











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El propósito de este artículo es evidenciar las potencialidades que posee el modelado geoespacial en el fortalecimiento del Análisis de Riesgo de Plagas (ARP) en diferentes escalas geográficas, visualizado como una estrategia técnico-científica que apoya la toma de decisiones ante agentes patógenos que ponen en riesgo la seguridad agroalimentaria. De manera específica, el riesgo fitosanitario que representa el hongo Fusarium oxysporum f. sp. cubense Raza 4 Tropical (FocR4T), recientemente clasificado como Fusarium odoratissimum (Maryani, Lombard, Kema y Crous, 2019) considerado una amenaza latente para la producción de plátano y banano en las zonas productoras del mundo.
A nivel mundial, las plagas y enfermedades agrícolas son responsables de pérdidas entre 20 y 40% de la productividad de los cultivos alimenticios (Bouwmeester et al., 2016). Específicamente, FocR4T puede afectar platanares de América pantropical, que en términos de producción alimentaria se consideran el cuarto cultivo más importante del mundo después del arroz, el trigo y el maíz, y de importancia económica de algunos países en desarrollo (Molina et al., 2008).
FocR4T es una enfermedad cuarentenaria, considerada letal por ocasionar daños irreversibles para el cultivo (Dita et al., 2018), puede provocar afectaciones cercanas al 40%; incluso logró devastar plantaciones de plátano en diferentes países de Asia, Medio Oriente, África y Oceanía (García-Bastidas et al., 2015). La primera detección en América representa un desafío para producción platanera de Latinoamérica y el Caribe, donde se cultiva el 36.4% de la producción mundial, con exportaciones de 14.5 millones de toneladas, equivalente al 66.2% del producto interno bruto de la economía regional. La probabilidad de establecimiento, dispersión y potencial de daño económico (PDE) por Fusariosis de las musáceas es alto, con posibles afectaciones en 1.3 millones de hectáreas y daños directos a 12.3 millones de personas que viven del cultivo (Dita et al., 2018).
La modelación geoespacial da el soporte a la toma de decisiones estratégicas en agricultura, ya que permite analizar de manera conjunta los requerimientos óptimos para estimar la probabilidad relativa en la invasión de plagas agrícolas (Donatelli et al., 2017; Tuomola et al., 2018). Aquí un elemento clave se considera el movimiento internacional de mercancías que posibilita el incremento en la dispersión de riesgos fitosanitarios entre regiones geográficas (Dyer et al., 2017), incluso se ha empleado como indicador en la determinación de la dispersión del riesgo fitosanitario (Thangavelu et al. 2020).
En 2016 se estimó una superficie de plátano de poco más de 20 millones de hectáreas distribuidas en 142 países, con una producción estimada de 289 mil toneladas, y la América pantropical aportó el 8.46% de la producción mundial (OIRSA, 2019). Esta información puede ser empleada para la estimación del riesgo fitosanitario causado por la humanidad mediante el movimiento de productos agrícolas a nivel global, denominado PDE.
El objetivo de este trabajo fue caracterizar las áreas potenciales bajo algún nivel de riesgo fitosanitario de Fusarium oxysporum, f. sp. cubense raza 4 tropical (RF_FocR4T), para identificar posibles escenarios de introducción y establecimiento del patógeno a escala global con énfasis en América pantropical, lo cual se sustentó en los objetivos específicos: 1) modelar el nicho ecológico del patógeno/hospedante; 2) estimar el PDE, y de manera conjunta, 3) estimar el RF_FocR4T a través de la MCDA en diferentes escalas.
Materiales y métodos
La aptitud climática del patógeno/hospedante (ACP y ACH, respectivamente) se estimó a través del modelo de máxima entropía que incorpora los datos de presencia (Phillips et al., 2006). Su fundamento parte de un vector z de componentes {f_j} donde j=1,…,n son los valores correspondientes a las variables ambientales y x(z) es un elemento con condiciones ambientales z en la ubicación x. La probabilidad de solo presencia Pr(y=1 | x(z)) toma la forma de una regresión logística con la interceptada estimada por la entropía de qλ, en función de los valores de las variables ambientales z en una determinada ubicación x (Phillips y Dudik, 2008):
Donde qλ es el estimador de máxima entropía de la distribución de Pr(x (z)| y=1) dado por la expresión siguiente:
En complemento, λ= (λl,...,λn) es un vector de ponderaciones de la j-ésima variable ambiental f_j. Mientras que Z_λ es una constante de normalización que asegura que las probabilidades q_λ sumen 1 en el área de estudio. H es la entropía de la función de densidad q_λ y se define por:
Se emplearon 54 detecciones del patógeno expresadas en coordenadas geográficas estandarizadas a la proyección Plate Carrée (latitud, longitud), Datum WGS84. Tales localizaciones fueron tomadas de artículos científicos correspondientes a tres regiones del mundo descritas en el Cuadro 1. Como complemento se identificaron 61 localizaciones de hospedantes de importancia económica distribuidas en América pantropical (Cuadro 2).
Cuadro 1 Sitios de presencia de FocR4T empleados en la modelación de la ACP (54).
| Detecciones | Área geográfica | Región | Fuente |
| 1 | Isla Mayotte | África | Aguayo et al., (2020) |
| 3 | Uttar Pradesh, Kushi Nagar y Ambedkar Nagar en India. | Asia | Damodaran et al., (2019) |
| 34 | Kalimatan, Borneo, Java, Aceh, Sumatra, Lampug, Papua Nueva Guinea, Salawesi y Tenggara | Sudeste Asiático | Maryani et al., (2019) |
| 2 | Llanura costera del Carmelo y Kibbutz Ein Gev, orilla oriental del lago de Galilea en Israel | Medio Oriente | Maymon et al., (2018) |
| 13 | Laos, Myanmar, Vietnam y Yunan | Sudeste Asiático | Zheng et al., (2018) |
| 1 | Valle del Jordán | Medio Oriente | García-Bastidas et al., (2015) |
Cuadro 2 Sitios de presencia del hospedante empleados en la modelación de la ACH (61).
| Detecciones | Área geográfica | Fuente |
| 15 | Zona de producción de banano en Colombia, Venezuela y Brasil | Corbi et al., 2013 (7 sitios) y De la Pava et al., 2020 (8 sitios). |
| 8 | Zona de la producción de banano en Norte y Centro América. | Smith et al., 2010 (1 sitio), Ramos-Hernández et al., 2011 (1 sitio), Arendt et al., 2012 (5 sitios), Rivera Rivas et al., 2015 (1 sitio), Sol-Sánchez et al., 2016 (1 sitio). |
| 38 | Sitios de producción de banano del país con mayor riesgo conforme al modelo de PDE. | Elbehri et al., 2015 (34 sitios), Cedeño-García et al., 2016 (4 sitios). |
La caracterización ambiental se realizó mediante la interacción con 19 variables macro ambientales de la base global de WorldClim versión 2.1 (Fick e Hijmans, 2017) (Cuadro 3), lo que permitió establecer límites ambientales para una posible propagación futura (Pitt et al., 2009). El porcentaje del tamaño de la muestra de validación (PTMV) se calculó con la fórmula propuesta por Schaafsma y Van Vark (1979), PTMV = [1+(p-1)1/2]-1, donde p es el total de variables explicatoria (19), lo que determinó un valor PTMV de 0.19.
Cuadro 3 Variables bioclimáticas empleadas en la modelación de nicho ecológico.
| Grupo | Variable | Descripción | Unidad de medida |
| Temperatura | BIO1 BIO2 BIO3 BIO4 BIO5 BIO6 BIO7 BIO8 BIO9 BIO10 BIO11 |
Temperatura media anual Rango diurno medio Isotermalidad (BIO2/BIO7)*(100) Estacionalidad de la temperatura (desviación estándar * 100) Temperatura máxima del mes más cálido Temperatura mínima del mes más frío Rango anual de temperatura (BIO5-BIO6) Temperatura media del trimestre más húmedo Temperatura media del trimestre más seco Temperatura media del trimestre más cálido Temperatura media del trimestre más frío |
° C ° C Adimensional % ° C ° C ° C ° C ° C ° C ° C |
| Precipitación | BIO12 BIO13 BIO14 BIO15 BIO16 BIO17 BIO18 BIO19 |
Precipitación anual Precipitación del mes más húmedo Precipitación del mes más seco Estacionalidad de la precipitación (coeficiente de variación) Precipitación del trimestre más húmedo Precipitación del trimestre más seco Precipitación del trimestre más cálido Precipitación del trimestre más frio |
Mm mm mm % mm mm mm mm |
El modelo se implementó mediante el uso del software MaxEnt v3.3.3e. El procesamiento se inició con el uso una semilla aleatoria, un PTMV de 19%, y un número de 10 000 “background points” (Lira-Noriega et al., 2018). MaxEnt se ejecutó de forma iterativa y generó 10 modelos de validación, con un modelo final promedio (West et al., 2016). El potencial predictivo del modelo se evaluó mediante el análisis del área bajo la curva de la característica operativa relativa (AUC, por sus siglas en inglés) (Fielding y Bell, 1997).
La modelación geoespacial del PDE involucró: 1) identificar países con producción de plátano del periodo de 2013 a 2017, conforme la fracción arancelaria 0803 (CCI, 2016); 2) intersectar con la base de datos global de EPPO (2020), conforme con la situación fitosanitaria del patógeno; 3) determinar los flujos de comercialización entre nodos (origen vs destino); 4) construir una base de datos espacial para dimensionar la producción de material vegetativo, y con ello, 5) crear mapas globales de la trazabilidad espacial de material vegetativo como posible fuente de inóculo.
El planteamiento para la determinación del PDE se sustentó bajo el principio de que una probable zona fuente de inóculo, representada por los nodos de origen, respecto a países con los que mantienen relaciones comerciales, ya que estas interacciones pueden favorecer la dispersión hacia una nueva área o territorio (FAO, 1995).
El RF_FocR4T en América pantropical se estimó mediante la asociación espacial de la ACP, la ACH y el PDE. Esta metodología involucró un proceso de estandarización mediante un procedimiento de membresía difusa para homogenizar a un mismo rango “0.0 a 1.0” (Cuadro 4). Posteriormente, se combinaron los criterios asignándoles la misma importancia a través de una fórmula de combinación lineal ponderada, para así obtener una estimación del RF_FocR4T, el cual fue reclasificado en cinco rangos: muy bajo (< 0.15), bajo (0.15-0.30), medio (0.30-0.50), alto (0.50-0.70), muy alto (>0.70).
Cuadro 4 Criterios empleados en la MCDA del RF_FocR4T.
| No | Criterio | Descripción | Unidades | Rango | Membresía difusa |
| 1 | ACP | Ambiente favorable para la ocurrencia de FocR4T | adimensional | 0-1 |
 (CLP: 0.0-1.0) |
| 2 | ACH | Ambiente favorable para el establecimiento del hospedante | adimensional | 0-1 | |
| 3 | PDE | Flujos de material vegetativo | ton/año | 31-3,353,600 |
Para evaluar la relación estadística existente entre las variables explicativas y el RF_FocR4T, se empleó la Correlación de Pearson para determinar el signo y magnitud de la tendencia entre los valores estimados de RF_FocR4T y las variables bioclimáticas con mayor valor de importancia que determinaron el modelo resultante.
Resultados y discusión
La ACP determinó que existen condiciones ambientales que favorecen al patógeno en cuatro regiones a nivel global: 1) Sureste de Asia (región insular); 2) Asia meridional; 3) África Central y Madagascar) y 4) Sudamérica, con porciones diversas en Centro y Norteamérica (Figura 1, superior).
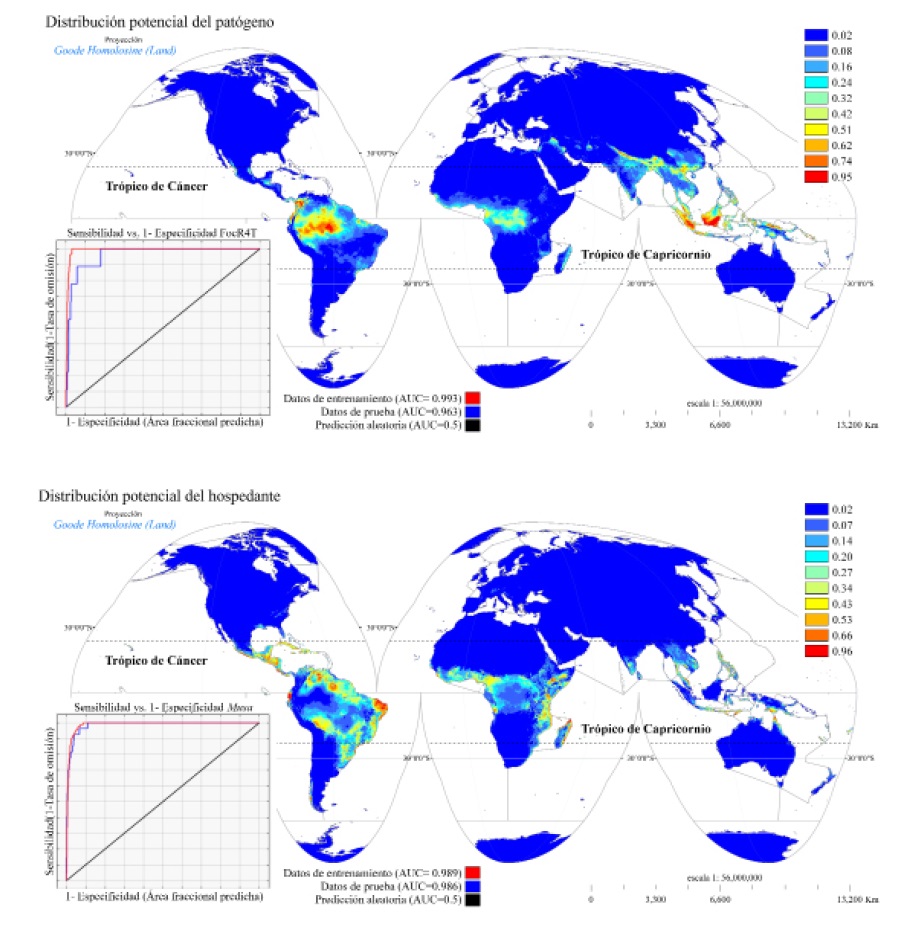
Figura 1 Modelo de Máxima Entropía. Aptitud climática del patógeno (superior) y Aptitud climática del hospedante (inferior).
El grado de contribución a la variabilidad espacial de la posible distribución del patógeno se logró definir por las variables de precipitación del trimestre más húmedo (BIO16, 33%) y estacionalidad de la temperatura (BIO4, 29.3%). Mientras que la contribución que tendría la variable explicatoria si se empleara sola (importancia de permutación), fue liderada por la temperatura media anual (BIO1) y la temperatura media del trimestre más seco (BIO9) con 59.1 % y 11.6 %, respectivamente. Se obtuvo un buen desempeño predictivo con un área bajo la curva de 0.975 (Cuadro 5).
Cuadro 5 Contribución porcentual e importancia de permutación de las variables empleadas en la modelación de Máxima Entropía del nicho ecológico del patógeno y hospedante.
| Grupo | Variable | ACP (FocR4T) | ACH (Plátano o Banano) | ||
| Contribución porcentual (%) |
Importancia de permutación (%) |
Contribución porcentual (%) |
Importancia de permutación (%) |
||
| Temperatura | BIO1 BIO2 BIO3 BIO4 BIO5 BIO6 BIO7 BIO8 BIO9 BIO10 BIO11 |
4.4 0.3 0.1 29.3 - 11.3 1 0.1 3.4 0.1 0.4 |
59.1 5 - 1.7 - 0.7 0.1 - 11.6 2.6 1.6 |
3.7 6.2 6.4 15.7 2.7 5.7 38.6 0.6 - 4.7 3.3 |
1.1 1.4 1.4 63.8 0.2 0.2 0.2 5.9 - 0.5 1.2 |
| Precipitación | BIO12 BIO13 BIO14 BIO15 BIO16 BIO17 BIO18 BIO19 |
- - 2.7 2.5 33 - 10.3 1.2 |
- - 0.1 7.8 0.2 - 6.8 2.6 |
- 0.3 4.5 5.3 1.7 0.1 0.2 0.4 |
- 3 8.8 9.2 0.8 0.7 0.6 1.1 |
Como complemento, los lugares que presentan condiciones ambientales que favorecen la ACH (plátano) se distribuyen en el centro norte de Sudamérica, en países como Brasil, Perú, Ecuador y Colombia (Figura 1, inferior). La contribución porcentual se definió por el rango anual de temperatura (BIO7, 38.6%) y la estacionalidad de la temperatura (BIO4, 15.7%). Sin embargo, la importancia de permutación fue por la estacionalidad de la temperatura (BIO4), con un valor de 63.8%, lo cual se respaldó con un valor COR: 0.978 (Cuadro 5).
Los países que presentan riesgo antrópico por PDE se distribuyen en tres continentes (América, Asia y África). La dinámica de la movilización de los productos del género Musa spp durante el periodo de estudio fue de 5 289 427 toneladas hacia 35 países. El PDE se agrupó en: a) los grandes exportadores como China (87.7%), Hong Kong (6.4 %), Jordania (4.0%) y Malasia (1.7%) y, b) los pequeños exportadores. que incluyeron Pakistán, Australia, Mozambique y Filipinas. que en conjunto aportaron menos del 1% del total (Figura 2)
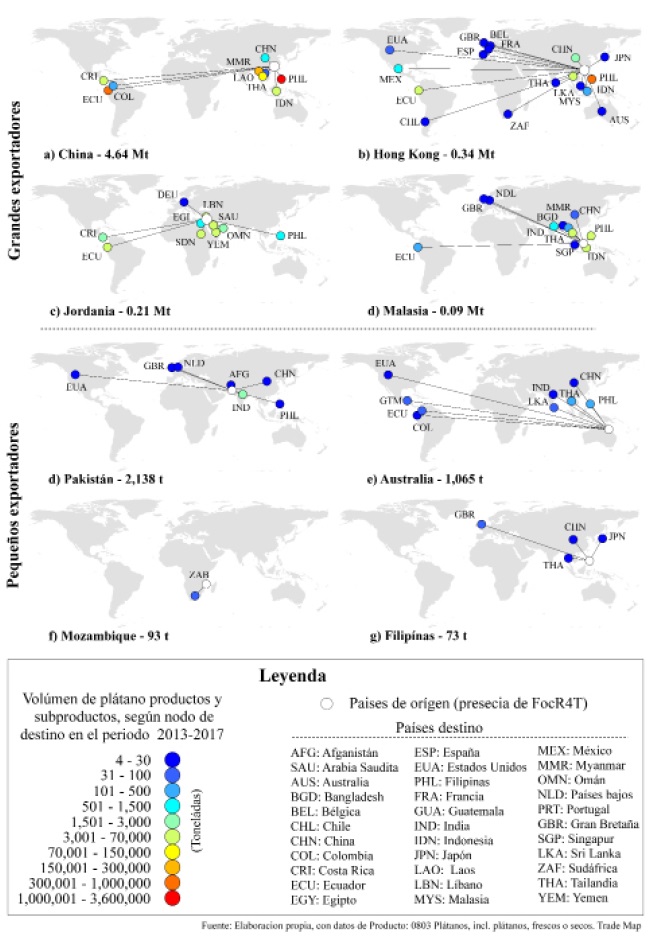
El mayor flujo por volumen se destacó en seis nodos destino: Filipinas (67.1%), Ecuador (17.9%), Myanmar (5.1%), Tailandia (2.9%), Vietnam (2.2%), Sudán (1.31%). Para el caso del continente americano, los nodos destino con mayor volumen fueron Ecuador (17.9%), Costa Rica (0.4%), México (<0.1%, 1,082 t), Colombia (<0.1%, 473 t), Estados Unidos (<0.1%, 98 t), Guatemala (<0.1%, 92 t) y Chile (<0.1%, 29 t) (Figura 2).
A nivel global, se identificaron cuatro áreas focales con diferentes niveles de RF_FocR4T. Cabe señalar que dentro de ellas se identificó una superficie prioritaria de 29 567 km2 en riesgo fitosanitario muy alto (> 0.9). Geográficamente se distribuyen en territorios que abarcan cuatro grandes extensiones integradas de la siguiente forma: a) en el Sudeste Asiático, una extensión de 2366.4 km2 (8.015%), distribuidos en Indonesia, Tailandia, Camboya, Malasia, Filipinas, Timor Leste y el norte de Australia; b) el Sureste de África, con una superficie de 434.06 km2 (1.46 %), distribuida en Madagascar, Mozambique, Malawi y Kenia; c) en África Occidental 7.56 km2 (0.025 %), en la región de Liberia y d) la región pantropical de América, de mayor extensión, con 26 758.9 km2 (90.5%), que se distribuye en Perú, Guyana, Ecuador, México, Panamá, Venezuela, Colombia, Guatemala y Brasil (Figura 3).
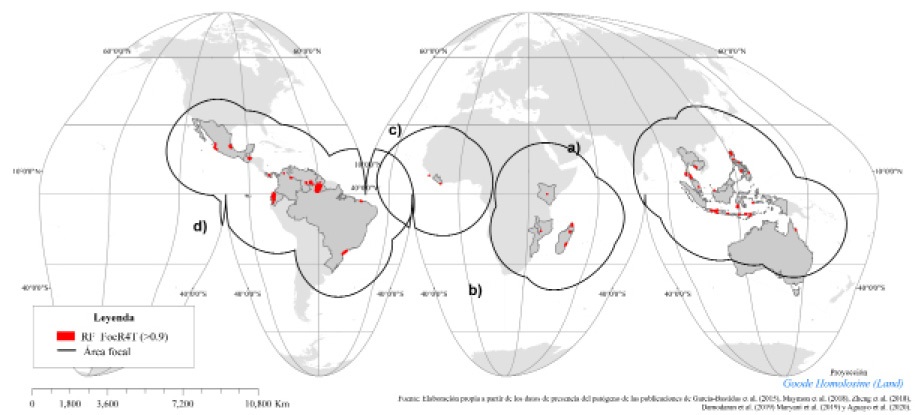
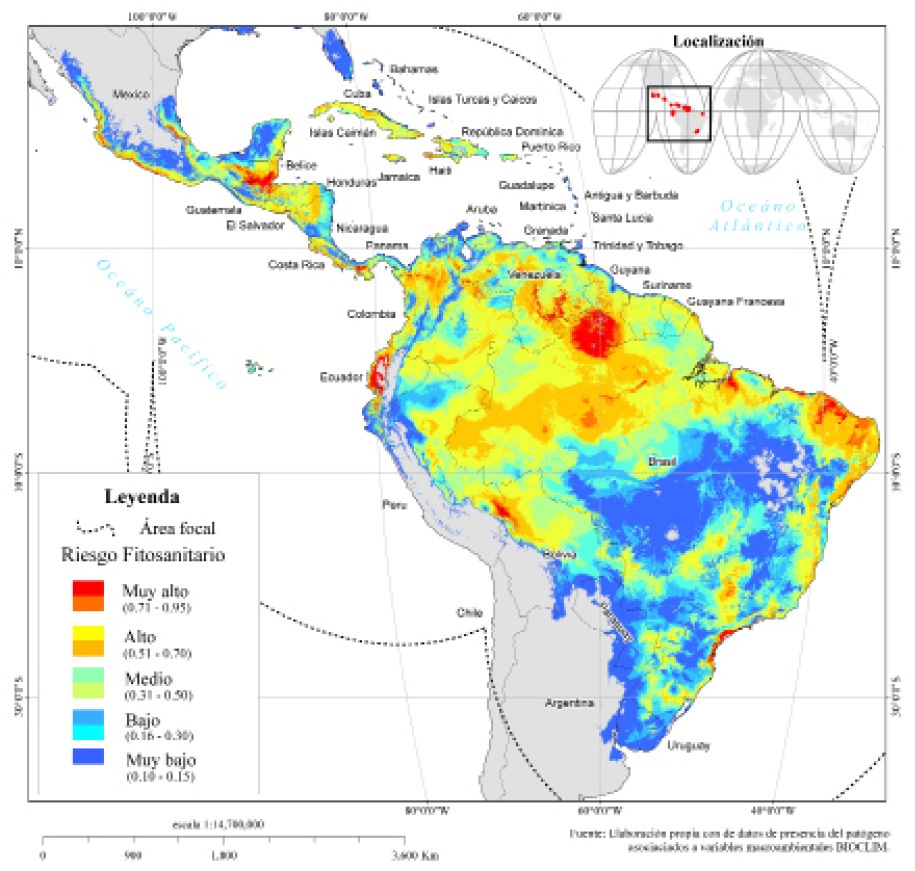
A escala subcontinental, en la región pantropical de América (Figura 4) se identificaron 15.8 millones de km2 con algún nivel de RF_FocR4T, lo cual se refleja en el Cuadro 6, donde se puede observar la superficie estimada por nivel de RF_FocR4T en los 45 países identificados.
Cuadro 6 Niveles de RF_FocR4T localizados en América pantropical.
| Región | País | Porcentaje del área total identificada por país y a nivel de RF_FocR4T (%) | Área total por país según nivel de RF_FocR4T (km2) |
Porcentaje del área identificada en América pantropical (%) |
||||||||
| <0.2 | 0.3 | 0.4 | 0.5 | 0.6 | 0.7 | 0.8 | 0.9 | <1 | ||||
| Norte | México | 41.23 | 11.58 | 11.18 | 9.38 | 15.52 | 7.57 | 2.17 | 1.78 | 0.51 | 806 547.62 | 5.09 |
| Estados Unidos | 88.30 | 6.20 | 2.74 | 1.18 | 1.54 | 0.05 | 262 842.37 | 1.66 | ||||
| Subtotal | 1 069 389.996.75 | 6.75 | ||||||||||
| Centro | Nicaragua | 12.74 | 17.14 | 20.72 | 11.63 | 15.72 | 21.73 | 0.31 | 113 151.46 | 0.84 | ||
| Honduras | 2.5 | 7.37 | 11.87 | 7.51 | 42.24 | 27.98 | 0.54 | 118 940.12 | 0.75 | |||
| Guatemala | 14.07 | 7.45 | 5.36 | 6.57 | 21.00 | 19.96 | 10.21 | 14.67 | 0.72 | 116 696.91 | 0.74 | |
| Panamá | 14.99 | 11.49 | 15.95 | 18.08 | 18.36 | 12.25 | 4.20 | 4.66 | 0.03 | 73 915.26 | 0.47 | |
| Costa Rica | 18.42 | 8.72 | 6.27 | 5.02 | 25.67 | 35.49 | 0.37 | 0.03 | 51 632.69 | 0.33 | ||
| Belice | 0.53 | 0.98 | 1.96 | 1.79 | 21.10 | 44.56 | 12.08 | 16.99 | 23 671.45 | 0.15 | ||
| El Salvador | 1.65 | 1.12 | 2.15 | 1.97 | 57.41 | 35.58 | 0.12 | 21 710.71 | 0.14 | |||
| Subtotal | 539 718.61 | 3.41 | ||||||||||
| Sur | Brasil | 20.39 | 13.02 | 12.60 | 14.15 | 20.82 | 15.29 | 2.00 | 1.67 | 0.06 | 8 808 039.49 | 55.58 |
| Colombia | 7.84 | 7.32 | 10.01 | 14.41 | 42.00 | 15.84 | 2.34 | 0.23 | 0.01 | 1 136 725.73 | 7.15.347 | |
| Venezuela | 6.01 | 6.14 | 13.59 | 21.12 | 29.28 | 18.49 | 3.39 | 1.97 | 0.01 | 919 696.09 | 5.8 | |
| Bolivia | 13.84 | 7.30 | 17.09 | 24.42 | 28.03 | 7.49 | 1.27 | 0.56 | 845,814.43 | 5.34 | ||
| Perú | 16.31 | 12.72 | 10.03 | 19.97 | 31.75 | 7.90 | 0.82 | 0.67 | 839 690.29 | 5.3 | ||
| Paraguay | 64.62 | 16.16 | 8.13 | 6.00 | 4.98 | 0.12 | 0.01 | 0.1 | 334 011.68 | 2.11 | ||
| Uruguay | 75.11 | 17.24 | 3.90 | 3.52 | 0.23 | 243 178.38 | 1.53 | |||||
| Argentina | 77.03 | 8.86 | 4.93 | 5.22 | 3.87 | 0.08 | 214 547.26 | 1.35 | ||||
| Guyana | 4.01 | 3.09 | 4.09 | 8.75 | 32.16 | 21.36 | 11.42 | 14.64 | 0.47 | 211 818.62 | 1.34 | |
| Ecuador | 10.69 | 5.91 | 11.07 | 12.85 | 24.48 | 8.43 | 5.41 | 12.47 | 8.68 | 206 362.40 | 1.3 | |
| Surinam | 2.47 | 0.74 | 8.33 | 29.3 | 55.32 | 3.83 | 0.01 | 145 688.92 | 0.92 | |||
| G. Francesa | 0.91 | 2.15 | 20.54 | 23.35 | 45.63 | 7.18 | 0.23 | 0.01 | 83 701.95 | 0.53 | ||
| Chile | 100 | 275.22 | 0.01 | |||||||||
| Subtotal | 13 989 550.46 | 88.27 | ||||||||||
| Caribe | Cuba | 4.27 | 1.56 | 1.63 | 17.37 | 60.65 | 14.04 | 0.49 | 123 628.39 | 0.78 | ||
| R. Dominicana | 6.03 | 5.78 | 21.71 | 27.76 | 37.68 | 1.04 | 50 434.01 | 0.32 | ||||
| Haití | 3.18 | 1.10 | 2.80 | 16.34 | 42.74 | 28.53 | 3.91 | 1.41 | 29 355.49 | 0.19 | ||
| Bahamas | 93.69 | 5.19 | 1.10 | 0.84 | 12 660.57 | 0.08 | ||||||
| Jamaica | 12.57 | 5.45 | 5.74 | 5.63 | 43.31 | 24.66 | 2.64 | 11 849.82 | 0.07 | |||
| Puerto Rico | 2.31 | 4.43 | 19.88 | 32.00 | 41.39 | 9672.60 | 0.06 | |||||
| T y Tobago | 80.29 | 7.00 | 4.56 | 2.01 | 6.14 | 5052.35 | 0.03 | |||||
| Guadalupe | 88.91 | 11.09 | 1671.15 | 0.01 | ||||||||
| Martinica | 75.29 | 24.22 | 0.49 | 1102.69 | 0.01 | |||||||
| Dominica | 42.40 | 56.48 | 1.12 | 773.23 | 0.005 | |||||||
| Santa Lucia | 60.39 | 37.80 | 1.81 | 635.8 | 0.004 | |||||||
| A y Barbuda | 100 | 448.24 | 0.003 | |||||||||
| Barbados | 100 | 444.92 | 0.003 | |||||||||
| San Vicente | 60.52 | 37.36 | 1.88 | 0.24 | 344.96 | 0.002 | ||||||
| Granada | 86.00 | 14.00 | 320.93 | 0.002 | ||||||||
| Islas Turcas | 60.40 | 39.60 | 298.49 | 0.002 | ||||||||
| Islas Vírgenes | 98.23 | 1.77 | 283.92 | 0.002 | ||||||||
| Antillas | 100 | 277.93 | 0.002 | |||||||||
| Islas Caimán | 100 | 185.43 | 0.001 | |||||||||
| San Cristóbal | 91.96 | 8.04 | 167.75 | 0.001 | ||||||||
| Montserrat | 84.36 | 107.13 | 0.001 | |||||||||
| Islas Británicas | 95.68 | 84.08 | 0.001 | |||||||||
| Anguila | 100 | 64.61 | 0.0004 | |||||||||
| Subtotal | 249 864.45 | 1.58 | ||||||||||
| Total | 21.75 | 11.13 | 11.76 | 14.52 | 23.5 | 13.40 | 2.05 | 1.73 | 0.17 | 15 848 524.00 | ||
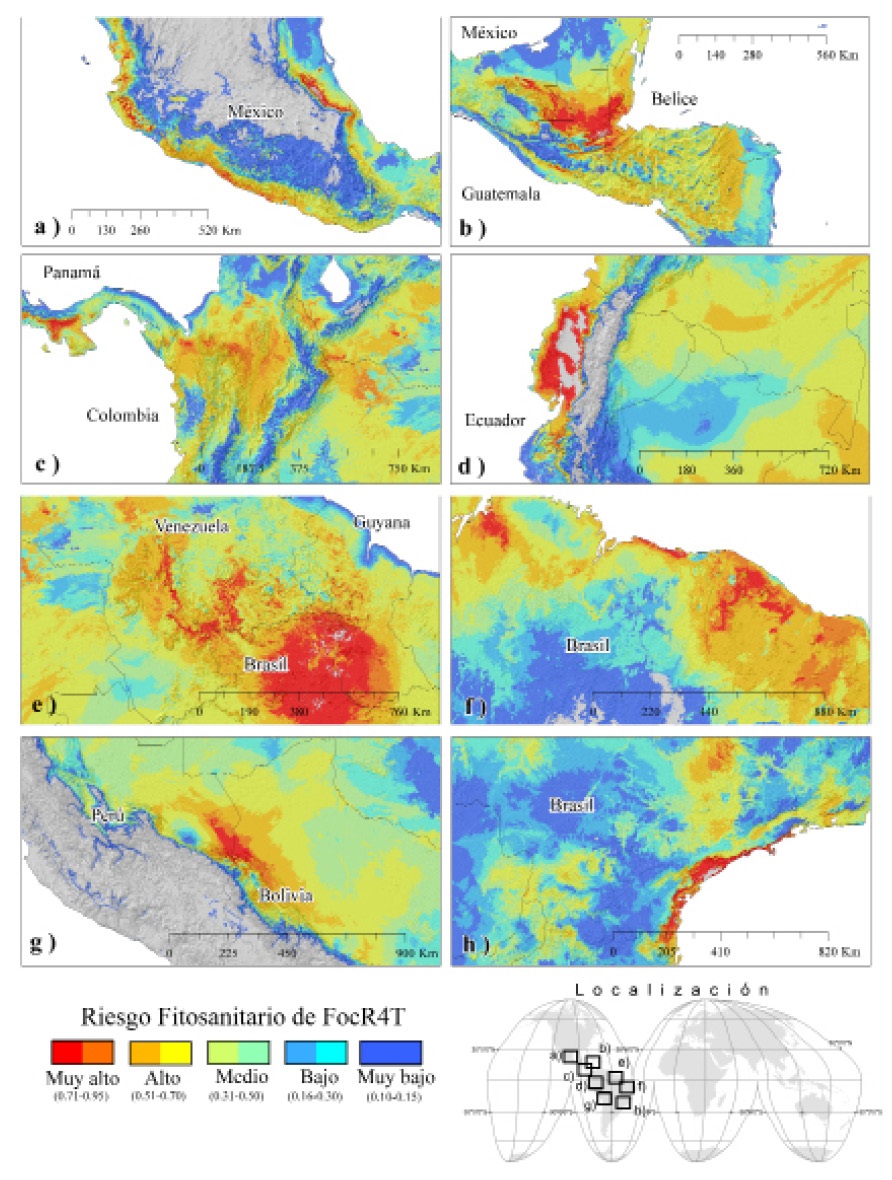
Con mayor detalle, en la Figura 5 se observan la regiones prioritarias territorialmente ubicadas en: a) la porción centro-sur y costa del océano pacífico en México; b) el centro-norte de Guatemala, sur de Belice y este de Chiapas en México; c) el centro norte de Colombia y porción central de Panamá; d) la costa de Ecuador; e) noroeste de Brasil, sur de Guyana y una porción del centro de Venezuela; f) la región noreste de Brasil; g) una región colindante entre Perú y Bolivia, y h) una porción en la región del sureste de Brasil. Esto sugiere que son áreas estratégicas donde se deben intensificar las acciones preventivas como exploración, monitoreo y vigilancia epidemiológica fitosanitaria.
El análisis de correlación de Pearson reveló que las variables (BIO16, BIO6 y BIO18) presentan una correlación positiva, es decir, los valores más altos en las zonas cercanas a los trópicos corresponden a los valores más altos de RF_FocR4T. Cabe señalar que la estacionalidad de la temperatura (BIO4) presentó una correlación negativa con respecto a la distribución espacial del RF_FocR4T, con valores mínimos en la zona cercana al Ecuador (Cuadro 7).
Cuadro 7 Correlación de Pearson (r) entre el RF_FocR4T) contra las variables bioclimáticas de mayor contribución porcentual.
| RF_FocR4T | BIO16 | BIO14 | BIO6 | BIO18 | |
| RF_FocR4T | 1 | ||||
| BIO16 | 0.32 | 1 | |||
| BIO14 | -0.25 | -0.41 | 1 | ||
| BIO6 | 0.34 | 0.43 | -0.68 | 1 | |
| BIO18 | 0.07 | 0.39 | 0.08 | -0.22 | 1 |
BIO16: precipitación del trimestre más húmedo, BIO4: estacionalidad de la temperatura, BIO6: la temperatura mínima del mes más frio y BIO18: precipitación del trimestre más cálido.
Lo anterior revela que la tendencia de la predicción del riesgo fitosanitario de la Fusariosis de la musáceas se da hacia las zonas más cercanas al Ecuador, por presentar una mayor precipitación del trimestre más húmedo, una mayor temperatura mínima del mes más frío, una mayor precipitación del trimestre más cálido, así como una menor estacionalidad de la temperatura. Es decir, los lugares cercanos al ecuador presentan las mejores condiciones para el desarrollo de la enfermedad, sin restarle importancia a las zonas con menor nivel de riesgo distribuidas en América pantropical, visibles en la Figura 5.
Los periodos de precipitación con más humedad y más cálidos, con una temperatura más fría, favorecen de manera positiva a las condiciones óptimas para FocR4T, y permiten caracterizar superficies que cuentan las condiciones óptimas en regiones próximas al ecuador. Por ello se identificaron porciones compactas con la condición de riesgo fitosanitario, muy alto en países como Brasil, Venezuela, Guyana, Colombia y Ecuador. Sin embargo, la estacionalidad de la temperatura se correlacionó en sentido inverso con una tendencia negativa (r = 0.28), es decir, a menor variabilidad térmica se estima un mayor RF. Lo anterior revela que la temperatura juega un papel importante en el desarrollo de la enfermedad.
Conclusiones y recomendaciones
Nuestros resultados se asemejan a los obtenido por Salvacion et al. (2019a), ya que la posible ocurrencia de FocR4T está muy relacionada a condiciones de menor variabilidad climática territorialmente en superficies próximas al Ecuador, ya que la severidad del marchitamiento por Fusarium spp se incrementa en relación con la precipitación y la temperatura (Peng et al., 1999). Incluso, ante el efecto del cambio climático, factores como baja en la precipitación en periodos de mayor humedad y altas precipitaciones en periodos más cálidos generan mayor probabilidad de ocurrencia del patógeno, hay quienes concluyen que para 2050 hasta un 91% de áreas aptas para cultivo de banano en Filipinas estarán infectadas de FocR4T (Salvacion et al., 2019b).
La totalidad de la región de América pantropical presentó algún nivel de RF_FocR4T. Sin embargo, se delimitaron de manera específica seis áreas compactas distribuidas en Norteamérica, Centroamérica, Sudamérica y el Caribe. Los países en orden de potencial afectación en Sudamérica son Brasil, Colombia, Venezuela y Perú, principalmente; para el caso de Centroamérica son Nicaragua, Honduras, Guatemala y Panamá entre otros; para el Caribe serían Cuba, República Dominicana, Haití, Bahamas, Jamaica, entre otros, y para Norteamérica, solo México.
Las ONPF y las ORPF, que integran América pantropical, pueden apoyarse en el modelo cartográfico de RF_FocR4T como insumos de respaldo en el manejo del patógeno, ante una posible introducción, establecimiento o dispersión entre los países que integran las organizaciones. Se puede considerar un insumo técnico/científico para dar soporte en la toma de decisiones en el proceso de vigilancia epidemiológica fitosanitaria como complemento de los programas de manejo integrado de plagas y enfermedades (Siamak et al., 2018; Thangavelu et al., 2020). Este tipo de modelado se concibe como una estrategia preventiva ante el riesgo fitosanitario de la Fusariosis de las musáceas a diferentes escalas geográficas.
Los organismos de protección fitosanitaria, tanto regionales como nacionales, (ONPF y ORPF), se deben coordinar con los centros de investigación para la elaboración de manera conjunta de modelos de riesgo fitosanitario más robustos que puedan fortalecer las acciones y estrategias operativas, entre las que destacan la identificación de sitios o áreas óptimas para realizar actividades de exploración, el diseño de rutas de trampeo y vigilancia, así como el establecimiento de parcelas centinelas y viveros trampa (Pérez-Vicente, 2015; SENASICA, 2019).
Una fortaleza del modelo geoespacial de RF_FocR4T desarrollado en la presente investigación es que puede ser actualizado con nuevas detecciones del patógeno provenientes de fuentes oficiales o literatura científica, así como información reciente de movilidad de material vegetal propagativo, lo que permite obtener un replanteamiento del escenario actual, ajustado a la realidad epidémica de la enfermedad. Sin embargo, es recomendable profundizar en las posibles consecuencias de FocR4T en América pantropical ante el inminente cambio climático.
Como investigación a futuro, se recomienda realizar trabajos a mayor resolución espacial con la incorporación del factor suelo como elemento clave en la propagación de cepas patogénicas (Orr y Nelson, 2018). Para ello se puede considerar el uso de modelos digitales de elevación que abarquen las zonas productoras de plátano, incorporar la geometría de la red hidrográfica (análisis de afluentes), así como el riesgo potencial de inundación, hacia las áreas plataneras o bananeras que se encuentren en las partes bajas de la cuenca en un contexto de análisis de conectividad y dinámica de parches de campos plataneros.

