J. L., J. A. ALVAREZ-BORREGO, E. ORELLANA-CEPEDA and R. CORTÉS-ALTAMIRANO. 1999. Diffraction pattern applicability in the identification of Ceratium species. Journal of Plankton Research 21 (8): 1455-1474. [ Links ]
PAVILLARD, J., 1931. Phytoplancton (Diatomées, Péridiniens) provenant des campagnes scientifiques du Prince Albert Ier de Monaco. Résultats de la Campagne Scientifique de Prince Albert I, 82: 208 pp., 3 pl. [ Links ]
PETERS, N. 1934. Die Bevölkerung des Südalantischen Ozeans mit Ceratien. Wissensch. Erg. Deut. Atl. Exp. Meteor 1925-1927, 11: 1-69. [ Links ]
SALGADO-UGARTE, I. H. 1992. El análisis exploratorio de datos biológicos. Marc Ediciones. México. 243 p. [ Links ]
SALGADO-UGARTE, I. H. 2002. Suavización no paramétrica para análisis de datos. FES Zaragoza, UNAM, México, 139 p. [ Links ]
SALGADO-UGARTE, I. H., M. SHIMIZU and T. TANIUCHI. 1993. Exploring the shape of univariate data using kernel density estimators. Stata Technical Bulletin 16: 8-19. [ Links ]
SCHILLER, J. 1937. Dinoflagellatae (Peridineae). In: Dr. L. Rabenhorst's Kryptogamen-Flora von Deutschland, Österreich und der Schweiz, 3, 2. Akademische Verlagsgesellschaft M.B.H. 590 p. [ Links ]
SCHMIDT, J. 1901. Preliminary report of the botanical results of the Danish Expedition to Siam (1899-1900). Part IV. Peridiniales. In: J. SCHMIDT. Flora of Koh Chang. [ Links ] Botanische Tidsskrift 24: 212-221. [ Links ]
SCOTT, D. W. 1992. Multivariate Density Estimation: Theory, Practice, and Visualitation. John Wiley & Sons, New York. [ Links ]
SERVIERE-ZARAGOZA, E., J. GONZÁLEZ-GONZÁLEZ and D. RODRÍGUEZ-VARGAS. 1993. Ficoflora de la región de Bahía de Banderas, Jalisco-Nayarit. In: S. I. SALAZAR-VALLEJO and N. E. GONZÁLEZ (Eds.) Biodiversidad marina y costera de México. CONABIO-CIQROO. Chetumal. 875 p. [ Links ]
SOURNIA, A. 1967. Le genre Ceratium (péridinien planctonique) dans le canal de Mozambique. Contribution a une révision mondiale. Vie et Milieu. Ser. A: Biologie marine, 18, 2-3A: 375-500. [ Links ]
SOURNIA, A. 1986. Atlas du phytoplancton marin. Vol. 1: Introduction, Cyanophycées, Dictyochophycées, Dinophycées et Raphidophycées. Editions du Centre National de la Recherche Scientifique, Paris. 219 p. [ Links ]
STEEMANN NIELSEN, E. 1934. Untersuchungen über die Verbreitung, Biologie und Variation der Ceratien im südlichen Stillen Ozean. The Carlsberg Foundation's Oceanographical Expedition round the world 1928-30 and previous "Dana"-Expeditions. Dana-Report, 4. Copenhagen, London. 67 p. [ Links ]
STEEMANN NIELSEN, E. 1939. Die Ceratien des Indischen Ozeans und der ostasiatischen Gewässer mit einer allgemeinen zusammenfassung über die verbreitung der Ceratien in den Weltmeeren. The Carlsberg Foundation's Oceanographical Expedition round the world 1928-30 and previous "Dana"-Expeditions. Dana-Report 17. Copenhagen, London. 33 p. [ Links ]
STEIDINGER, K. A. and TANGEN K. 1996. Dinoflagellates. In: C. R. TOMAS (Ed.). Identifying marine diatoms and dinoflagellates. Academic Press, Inc., Harcourt Brace & Company, San Diego et al.: 387-598. [ Links ]
SUBRAHMANYAN, R. 1968. The Dinophyceae of the Indian seas. I. Genus Ceratium Schrank. Marine Biological Association of India, Mandapam Camp. Memoir II. 334 p., 79 pl. [ Links ]
TAYLOR, F. J. R. 1976. Dinoflagellates from the International Indian Ocean Expedition. A report on material collected by the R.V. "Anton Bruun" 1963-1964. E. Schweizerbart'sche Verlagsbuchhandlung (Nägele u. Obermiller), Sttutgart. 234 p., 46 pl. [ Links ]
TAYLOR, F. J. R. 1987. Dinoflagellate morphology. In: F. J. R. TAYLOR (Ed.). The biology of dinoflagellates. Botanical Monographs, 21. Blackwell Scientific Publications, Oxford et al.: 24-91. [ Links ]
THOMAS, W. H. and A. N. DODSON. 1972. On the nitrogen deficiency in tropical Pacific oceanic phytoplankton II. Photosynthetic and cellular characteristics of a chemostat-grown diatom. Limnology and Oceanography 17 (4): 515-523. [ Links ]
WOOD, E. J. F. 1954. Dinoflagellates in the Australian region. Australian Journal of Marine and Freshwater Research 5: 171-351. [ Links ]
WYRTKY, K. 1965. Surface current on the Eastern Tropical Pacific Ocean. Bulletin Inter-American Tropical Tuna Commission. Vol IX (5): 269-304. [ Links ]
YARRANTON, G. A. 1967. Parameters for use in distinguishing populations of Euceratium Gran. Bulletin of Marine Ecology 6: 147-158. [ Links ]
ZAR, J. H. 1999. Biostatistical analysis. 4th ed. Prentice May, Upper Saddle River. 591 p. [ Links ]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}