











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La reducida disponibilidad de fósforo (P) en el suelo es uno de los principales factores que limitan el crecimiento y rendimiento de los cultivos mundialmente (Lynch, 2013). Alcalá et al. (2001) señalan que en andosoles, la reducida disponibilidad de P (< 10 mg kg-1) y la alta capacidad de retención de fosfatos (> 85 %), debido a la alta composición mineral de óxidos de Fe y Al, son factores limitantes en la producción de maíz (Zea mays L.) de la Meseta P'urhépecha. La sostenibilidad de los sistemas agrícolas de esta región, requiere de la identificación y desarrollo de genotipos eficientes para adquirir y utilizar el P sea del suelo o del fertilizante (Lynch, 2013).
El mejoramiento genético de la eficiencia en el uso de P (EUP), definida como el incremento de la producción de materia seca o rendimiento por unidad del nutriente absorbido (Wang et al., 2010) se ha realizado con variedades mejoradas derivadas de un reducido acervo de germoplasma y predominantemente adaptadas a una agricultura intensiva y ambientes óptimos (Manschadi et al., 2014). En contraste, las variedades tradicionales de maíz que se han desarrollado en ambientes específicos o marginados con reducida disponibilidad de nutrientes, representan una fuente importante de variación para la selección de genotipos adaptados a sistemas rurales (Newton et al., 2010). Por ejemplo, las variedades nativas de la Meseta P'urhépecha están adaptadas a los ambientes de reducida fertilidad y poseen características en la raíz que no son comunes en el germoplasma élite (Bayuelo-Jiménez et al., 2011). Las variedades eficientes a P poseen raíces axilares (seminales o nodales) más superficiales y ramificadas que incrementan la absorción de P en la superficie del suelo y el crecimiento durante la etapa de plántula (Bayuelo-Jiménez et al., 2012) y etapa vegetativa (Bayuelo-Jiménez et al., 2011). A pesar de estos hallazgos, no existen estudios sistemáticos que evalúen la eficiencia a P en diferentes condiciones agroecológicas y etapas fenológicas del cultivo.
El objetivo de este estudio fue evaluar la variabilidad genotípica del en maíz nativo de la Meseta Puhrépecha para eficiencia y respuesta a P durante la etapa de floración y de esta manera probar la hipótesis de que existen variedades nativas de maíz eficientes en adquirir y utilizar el P del suelo.
Materiales y métodos
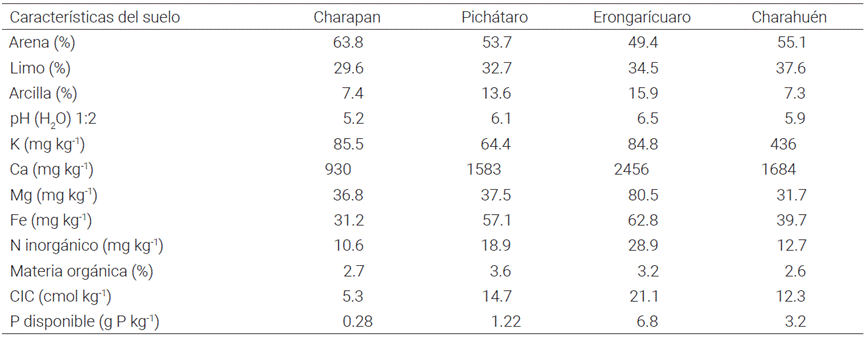
El presente estudio se realizó en condiciones de temporal, de mayo a diciembre de 2014. Las parcelas experimentales se ubicaron en los municipios de Charapan (19° 39' N y 101° 54' O), Erongarícuaro (19° 35' N y 101° 43' O), Pichátaro (19° 34' N y 101° 43' O) y Charahuén (19° 31' N y 101° 42' O), en el estado de Michoacán. El clima es templado sub-húmedo con lluvias en verano [C (w2) (w)], con una temperatura media anual de 20 oC y altura de 2260 a 2414 msnm (Muñoz, 2003). En el Cuadro 1 se muestran las características físico-químicas del suelo.
Los experimentos incluyeron dos niveles de fertilización con una baja (25 kg ha-1) y alta (50 kg ha-1) aplicación de P2O5 proveniente de superfosfato triple, combinados con dos testigos de maíz (cv. Ocelote y un genotipo de maíz nativo de la región) y 25 genotipos de maíz provenientes del programa de mejoramiento del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) e integrados por genotipos nativos de la Meseta P'urhépecha, así como maíces mejorados de amplia base genética. De acuerdo con el tiempo (días) en alcanzar el 50 % de floración masculina los materiales se clasificaron como: precoces (de 75 a 80 d con 33 materiales), intermedios (de 85 a 95 d con 40 materiales) y tardíos (de 95 a 110 d con 35 materiales), y en cuatro grados de mejoramiento: nativos (N), nativos mejorados (NM), híbridos × nativos (H×N) e híbridos mejorados (S) (Cuadro 2). A todas las unidades experimentales se les aplicó una dosis de 60 kg N ha-1 en forma de urea a la siembra y una de 60 kg N ha-1 a los 42 d después de la siembra.
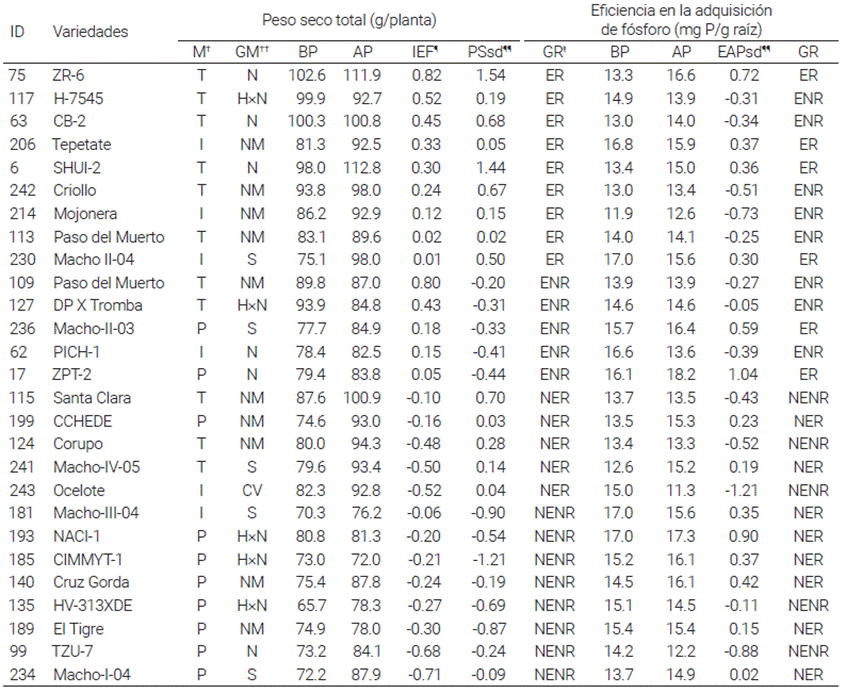
†M: Madurez: Precoz (P), Intermedio (I) y Tardío (T). ††GM: Grupo de Mejoramiento: Nativo (N), Nativo Mejorado (CM), Híbrido × Nativo (H×N), Sintético (S), Cultivar (cv). *IEF: Índice de eficiencia a fósforo, obtenido del análisis de componentes principales. **PSsd, EAPsd: Valor estandarizado del peso seco total y eficiencia en la adquisición de P en condiciones de AP. GRŧ: Respuesta a fósforo; se expresó como el peso seco total o EAP en condiciones de AP. Se definieron cuatro categorías: eficiente y responde (ER), eficiente y no responde (ENR), no eficiente y responde (NER), no eficiente y no responde (NENR).
Cuadro 2 Medias del peso seco total y eficiencia en la adquisición de fósforo en variedades de maíz cultivadas en un suelo con reducida (BP) y alta (AP) dosis de fertilización fosforada. Los genotipos se ordenaron de acuerdo con el índice de eficiencia a fósforo (IEF) del peso seco total.
Las plantas se muestrearon en la etapa de floración. Las hojas, tallo, raíz y espiga de cada conjunto de cuatro plantas/genotipo se deshidrataron en una estufa a 65 °C por 72 h, para luego registrar el peso seco. El tejido seco se molió en un molino Wiley-Thomas con una malla de 1 mm y se utilizaron sub-muestras de 70 mg para calcinarlas en una mufla (CEM®, mod. MAS-7000, USA) a 500 °C por 8 h. La estimación de la concentración de P se realizó por colorimetría mediante la reacción con una solución de molibdato [(NH4) 6M°7O24] y su medición en un espectrofotómetro de UV/VIS (Perkin Elmer®, mod. Lamda 40, Uberlingen, Germany) a una longitud de 880 nm (Murphy y Riley, 1962). Se calculó el contenido de P (mg g-1) de raíz, vástago y espiga mediante la multiplicación de la concentración de P con la materia seca acumulada en cada órgano. La eficiencia en la adquisición de fósforo (EAP) es una medida del índice de la absorción de P por unidad de peso seco de la raíz (mg P g-1), y se calculó con base en la relación del contenido total de P y el peso seco de la raíz en gramos (Fageria et al., 2008).
Se empleó un diseño de bloques completos al azar con un arreglo de tratamientos en parcelas divididas con cinco repeticiones por tratamiento. Los niveles de P correspondieron a la parcela principal y los genotipos, distribuidos aleatoriamente dentro de cada parcela, correspondieron a las sub-parcelas. Cada unidad experimental consistió de dos surcos de 5 m de longitud por genotipo. Las semillas se sembraron a una profundidad de 6 cm y a una distancia de 25 cm entre cada planta y a 60 cm de distancia entre cada surco. Las diferencias entre genotipos en respuesta a los niveles de P aplicados se evaluaron mediante análisis de varianza (SAS, 2000). Previo al análisis se realizó una prueba de Bartlett para la homogeneidad de varianza en todas las variables incluidas (SAS, 2000). La varianza del error fue heterogénea para algunas variables, así que los datos fueron transformados a base logarítmica y raíz cuadrada (SAS, 2000). Se utilizó la prueba de Tukey con valores de P ≤ 0.05 para comparar las diferencias entre genotipos y tratamientos de P en las distintas variables evaluadas.
La eficiencia a P, definida como el crecimiento sostenido en condiciones de baja disponibilidad de P, se determinó de acuerdo con el índice de eficiencia a P (IEF) (Pan et al., 2008) obtenido mediante un análisis de componentes principales (ACP) a partir de los datos estandarizados de crecimiento en condiciones de bajo contenido de P (BP) y relativo a la condición en alto contenido de P (AP) (SAS, 2000). Los valores del IEF de cada genotipo se calcularon de acuerdo con el componente principal retenido (CP) y su contribución relativa (CR): IEF = 27∑ i=1 CPi x CRi .
Posteriormente, con los datos de IEF por genotipo se hizo un análisis de conglomerados para agrupar por categorías de eficiencia [eficientes (E), moderadamente eficientes (ME) e ineficientes (I)], y en cuatro categorías de respuesta a P de acuerdo con el IEF en combinación con la respuesta a P (peso seco total y EAP en AP) [eficientes y con respuesta (ER); no eficientes y con respuesta (NER), no eficientes y sin respuesta (NENR), y eficientes y sin respuesta (ENR)]. El análisis de conglomerados se realizó según el método de Ward (Romerburg, 1984).
Resultados y discusión
Según el análisis de componentes principales (CP), de las características de crecimiento de los genotipos evaluados los componentes CP1 al CP4 contribuyeron con 30, 20, 18 y 12 % de la variación entre grupos. Estos componentes en conjunto aportaron 80 % de la variación total observada. La variabilidad del primer componente (CP1) estuvo determinada por valores positivos de: peso seco de la raíz (0.87), peso seco de vástago (0.87) y peso seco total (0.90) en condiciones de bajo fósforo (BP). El componente 2 (CP2) estuvo influenciado positivamente por el contenido de P en vástago (0.67) en BP y los valores relativos a la condición de alto contenido de P (AP) en vástago (0.81) y espiga (0.67). En el componente 3 (CP3) ejercieron mayor peso los índices relativos del peso seco de vástago (0.74), peso seco total (0.69) y eficiencia en adquisición de P (EAP) (0.62). En el componente 4 (CP4) influyó positivamente el peso seco de la espiga (0.86).
En la Figura 1 se presenta la formación de los grupos de eficiencia (combinada de las cuatro localidades) con base en sus parámetros de crecimiento en BP y sus valores relativos al tratamiento de AP. El Grupo 1 (Índice de eficiencia a P, IEF < 0.60) consistió de cinco genotipos eficientes, de los cuales 20 y 80 % fueron de madurez intermedia y tardía. El Grupo 2 (0.11 < IEF < -0.39) estuvo compuesto por 11 genotipos moderadamente eficientes con 27, 19 y 54 % de madurez precoz, intermedia y tardía. El Grupo 3 (IEF < -0.39) lo integraron 11 genotipos ineficientes a P, la mayoría de los cuales fueron variedades precoces (64 %). La comparación de la eficiencia a P combinada de los 27 genotipos mostró que la clasificación de los genotipos evaluados fue constante en los tres grupos de eficiencia. Los genotipos 63, 75, 109, 117 y 127 fueron los más eficientes a P. Estos genotipos acumularon mayor peso y contenido de P en la condición de BP (Figuras 2 y 3).
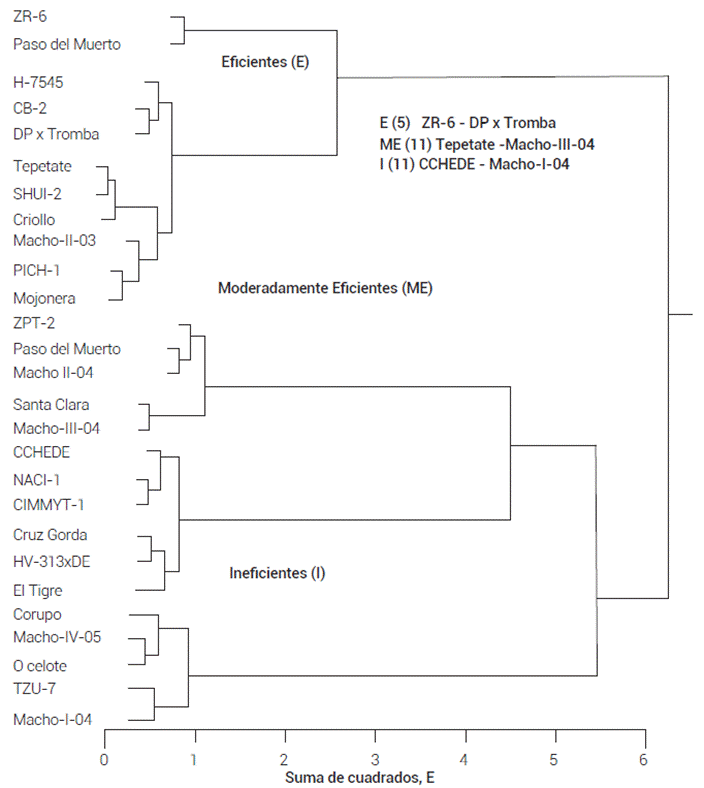
Figura 1 Clasificación combinada de 27 genotipos de maíz de acuerdo con el índice de eficiencia a fósforo (IEF), mediante el método de agrupación de Ward. Los grupos I, ME y E representan los genotipos ineficientes (11), moderadamente eficientes (11) y eficientes (5).
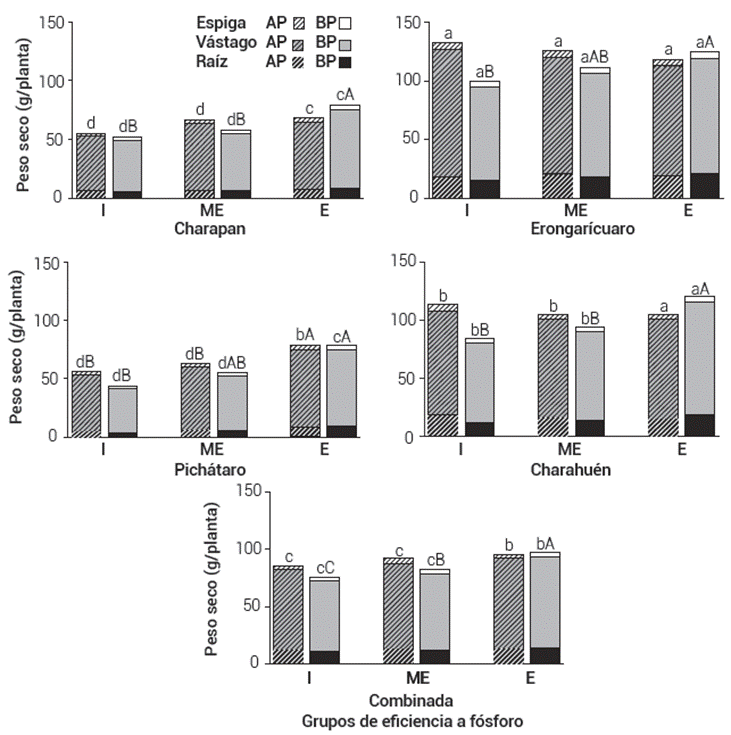
Figura 2 Peso seco de la raíz, vástago y espiga en condiciones de reducida (I), moderada (ME) y alta eficiencia (E) a fósforo. Las barras representan la media de 11, 11 y 5 genotipos. Las medias con letras iguales entre localidades (minúsculas) y entre grupos de eficiencia (mayúsculas) por nivel de fósforo son estadísticamente iguales entre sí (Tukey, 0.05).
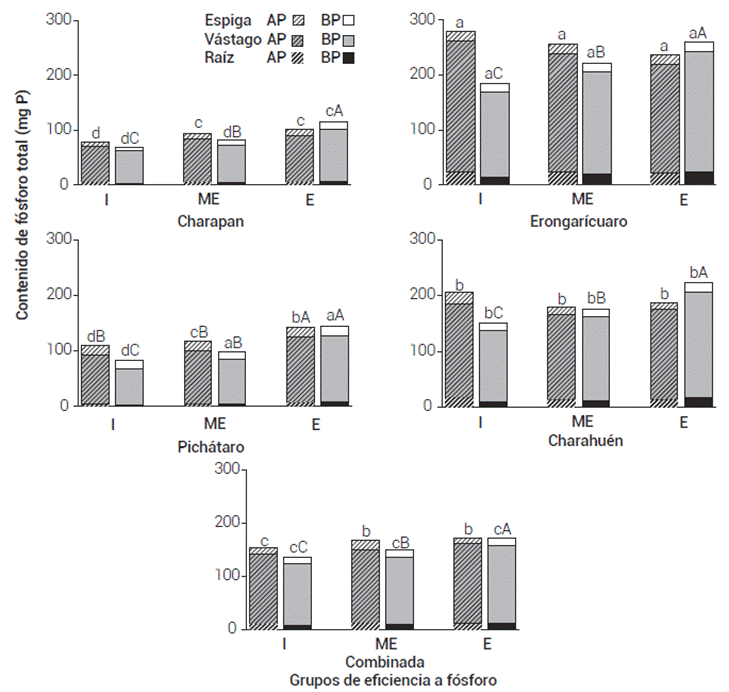
Figura 3 Contenido de fósforo en raíz, vástago y espiga en condiciones de reducida (I), moderada (ME) y alta eficiencia (E) a fósforo. Las barras representan las medias de 11, 11 y 5 genotipos. Las medias con letras iguales entre localidades (minúsculas) y entre grupos de eficiencia (mayúsculas) por nivel de fósforo son estadísticamente iguales entre sí (Tukey, 0.05).
Los 27 genotipos se separaron en cuatro grupos de acuerdo con su respuesta a P (expresada por el peso seco total y EAP en AP, estandarizado) e IEF. Las categorías fueron: (i) eficientes y con respuesta (ER), (ii) ineficientes y con respuesta (NER), (iii) ineficientes y sin respuesta (NENR) y (iv) eficientes y sin respuesta (ENR) (Figura 4). En la comparación de la respuesta y eficiencia a P se identificaron nueve genotipos eficientes en BP, que respondieron a la mayor aplicación del mismo (ER) (Figura 4; Cuadro 2). Los genotipos 6, 63, 75, 113, 117, 206, 214, 230 y 242 (testigo maíz nativo) exhibieron una similar acumulación del peso seco total en ambos ambientes.
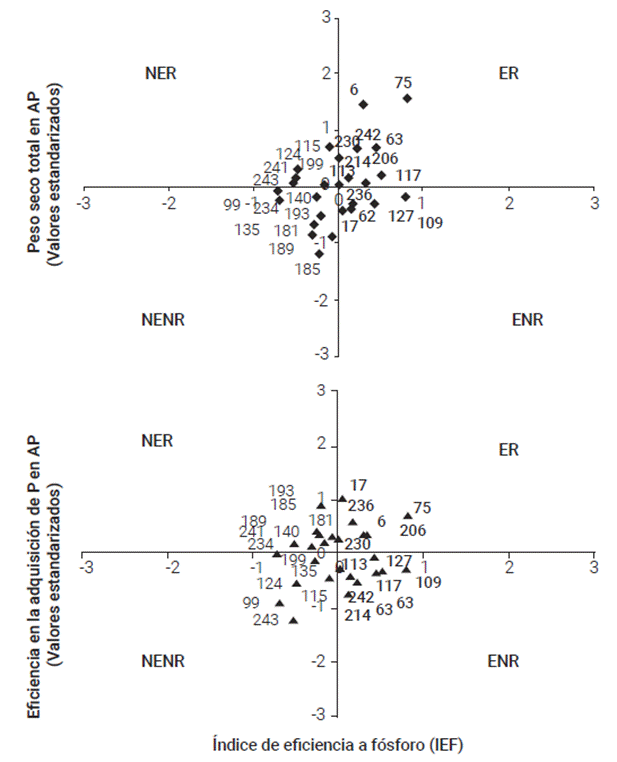
Figura 4 División de los 27 genotipos de maíz de acuerdo con el índice de eficiencia a fósforo (IEF) y de los valores estandarizados del peso seco total y la eficiencia en la adquisición de fósforo (EAP) en AP. Los valores estandarizados del peso seco total y EAP se estimaron de acuerdo con la siguiente función: Xs = (X X)/SD. Las categorías están representadas con genotipos eficientes con respuesta (ER), no eficientes con respuesta (NER), eficientes sin repuesta (ENR) y no eficientes y sin respuesta (NENR), respectivamente. Se indica el número de cada genotipo.
Cinco genotipos (115, 124, 199, 241 y 243) (testigo cv. Ocelote) presentaron una alta respuesta a P, manifestada por un incremento en el peso seco total pero que fueron ineficientes en la acumulación de materia seca en BP; mientras que ocho genotipos (99, 135, 140, 181, 185, 189, 193 y 234) fueron ineficientes a BP y no respondieron a la condición de AP. En la categoría eficientes y sin respuesta (ENR) destacan cinco genotipos (17, 62, 109, 127, 236) de moderada y alta eficiencia, que incrementaron el peso seco total en el ambiente de BP, pero no respondieron a la mayor disponibilidad del mismo. En la clasificación combinada de las cuatro localidades se determinó que los genotipos SHUI-2 (6), ZR-6 (75), Paso del Muerto (113), Tepetate (206) y Macho II-04 (230) fueron los más eficientes en BP y que también respondieron a la alta fertilización.
En este estudio se adoptaron dos índices para seleccionar genotipos eficientes a P. El primero estimó el crecimiento relativo del cultivo obtenido en condiciones limitantes de P, con respecto al crecimiento alcanzado en condiciones nutrimentales óptimas (Fageria et al., 2008). Adicionalmente, se utilizó el sistema propuesto por Gerloff (1977) para clasificar los genotipos de maíz con base en el crecimiento o acumulación de P en condiciones óptimas y de deficiencia. Este sistema incluyó genotipos: 1) Eficientes y con respuesta (ER), plantas con mayor crecimiento con reducida disponibilidad de P y que respondieron a los altos niveles de fertilización; 2) Ineficientes y con respuesta (NER), plantas con reducido crecimiento en BP y con alta respuesta a la adición del nutriente; 3) Eficientes y sin respuesta (ENR), plantas con alto crecimiento en bajo nivel de P, pero que no responden a la adición de P; y 4) Ineficientes y sin respuesta (NENR), plantas con reducido crecimiento en BP y sin respuesta a condiciones de alto nivel nutrimental.
Con este sistema de categorización fue posible obtener información para comparar los genotipos que se desarrollaron en condiciones de deficiencia de P en el suelo (eficientes vs. ineficientes) y condiciones adecuadas de fertilización (con respuesta vs. sin respuesta) (Figura 4). Los resultados obtenidos sugieren que el uso de ambos sistemas es útil para identificar germoplasma eficiente a P y contrastar su eficiencia en diferentes condiciones edáficas (Bayuelo-Jiménez y Ochoa-Cadavid, 2014; Manschadi et al., 2014).
En la comparación de la eficiencia en la adquisición de fósforo (EAP), hubo una reducida variación en la distribución de los genotipos. Mientras que los genotipos 6, 75, 206, 230 y 236 se mantuvieron como los más eficientes en la EAP en ambos ambientes; los genotipos 63, 113, 117, 214 y 242 (testigo, maíz nativo de la región) se clasificaron como los más eficientes en BP, pero con una reducida respuesta a la condición de AP. Estos genotipos fueron muy eficientes para adquirir este nutriente en BP, pero no respondieron a la alta adición del mismo. La comparación de la eficiencia y respuesta a P basada en el peso seco total y EAP identificó a cuatro genotipos eficientes en BP y que respondieran a la alta dosis de fertilización. Los resultados de este estudio mostraron que existe variación genotípica en maíz nativo para eficiencia en la adquisición de P, y que esta variación está acentuada en los genotipos de maduración tardía.
La eficiencia en el uso de fósforo (EUP) es el producto de la eficiencia en la adquisición (EAP; P total en la planta por unidad de nutriente suministrado) y utilización de P (EUtP; el crecimiento y rendimiento del cultivo por unidad de P absorbido) (Wang et al., 2010). Una mayor EUP puede conseguirse mediante estrategias que mejoren la exploración, adquisición y removilización de P del suelo (Manschadi et al., 2014; Simpson et al., 2011). Debido a la relativa inmovilidad y heterogénea distribución de P en la capa arable del suelo, la configuración de las raíces en espacio y tiempo son características clave para optimizar su adquisición (Lynch, 2013).
El sistema radical de maíz está conformado por raíces nodales que absorben los nutrientes móviles e inmóviles del suelo y que responden diferencialmente al estrés impuesto por el ambiente (Postma et al., 2014). Los genotipos eficientes que incrementan el número de raíces nodales en condiciones de reducida disponibilidad de P tienen una mayor capacidad para mantener un crecimiento continuo y para adquirir P en la superficie del suelo (Zhu et al., 2005). El ángulo de las raíces axiales, el número de raíces adventicias, el número de raíces seminales y nodales, la densidad y longitud de pelos radicales y la formación de aerénquima, son algunas de las características que favorecen la adquisición de P del suelo (Simpson et al., 2011). En maíces nativos de la Meseta P'urhépecha, la mayor absorción de P del suelo está mediada por el incremento del número de raíces nodales, una mayor ramificación de raíces laterales y el desarrollo de pelos radicales más densos y largos (Bayuelo-Jiménez et al., 2011).
La asociación con hongos micorrícicos arbusculares (HMA) también incrementa la superficie de absorción de la raíz y, por tanto, la adquisición de nutrientes minerales como P, Cu y Zn. Sin embargo, los HMA son ubicuos y están menos sujetos a selección. A diferencia de las características que definen la arquitectura de la raíz, la información sobre la variabilidad genotípica de HMA es reducida. Además, los resultados de diversas investigaciones sugieren que las características de la raíz actúan de forma aditiva y sinérgica con los HMA (Lynch, 2013).
La eficiencia en la adquisición de fósforo (EAP) está fuertemente afectada por el estado hídrico del suelo (McBeath et al., 2012). Los prolongados periodos de sequía combinados con altas temperaturas afectan la productividad agrícola al reducir la eficiencia en la adquisición de nutrientes (Lynch, 2013). El déficit hídrico, reduce el crecimiento de la raíz y la EAP al alterar el transporte de P en el suelo. En condiciones de déficit hídrico, la eficiencia del fertilizante fosforado puede disminuir hasta 3 %, mientras que durante el periodo de lluvia incrementa hasta 30 % (McBeath et al., 2012). A pesar de que no se evaluó el efecto del estrés hídrico, es plausible que éste tenga implicaciones en la productividad de los sistemas agrícolas de temporal de ésta y otras regiones de México. El desarrollo de genotipos eficientes en el uso de P y agua son aspectos importantes en la sostenibilidad económica y ambiental de este sector agrícola.
Aunado a la configuración y morfología de la raíz, la fenología de la planta beneficia la absorción de nutrientes del suelo (Lynch, 2013). La fenología determina la duración del periodo de adquisición y utilización de los nutrientes y sincroniza las relaciones de fuente y demanda del recurso (Nord y Lynch, 2009). Una mayor duración del crecimiento del vástago promueve una mayor utilización fisiológica del nutriente (Bayuelo-Jiménez y Ochoa, 2014). Los resultados confirman que independientemente de la localidad estudiada, los genotipos eficientes de maduración tardía acumularon mayor peso y contenido de P en el vástago y espiga que los genotipos ineficientes, mayormente precoces (Figuras 2 y 3). En la mayoría de las condiciones de cultivo, la disponibilidad de P en la solución del suelo fue limitante (0.28 a 6.8 mg P kg-1), pero los genotipos tardíos tuvieron una mayor oportunidad de adquirir P durante más tiempo.
Conclusiones
Los genotipos de maíz de la Meseta P'urhépecha mostraron una gran variabilidad en el crecimiento vegetal, eficiencia y respuesta a fósforo, en etapa de floración. La eficiencia a fósforo estuvo determinada por una mayor producción de materia seca en la raíz, el vástago y la espiga; una mayor asignación de fósforo hacia los vástagos, y una mayor eficiencia en la absorción de fósforo por unidad de peso seco de la raíz. La maduración tardía de los genotipos eficientes favoreció una mayor adquisición de fósforo y crecimiento en la planta. Estos resultados indican que existe variabilidad genotípica para la eficiencia a fósforo en el germoplasma de maíz.

