











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introducción
Los bosques templados del mundo se encuentran en su mayoría en las regiones Neártica y Paleártica, formando parte del conjunto de vegetación típico de latitudes no tropicales, en donde el clima tiene su estacionalidad anual marcada y un invierno durante el cual la temperatura cae por debajo de 0 °C (Olson et al., 2001; Challenger, 2003). En México, este tipo de bosque está asociado estrechamente con las zonas montañosas debido a la ubicación geográfica del país y a la relación que existe entre el clima y la altitud (Tamayo, 1990; Sánchez et al., 2003).
Según la Organización de las Naciones Unidas para la Alimentación y Agricultura (FAO, 2010), 35% de la superficie de México se encuentra ocupada por ecosistemas forestales, de los cuales 24% corresponden a bosques templados (CONAFOR, 2014), en donde la zona de mayor concentración de ecosistemas boscosos es la Sierra Madre Occidental (Challenger y Soberón, 2008). En esta región se presenta gran diversidad de asociaciones de Pinus L., Quercus L. y Arbutus L. (González-Elizondo et al., 2007). La importancia de este tipo de vegetación no es sólo por su alta diversidad, sino también porque las especies de Pinus y Quercus son los árboles de mayor interés económico (Sánchez et al., 2003).
El manejo forestal maderable realizado en las últimas décadas ha sido muy selectivo, sobre todo en las mejores especies de árboles e individuos, ocasionando cambios en la composición, la estructura y la función del bosque. Este manejo crea condiciones de desarrollo para especies de mayor importancia económica, y disminuye la proporción de especies de importancia ecológica por tener menor valor económico (Fregoso et al., 2001; Hernández-Salas et al., 2013, 2018).
El conocimiento de la estructura constituye uno de los fundamentos principales para entender el funcionamiento de los ecosistemas, lo cual puede aportar elementos de decisión para contribuir al manejo adecuado de los bosques (Cascante y Estrada, 2001; Franklin et al., 2002; Aguirre et al., 2003; Castellanos-Bolaños et al., 2008). La variación estructural de las poblaciones forestales, depende en gran medida de la composición de especies de las comunidades, de tal forma que una misma especie puede presentar diferente condición estructural de acuerdo con la asociación florística donde se encuentra coexistiendo (Johnson et al., 2002).
La estructura horizontal de los bosques está dada por la abundancia, distribución y dominancia de las especies que lo conforman con respecto a la masa boscosa total, por lo que los parámetros más usados para medir la estructura son la densidad de árboles y el área basal (Rubio Camacho et al., 2014). Verticalmente se pueden distinguir tres estratos arbóreos: el estrato alto o superior, el estrato medio y el estrato bajo o inferior (Richards, 1966; Lamprecht, 1977; Pretzsch, 2009).
La información derivada de sitios permanentes (aquellas medidas sucesivas al menos tres veces) constituyen una fuente de información importante en manejo e investigación forestal para conocer los cambios respecto al crecimiento, producción y evolución de las masas forestales (Gadow et al., 1999; Kleinn y Morales, 2002). Una de las premisas del aprovechamiento sustentable es el mantenimiento de la composición e incremento del volumen maderable.
El objetivo de la investigación fue comparar la estructura (abundancia, área basal, área de copa, volumen e índice de Pretzsch), riqueza y complejidad biológica de especies arbóreas en un periodo de 10 años, medidas en intervalos de cinco años (2007, 2012, 2017) en un bosque de Durango, México.
Materiales y Métodos
El estudio se realizó en el ejido San Esteban y Anexos, municipio Pueblo Nuevo, ubicado en la parte suroeste del estado de Durango, en el macizo montañoso de la Sierra Madre Occidental (noroeste de México). Dicho ejido está enmarcado geográficamente entre los paralelos 23°43'46" y 23°55'17" de latitud Norte y los meridianos 105°22'23" y 105°29'59" de longitud Oeste (Fig. 1).
La altitud sobre el nivel del mar varía entre 2400 y 2600 m. Los tipos de suelo presentes son litosol, cambisol y regosol con textura predominantemente gruesa a media. El tipo de roca predominante es ígnea extrusiva ácida (INEGI, 2015). En esta zona prevalecen los climas templado subhúmedo con lluvias en verano y semifrío subhúmedo con lluvias en verano, los valores promedio anuales de precipitación y temperatura son 1200 mm y 18 ºC respectivamente (INEGI, 2009). Los tipos de vegetación predominantes corresponden a formaciones naturales de bosques de Pinus y Pinus-Quercus.
El manejo forestal que se lleva a cabo en este ejido contempla técnicas silvícolas del Método Mexicano de Ordenación de Bosques Irregulares (MMOBI) y del Método de Desarrollo Silvícola (MDS), realizando cortas de selección en sitios forestales con menor productividad y cortas más intensas, en calidades de estación altas y en sitios donde el riesgo de dañar los recursos asociados al bosque sea mínimo (UPSE 6, 2007).
Los datos dasométricos para el estudio se tomaron de ocho parcelas permanentes de muestreo, de forma cuadrangular de 2500 m2 (50 × 50 m). Las parcelas se establecieron en el 2007 siguiendo las metodologías desarrolladas por Corral-Rivas et al. (2009) y fueron evaluadas en intervalos de cinco años (2007, 2012 y 2017). A todos los árboles con diámetro a la altura del pecho ≥7.5 cm (DAP ≥7.5 cm, a una altura >1.3 m) conforme a los criterios de análisis de datos establecidos por Corral-Rivas et al. (2009), se les realizaron mediciones de DAP (cm), altura total (m), diámetro de la copa (mayor y menor, m). La identificación de cada una de las especies se realizó con base en la consulta de bibliografía especializada (García y González, 1998; González-Elizondo et al., 2007) y The Plant List (TPL, 2013), para confirmar el nombre científico correcto.
Análisis de datos
Para la caracterización de la estructura vertical de las especies se utilizó el índice de distribución vertical de especies (A) el cual es una modificación del índice Shannon (Pretzsch, 1996; Del Río et al., 2003). Donde A tiene valores entre cero y un valor máximo (A max ); cuando un valor A=0 significa que el rodal está constituido por una sola especie que ocurre en un solo estrato y A max se alcanza cuando la totalidad de las especies ocurren en la misma proporción tanto en el rodal como en los diferentes estratos y depende del número de especies y de estratos (Nagel, 1994; Pretzsch, 1996). Para la estimación de la distribución vertical de las especies, se definieron tres zonas de altura (Jiménez et al., 2001): zona I (alta): 80 a 100% de la altura máxima del área; zona II (media): 50 a 80% de la altura máxima, y zona III (baja): de cero a 50% de la altura máxima. El índice se estima mediante la fórmula:
donde S es el número de especies presentes, Z es el número de estratos de altura, p ij es el porcentaje de especies en cada zona, y se estima mediante la siguiente ecuación p ij =n i,j /N, donde n i,j es el número de individuos de la misma especie (i) en la zona (j) y N es el número total de individuos. El valor de A se estandariza de la siguiente forma:
Los valores de A rel cercanos a 100% indican que todas las especies se encuentran distribuidas de forma equitativa en los tres estratos de altura. En cada uno de los estratos de altura se caracterizó la estructura horizontal; se determinó su abundancia de acuerdo con el número de árboles, su dominancia en función del área basal, y su frecuencia con base en su presencia en los sitios de muestreo. Las variables relativizadas se utilizaron para obtener un valor ponderado a nivel de especie denominado índice de valor de importancia (IVI), que adquiere valores porcentuales en una escala de cero a 100 (Mueller y Ellenberg, 1974). Las fórmulas para estos cálculos se describen a continuación.
donde A i es la abundancia absoluta, AR i es la abundancia relativa de la especie i respecto a la abundancia total, N i es el número de individuos de la especie i, y E la superficie de muestreo (ha).
La dominancia (área basal) relativa se evaluó mediante la siguiente fórmula:
donde D i es la dominancia absoluta, DR i es dominancia relativa de la especie i respecto a la dominancia, Ab i es el área basal de la especie i y E la superficie (ha).
La frecuencia relativa se obtuvo con la siguiente ecuación:
donde F i es la frecuencia absoluta, FR i es la frecuencia relativa de la especie i respecto a la suma de las frecuencias, P i es el número de sitios en el que está presente la especie i y NS el número total de sitios de muestreo.
El índice de valor de importancia (IVI) se define como (Whittaker, 1972; Moreno, 2001):
También se determinó la cobertura total (área de copa) y por estrato en las tres mediciones:
donde C i es la cobertura absoluta y E la superficie (ha) y Ac i el área de copa de la especie i (Ac=π/4*d 2 ) (Romahn y Ramírez, 2010).
Para cada una de las especies se determinó el volumen rollo total árbol utilizando las ecuaciones de Schumacher y Hall sugeridas por Contreras y Návar (1997) las cuales se utilizan para estimar el volumen de las principales especies arbóreas en la Sierra Madre Occidental (Cuadro 1).
Cuadro 1: Ecuaciones para estimar volumen rollo total árbol de Schumacher y Hall (Contreras y Návar, 1997; 2002) para diferentes especies de pinos y latifoliadas de la Sierra Madre Occidental. V=Volumen (m3 ha-1); DAP=Diámetro a la altura del pecho (m); H=Altura total (m).
| Especie | Ecuación |
|---|---|
| Pinus cooperi C.E. Blanco | V=0.00004010*(DAP^1.9515)*(H0.9786) |
| Pinus durangensis Martínez | V=0.00001771*(DAP^2.1441)*(H1.0383) |
| Pinus engelmannii Carrière | V=0.00006498*(DAP^1.8561)*(H0.9701) |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | V=0.00006815*(DAP^1.8346)*(H0.9319) |
| Pinus teocote Schiede ex Schltdl. & Cham. | V=0.00035377*(DAP^1.4896)*(H0.8052) |
| Otras coníferas (Pinus sp. y Juniperus sp.) | V=0.00024698*(DAP^1.6254)*(H0.8550) |
| Otras hojosas (Quercus sp. y Arbutus sp.) | V=0.00024732*(DAP^1.6169)*(H1.0941) |
La diversidad alfa (α) se determinó con la riqueza de especies (S) y con el índice de Shannon-Wiener (H’), el cual es una medida legítima de la complejidad de las comunidades biológicas (Moreno, 2001; Leyva-López, et al., 2010; Jost y González-Oreja, 2012). Para determinar la existencia de diferencias estadísticamente significativas entre mediciones se realizó un análisis de varianza (ANOVA) de un factor a cada parámetro dasométrico, primero se constató que los datos cumplieran con los supuestos de normalidad, para lo cual se realizó la prueba de Shapiro-Wilk, mientras que el de homocedasticidad mediante la prueba de Levene. En caso de diferencias significativas se usaron las pruebas de comparaciones múltiples de Tukey. El nivel de significancia para todas las pruebas fue α=0.05. Los análisis estadísticos se llevaron a cabo con el programa SPSS 22.0 para Windows (IBM Corp. Released, 2013).
Resultados
En total se registraron 16 especies distribuidas en cuatro géneros y cuatro familias. Las familias Pinaceae y Fagaceae fueron la más representadas con seis especies cada una. En total estas dos familias integraron 12 especies (Cuadro 2).
Cuadro 2: Nombre científico y común, y familia a la que pertenecen las especies arbóreas registradas en el ejido San Esteban y Anexos, Pueblo Nuevo, Durango, México.
| Nombre científico | Nombre común | Familia |
|---|---|---|
| Arbutus arizonica (A. Gray) Sarg. | madroño liso | Ericaceae |
| Arbutus madrensis S. González | madroño roñoso | Ericaceae |
| Arbutus tessellata P.D. Sørensen | madroño pegajoso | Ericaceae |
| Juniperus deppeana Steud. | táscate | Cupressaceae |
| Pinus cooperi C.E.Blanco | pino chino | Pinaceae |
| Pinus durangensis Martínez | ocote | Pinaceae |
| Pinus engelmannii Carrière | pino real | Pinaceae |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | pino prieto | Pinaceae |
| Pinus strobiformis Engelm. | pino blanco | Pinaceae |
| Pinus teocote Schied. ex Schltdl. & Cham. | pino colorado | Pinaceae |
| Quercus arizonica Sarg. | encino | Fagaceae |
| Quercus conzattii Trel. | encino negro | Fagaceae |
| Quercus crassifolia Bonpl. | encino prieto | Fagaceae |
| Quercus obtusata Bonpl. | encino roble | Fagaceae |
| Quercus rugosa Née | encino blanco | Fagaceae |
| Quercus sideroxyla Bonpl. | encino colorado | Fagaceae |
Índice de distribución vertical
Por el índice de distribución vertical (A) se definieron tres estratos de altura (I, II y III) para cada año de medición, tomando en cuenta la altura mayor de cada medición, la cual aumentó en promedio (± desviación estándar) 1.5 ± 0.07 m cada cinco años. Para la primera medición (2007) el estrato I (19.9-24.9 m) incluyó 41 árboles ha-1 distribuidos en seis especies, en el II (12.5-19.8 m) se registraron 169 árboles ha-1 distribuidos en 14 especies, mientras que el III (<12.4 m) se conformó por 505 árboles ha-1 distribuidos en las 16 especies totales.
Por otro lado, en la segunda medición (2012) en el estrato I (21.1-26.4 m) disminuyó el número de individuos a 25 árboles ha-1 en cuatro especies, en el II (13.2-21.0 m) el número de individuos aumentó a 177 árboles ha-1 distribuidos en las mismas 14 especies y finalmente en el III (<13.1 m) el número de individuos disminuyó a 484 árboles ha-1 distribuidos en las 16 especies.
En la última medición (2017) el estrato I (22.2-27.8 m) disminuyó en número de individuos respecto a las mediciones anteriores, pues se registraron 18 árboles ha-1, pero aumentó el número de especies a cinco; en el II (13.9-22.1 m) el número de individuos disminuyó a 171 árboles ha-1 distribuidos en 10 especies, que también disminuyeron respecto a la medición anterior y en el III (2.3-13.8 m) el número de individuos aumentó a 525 árboles ha-1 distribuidos en las 16 especies originales.
Las especies Pinus cooperi C.E. Blanco, Pinus durangensis Martínez, Pinus teocote Schied. ex Schltdl. & Cham. y Quercus sideroxyla Bonpl., estuvieron presentes en todos los estratos de altura en las tres mediciones, donde su proporción de individuos fue mayor en el III y disminuyó en el I (Cuadro 3).
Cuadro 3: Número de individuos (N ha-1), por especie y por estrato de acuerdo con el índice vertical de Pretzsch (A) para los años 2007, 2012 y 2017 en ocho parcelas de 2500 m2 establecidas en el ejido San Esteban y Anexos, Pueblo Nuevo, Durango, México. El estrato I (alto): 80 a 100% de la altura máxima del área; II (medio): 50 a 80% de la altura máxima, III (bajo): de cero a 50% de la altura máxima.
| Especie | 2007 | 2012 | 2017 | |||
|---|---|---|---|---|---|---|
| N ha-1 | (%) | N ha-1 | (%) | N ha-1 | (%) | |
| Estrato I | (19.9-24.9 m) | (21.1-26.4 m) | (22.2-27.8 m) | |||
| Pinus cooperi C.E. Blanco | 5 | 0.7 | 4 | 0.6 | 4 | 0.5 |
| Pinus durangensis Martínez | 25 | 3.5 | 16 | 2.3 | 11 | 1.5 |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | 1 | 0.1 | - | - | - | - |
| Pinus strobiformis Engelm. | 1 | 0.1 | - | - | 1 | 0.1 |
| Pinus teocote Schied. ex Schltdl. & Cham. | 6 | 0.8 | 3 | 0.4 | 2 | 0.3 |
| Quercus sideroxyla Bonpl. | 3 | 0.4 | 2 | 0.2 | 1 | 0.1 |
| Suma | 41 | 5.7 | 25 | 3.6 | 18 | 2.5 |
| Estrato II | (12.5-19.8) | (13.2-21.0 m) | (13.9-22.1 m) | |||
| Arbutus madrensis S. González | 1 | 0.1 | 1 | 0.1 | - | - |
| Arbutus tessellata P.D. Sørensen | 1 | 0.1 | 1 | 0.1 | - | - |
| Juniperus deppeana Steud. | 1 | 0.1 | 1 | 0.1 | 2 | 0.2 |
| Pinus cooperi C.E.Blanco | 14 | 1.9 | 16 | 2.3 | 14 | 1.9 |
| Pinus durangensis Martínez | 72 | 10.1 | 75 | 10.9 | 77 | 10.8 |
| Pinus engelmannii Carrière | 3 | 0.4 | 3 | 0.4 | 3 | 0.4 |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | 9 | 1.2 | 6 | 0.9 | 6 | 0.8 |
| Pinus strobiformis Engelm. | 13 | 1.8 | 17 | 2.4 | 17 | 2.3 |
| Pinus teocote Schied. ex Schltdl. & Cham. | 22 | 3.0 | 27 | 3.9 | 26 | 3.6 |
| Quercus arizonica Sarg. | 1 | 0.1 | 1 | 0.1 | 1 | 0.1 |
| Quercus conzattii Trel. | 3 | 0.3 | 1 | 0.1 | 1 | 0.1 |
| Quercus crassifolia Bonpl. | 1 | 0.1 | 1 | 0.1 | - | - |
| Quercus rugosa Née | 1 | 0.1 | 1 | 0.1 | - | - |
| Quercus sideroxyla Bonpl. | 31 | 4.3 | 29 | 4.2 | 27 | 3.8 |
| Suma | 169 | 23.7 | 177 | 25.8 | 171 | 24.0 |
| Estrato III | (2.0-12.4 m) | (2.5-13.1 m) | (2.3-13.8 m) | |||
| Arbutus arizonica (A. Gray) Sarg. | 10 | 1.3 | 5 | 0.7 | 5 | 0.7 |
| Arbutus madrensis S. González | 28 | 3.9 | 27 | 3.9 | 29 | 4.1 |
| Arbutus tessellata P.D. Sørensen | 12 | 1.6 | 12 | 1.7 | 13 | 1.8 |
| Juniperus deppeana Steud. | 20 | 2.8 | 19 | 2.8 | 26 | 3.6 |
| Pinus cooperi C.E.Blanco | 7 | 1.0 | 5 | 0.7 | 7 | 0.9 |
| Pinus durangensis Martínez | 98 | 13.6 | 100 | 14.6 | 103 | 14.4 |
| Pinus engelmannii Carrière | 4 | 0.6 | 4 | 0.6 | 5 | 0.6 |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | 25 | 3.4 | 24 | 3.4 | 26 | 3.6 |
| Pinus strobiformis Engelm. | 35 | 4.9 | 37 | 5.4 | 50 | 7.0 |
| Pinus teocote Schied. ex Schltdl. & Cham. | 25 | 3.5 | 22 | 3.2 | 27 | 3.7 |
| Quercus arizonica Sarg. | 8 | 1.1 | 8 | 1.2 | 9 | 1.2 |
| Quercus crassifolia Bonpl. | 13 | 1.8 | 13 | 1.9 | 15 | 2.1 |
| Quercus conzattii Trel. | 39 | 5.5 | 41 | 5.9 | 42 | 5.9 |
| Quercus obtusata Bonpl. | 1 | 0.1 | 1 | 0.1 | 1 | 0.1 |
| Quercus rugosa Née | 3 | 0.3 | 3 | 0.4 | 5 | 0.6 |
| Quercus sideroxyla Bonpl. | 180 | 25.2 | 166 | 24.2 | 166 | 23.3 |
| Suma | 505 | 70.7 | 484 | 70.7 | 525 | 73.5 |
| Suma total | 715 | 100.0 | 685 | 100.0 | 714 | 100.0 |
El valor absoluto del índice de Pretzsch (A) para el año 2007 fue de A=2.72 con A max =3.87 y A rel =70%, para los siguientes años estos valores presentaron una disminución mínima, para el año 2012 fueron de A=2.67 con A max =3.87 y A rel =69%, mientras que para el 2017 los valores fueron de A=2.66 con A max =3.87 y A rel =68%.
Estructura de la vegetación
La abundancia total no presentó cambios significativos del año 2007 al 2017 (F 2,21=0.051, P=0.950), en 2007 el promedio (± error estándar) fue 715±74 árboles ha-1, disminuyendo levemente en 2012 a 685±71 árboles ha-1, mientras que en 2017 el valor fue 714±77 árboles ha-1. Esta abundancia disminuyó ligeramente en estrato I de 2007 (54±15 árboles ha-1) a 2017 (36±5 árboles ha-1), P. durangensis fue la especie de mayor abundancia en 2007 (25 árboles ha-1), la cual disminuyó 56% en la última medición (11 árboles ha-1). En el II fue constante de 2007 (169±26 árboles ha-1) a 2017 (171±40 árboles ha-1), P. durangensis presentó el mayor valor (72 árboles ha-1) sin presentar cambios considerables a través del tiempo. El III presentó un aumento de 2007 (505±84 árboles ha-1) a 2017 (525±75 árboles ha-1), Q. sideroxyla fue la especie más abundante (180 árboles ha-1), disminuyendo su presencia en 8% en las siguientes mediciones (166 árboles ha-1) (Fig. 2A).
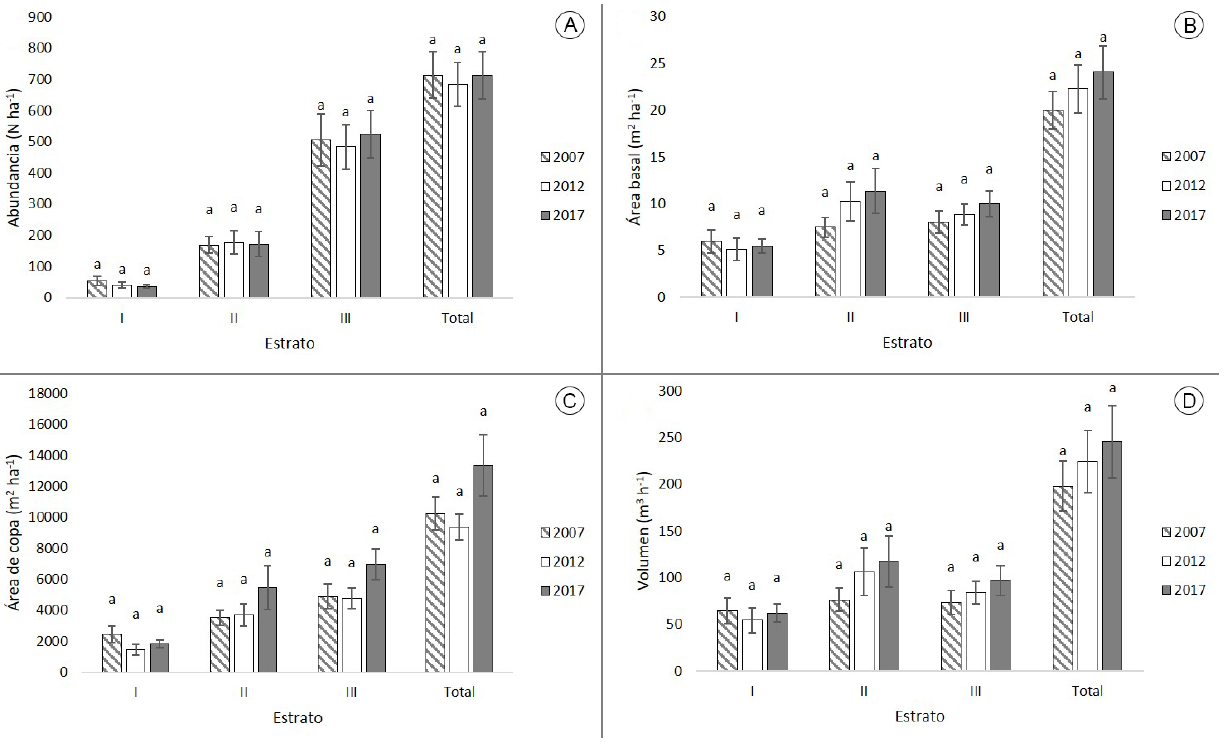
Figura 2: Valores por estratos y totales de los parámetros estructurales en el ejido San Esteban y Anexos, Pueblo Nuevo, Durango, México. A. número de árboles por ha (N ha-1); B. área basal (m2 ha-1); C. área de copa (m2 ha-1); D. volumen total árbol (m3 ha-1). Valores promedios ± error estándar. Letras diferentes (a, b) indican diferencia significativa entre años (p<0.05).
El área basal total (dominancia) del arbolado no mostró cambios significativos del año 2007 a 2017 (F 2,21=0.670, P=0.522). En el 2007 el promedio (± error estándar) fue de 19.99±2.00 m2 ha-1 y aumentó en las siguientes mediciones hasta alcanzar 24.07±2.86 m2 ha-1 en 2017. El estrato I mantuvo un valor constante (5.97±1.27 m2 ha-1 en 2007 y 5.47±0.77 m2 ha-1 en 2017), la mayor dominancia fue la de P. durangensis en 2007 (2.24 m2 ha-1), que disminuyó en 2017 (1.49 m2 ha-1). El II aumentó de 2007 (7.48±1.06) a 2017 (11.35±2.39 m2 ha-1), la especie P. durangensis presentó mayor dominancia; sin embargo, aumentó conforme pasó el tiempo (2.49 a 4.27 m2 ha-1). El III se incrementó de 2007 (8.03±1.18 m2 ha-1) a 2017 (9.98±1.36 m2 ha-1), Q. sideroxyla fue la especie más dominante, aumentando su área basal en el período (3.25 a 3.65 m2 ha-1) (Fig. 2B).
El área de copa total del arbolado no presentó cambios significativos de 2007 a 2017 (F 2,21 =2.238, P=0.131), el promedio (± error estándar) para 2007 fue de 10251±1079 m2 ha-1, mientras que para el 2012 disminuyó a 9388±830 m2 ha-1 y para 2017 aumentó a 13333±1978 m2 ha-1. Los estratos no presentaron cambios significativos. En el estrato I el promedio de 2007 fue de 2443±544 m2 ha-1, el cual disminuyó en 2017 donde registró un valor de 1816±242 m2 ha-1. En el estrato II el promedio para 2007 fue de 3523±491 m2 ha-1, el cual aumentó considerablemente hasta 5456±1404 en 2017. Para el estrato III se presentó un promedio para 2007 de 4895±797, mientras que para 2017 aumentó considerablemente a 6968±995 m2 ha-1 (Fig. 2C).
El volumen total del arbolado presentó un aumento, pero no significativo del año 2007 a 2017 (F 2,21 =0.517, P=0.604), para 2007 el promedio (± error estándar) fue de 198±27 m3 ha-1, el cual aumentó en 2017 a 246±39 m3 ha-1. Los estratos no mostraron cambios significativos. El promedio para el estrato I en 2007 fue de 64±14 m3 ha-1, el cual disminuyó levemente a 62±10 m3 ha-1 en 2017. En el estrato II el promedio para 2007 fue de 76±13 m3 ha-1 aumentando considerablemente en 2017 a 117±28 m3 ha-1. Para el estrato III el promedio para 2007 fue 73±13 m3 ha-1 aumentando a 97±17 m3 ha-1 en 2017 (Fig. 2D).
Índice de valor de importancia (IVI)
Las especies con mayor abundancia en los tres estratos (I, II y III) de los tres años de medición (2007, 2012 y 2017) fueron casi las mismas. En el estrato I resultó ser P. durangensis (62.7, 65.3 y 61.1%) seguida de P. teocote para el año 2007 (14.8%) y P. cooperi en los años 2012 y 2017 (16.3 y 19.4%). Situación similar se presentó en el II donde P. durangensis fue la más abundante en las tres mediciones (42.6, 42.5 y 45%), pero seguida por Q. sideroxyla en los tres años (18.3, 16.4 y 15.8%). Q. sideroxyla fue la especie con mayor porcentaje de individuos en las tres evaluaciones del III (35.6, 34.3 y 31.6%).
Los tres estratos de cada año estuvieron dominados por las mismas especies. Pinus durangensis fue la más dominante en el I en los tres años (50, 60.2 y 54.7%), seguida por Q. sideroxyla. En el II también P. durangensis dominó en todas las evaluaciones (33.4, 33.6 y 37.6%), seguida de Q. sideroxyla (29, 30, 26.9%). Esta última especie fue la más dominante en el III (40.4, 39.3 y 36.6%) seguida de P. durangensis (14.3, 15.8 y 14.8%).
No todos los estratos tuvieron la misma frecuencia de especies. La especie Pinus durangensis fue la más frecuente en el estrato I en todos los años (26.3, 41.7, 30%). Mismo caso para el II donde P. durangensis presentó la mayor frecuencia en las tres mediciones (17.8, 18.2 y 21.1%). Por otro lado, Q. sideroxyla fue la más frecuente en el III (12.1, 12.7 y 13.6%), en todas las evaluaciones.
De acuerdo con los resultados anteriores P. durangensis resultó ser la especie con mayor peso ecológico en las tres mediciones de los estratos I (46, 55.7 48.6%) y II (31.3, 31.4 y 34.6%). Mientras que para el III fue Q. sideroxyla (29.4, 28.8 y 27.3%) (Cuadro 4).
Cuadro 4: Parámetros estructurales estimados para las especies registradas por estrato en cada año de medición (2007, 2012 y 2017) en el ejido San Esteban y Anexos, Pueblo Nuevo, Durango. Ar=Abundancia relativa; Dr=Dominancia relativa (área basal); Fr=Frecuencia relativa; IVI=Índice de valor de importancia.
| Especie | 2007 | 2012 | 2017 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Ar (%) |
Dr (%) |
Fr (%) |
IVI (%) |
Ar (%) |
Dr (%) |
Fr (%) |
IVI (%) |
Ar (%) |
Dr (%) |
Fr (%) |
IVI (%) |
|
| Estrato I | ||||||||||||
| Pinus cooperi C.E. Blanco | 12.3 | 11.8 | 10.5 | 11.6 | 16.3 | 14.7 | 16.7 | 15.9 | 19.4 | 16 | 20 | 18.5 |
| Pinus durangensis Martínez | 61.7 | 50 | 26.3 | 46 | 65.3 | 60.2 | 41.7 | 55.7 | 61.1 | 54.7 | 30 | 48.6 |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | 1.2 | 0.7 | 5.3 | 2.4 | - | - | - | - | - | - | - | - |
| Pinus strobiformis Engelm. | 2.5 | 1.3 | 10.5 | 4.8 | - | - | - | - | 2.8 | 1.5 | 10 | 4.8 |
| Pinus teocote Schied. ex Schltdl. & Cham. | 14.8 | 15.9 | 26.3 | 19 | 12.2 | 15.5 | 25 | 17.6 | 11.1 | 14.2 | 20 | 15.1 |
| Quercus sideroxyla Bonpl. | 7.4 | 20.3 | 21.1 | 16.3 | 6.1 | 9.7 | 16.7 | 10.8 | 5.6 | 13.6 | 20 | 13 |
| Suma | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Estrato II | ||||||||||||
| Arbutus madrensis S. González | 0.3 | 0.5 | 2.2 | 1 | 0.6 | 0.5 | 4.5 | 1.9 | - | - | - | - |
| Arbutus tessellata P.D. Sørensen | 0.3 | 1 | 2.2 | 1.2 | 0.3 | 0.8 | 2.3 | 1.1 | - | - | - | - |
| Juniperus deppeana Steud. | 0.6 | 1.3 | 2.2 | 1.4 | 0.6 | 1.1 | 2.3 | 1.3 | 0.9 | 1.5 | 2.6 | 1.7 |
| Pinus cooperi C.E.Blanco | 8 | 6.7 | 8.9 | 7.9 | 8.8 | 6.5 | 9.1 | 8.1 | 7.9 | 6.3 | 5.3 | 6.5 |
| Pinus durangensis Martínez | 42.6 | 33.4 | 17.8 | 31.3 | 42.5 | 33.6 | 18.2 | 31.4 | 45 | 37.6 | 21.1 | 34.6 |
| Pinus engelmannii Carrière | 1.8 | 1.4 | 4.4 | 2.5 | 1.7 | 1.2 | 4.5 | 2.5 | 1.5 | 1 | 5.3 | 2.6 |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | 5 | 4.9 | 11.1 | 7 | 3.4 | 3.5 | 11.4 | 6.1 | 3.5 | 3.6 | 10.5 | 5.9 |
| Pinus strobiformis Engelm. | 7.7 | 7.1 | 8.9 | 7.9 | 9.3 | 7.2 | 9.1 | 8.5 | 9.6 | 7.3 | 13.2 | 10 |
| Pinus teocote Schied. ex Schltdl. & Cham. | 12.7 | 11.4 | 15.6 | 13.2 | 15 | 14.1 | 15.9 | 15 | 15.2 | 15.2 | 18.4 | 16.3 |
| Quercus arizonica Sarg. | 0.3 | 0.5 | 2.2 | 1 | 0.3 | 0.4 | 2.3 | 1 | 0.3 | 0.4 | 2.6 | 1.1 |
| Quercus conzattii Trel. | 1.5 | 1.2 | 2.2 | 1.6 | 0.6 | 0.4 | 2.3 | 1.1 | 0.3 | 0.2 | 2.6 | 1 |
| Quercus crassifolia Bonpl. | 0.6 | 0.6 | 2.2 | 1.2 | 0.3 | 0.2 | 2.3 | 0.9 | - | - | - | - |
| Quercus rugosa Née | 0.3 | 0.9 | 2.2 | 1.1 | 0.3 | 0.6 | 2.3 | 1.1 | - | - | - | - |
| Quercus sideroxyla Bonpl. | 18.3 | 29 | 17.8 | 21.7 | 16.4 | 30 | 13.6 | 20 | 15.8 | 26.9 | 18.4 | 20.4 |
| Suma | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Estrato III | ||||||||||||
| Arbutus arizonica (A. Gray) Sarg. | 1.9 | 3.5 | 4.5 | 3.3 | 1 | 2.6 | 4.8 | 2.8 | 1 | 1.4 | 1.7 | 1.3 |
| Arbutus madrensis S. González | 5.5 | 4.1 | 9.1 | 6.3 | 5.5 | 3.8 | 9.5 | 6.3 | 5.5 | 4.4 | 10.2 | 6.7 |
| Arbutus tessellata P.D. Sørensen | 2.3 | 1.4 | 4.5 | 2.8 | 2.4 | 1.6 | 4.8 | 2.9 | 2.5 | 2.6 | 5.1 | 3.4 |
| Juniperus deppeana Steud. | 4 | 4.5 | 10.6 | 6.4 | 3.9 | 4.5 | 11.1 | 6.5 | 4.9 | 4.7 | 11.9 | 7.1 |
| Pinus cooperi C.E.Blanco | 1.4 | 1.7 | 3 | 2 | 1 | 1 | 1.6 | 1.2 | 1.2 | 1.5 | 1.7 | 1.5 |
| Pinus durangensis Martínez | 19.3 | 14.3 | 12.1 | 15.2 | 20.7 | 15.8 | 11.1 | 15.9 | 19.5 | 14.8 | 11.9 | 15.4 |
| Pinus engelmannii Carrière | 0.8 | 0.6 | 4.5 | 2 | 0.8 | 0.7 | 4.8 | 2.1 | 0.9 | 1.1 | 3.4 | 1.8 |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | 4.9 | 5.3 | 7.6 | 5.9 | 4.9 | 5.1 | 7.9 | 6 | 4.9 | 5.1 | 6.8 | 5.6 |
| Pinus strobiformis Engelm. | 6.9 | 5.3 | 10.6 | 7.6 | 7.6 | 5.3 | 11.1 | 8 | 9.5 | 6.5 | 10.2 | 8.7 |
| Pinus teocote Schied. ex Schltdl. & Cham. | 5 | 4.5 | 10.6 | 6.7 | 4.5 | 4 | 11.1 | 6.5 | 5.1 | 4.8 | 11.9 | 7.2 |
| Quercus arizonica Sarg. | 1.6 | 1.7 | 1.5 | 1.6 | 1.7 | 1.9 | 1.6 | 1.7 | 1.6 | 1.9 | 1.7 | 1.8 |
| Quercus conzattii Trel. | 7.7 | 7.6 | 3 | 6.1 | 8.4 | 8.5 | 1.6 | 6.2 | 8 | 8.4 | 3.4 | 6.6 |
| Quercus crassifolia Bonpl. | 2.6 | 4.5 | 1.5 | 2.9 | 2.7 | 5.1 | 1.6 | 3.1 | 2.9 | 4.8 | 1.7 | 3.1 |
| Quercus obtusata Bonpl. | 0.1 | 0.2 | 1.5 | 0.6 | 0.1 | 0.2 | 1.6 | 0.6 | 0.1 | 0.2 | 1.7 | 0.7 |
| Quercus rugosa Née | 0.5 | 0.4 | 3 | 1.3 | 0.5 | 0.4 | 3.2 | 1.4 | 0.9 | 1.3 | 3.4 | 1.8 |
| Quercus sideroxyla Bonpl. | 35.6 | 40.4 | 12.1 | 29.4 | 34.3 | 39.3 | 12.7 | 28.8 | 31.6 | 36.6 | 13.6 | 27.3 |
| Suma | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Suma general | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 |
Distribución por clases diamétricas
El número de árboles ha-1 por clase diamétrica, en cada estrato fue diferente. El III registró la mayor cantidad de individuos, mostrando un comportamiento decreciente (J invertida) en las tres mediciones, presentándose mayor abundancia de árboles en las clases diamétricas menores y decreciendo conforme aumentan estas. El estrato II tiene un comportamiento similar, pero a menor escala, mientras que en el I la menor abundancia de árboles ha-1 se encuentra en las categorías mayores (Fig. 3).
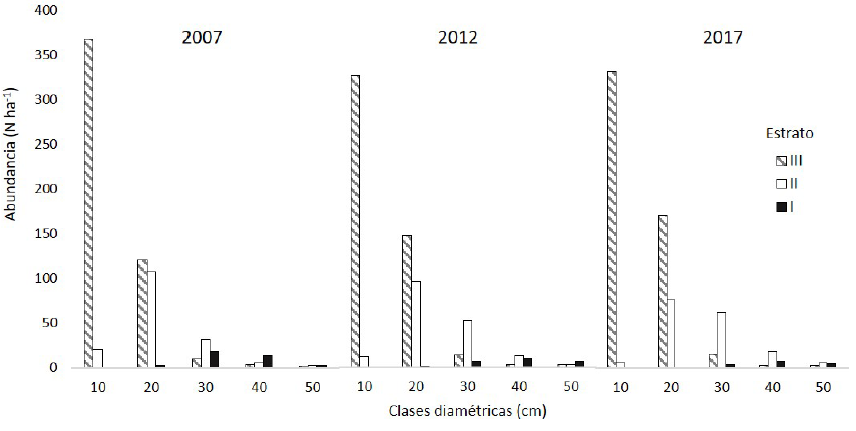
Diversidad alfa (α)
La riqueza de especies total (S) no presentó cambios significativos de 2007 a 2017 (F 2,21 =1.054, P=0.366), el promedio (± error estándar) para 2007 fue de 8.8±0.45 especies, el cual disminuyó a 7.88±0.52 especies en 2017. Los estratos no presentaron cambios. En I en 2007 el promedio fue de 2.38±0.60 especies disminuyendo a 1.25±0.49 en 2017. En el estrato II el valor de 2007 fue de 5.63±0.32 especies, valor que disminuyó a 4.75±0.25 en 2017. Para el estrato III el promedio para 2007 fue de 8.25±0.53 especies, disminuyendo a 7.38±0.60 en 2017 (Fig. 4A).
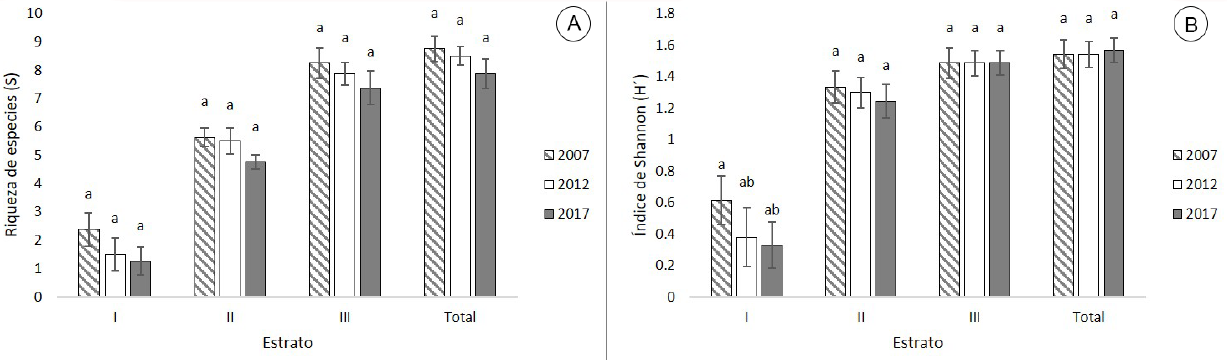
Figura 4: Valores de la diversidad alfa (α) en el ejido San Esteban y Anexos, Pueblo Nuevo, Durango, México. A. riqueza de especies por estrato y total; B. índice de Shannon-Wiener por estrato y total. Valores promedios ± error estándar. Letras diferentes (a, b,) indican diferencia significativa entre años (p<0.05).
La complejidad biológica total medida a través del índice de Shannon-Wiener (H´) no presentó cambios significativos de 2007 a 2017 (F 2,21 =0.028, P=0.973), el valor promedio para 2007 fue de 1.54±0.09, mientras que para 2017 fue muy similar (1.57±0.08). Los estratos no presentaron cambios significativos. El valor del estrato I para 2007 fue de 0.61±0.16, disminuyendo en las siguientes mediciones hasta registrar un valor de 0.33±0.15 en 2017. El promedio para el estrato II para 2007 fue de 1.33±0.10, disminuyendo a 1.24±0.11 en 2017. Para el estrato III en el 2007 el valor promedio fue de 1.48±0.10 manteniéndose similar en las siguientes mediciones (1.48±0.08) (Fig. 4B).
Discusión
La estructura general del bosque templado estudiado se caracteriza por el dominio de las familias Pinaceae y Fagaceae, que han sido registradas como las de mayor representatividad en los bosques templados de México (Aragón-Piña et al., 2010; López-Hernández et al., 2017). La familia Pinaceae y el género Pinus son de amplia distribución en todas las cadenas montañosas del país (García y González 1998; Hernández-Salas et al., 2013).
Según Lamprecht (1986), el análisis de especies arbóreas se debe realizar mediante el estudio de los estratos en las copas. La altura máxima que alcanzaron los individuos localizados en el área de muestreo fue diferente en cada año de medición, aumentando de 24.9 m en 2007 a 27.8 m en 2017, a partir de las cuales se determinaron los tres estratos del índice A (Aguirre, 2002; Pretzsch, 2009). Los valores del índice A fueron muy similares en las tres mediciones (A=2.66 con A max =3.87 y A rel =68%). Lo que indica que la zona evaluada tiene una alta uniformidad y no presentó cambios considerables en la distribución de especies en los estratos de altura en el tiempo, siendo el estrato III el que presenta la mayor heterogeneidad. Estos valores coinciden con Graciano-Ávila et al. (2017) quienes registraron valores de A=2.07 y A max =3.58 al caracterizar un bosque de la misma región y con Jiménez et al. (2001) que registraron valores similares (A=2.07; A max =3.50) al analizar la estructura horizontal y vertical en un ecosistema multicohortal de pino-encino en el norte de México.
En la densidad, los cambios más notorios fueron por la muerte, tala e incorporación de individuos en las mediciones del 2012 y 2017, aun así, no se registraron cambios significativos, ya que el número de individuos fue constante a través del tiempo. Los estratos I y II se encuentran dominados por P. durangensis en las tres mediciones, dicha especie se caracteriza por ser una de las que alcanza mayor altura en los bosques templados de México (hasta 42 m) (González-Elizondo et al., 2012). Además, ha sido registrada en diversos estudios como la especie más abundante en los bosques templados de Durango (Noroeste de México) (Valenzuela-Núñez y Granados-Sánchez, 2009). En el estrato III existe mayor presencia de Q. sideroxyla en las tres mediciones, esta especie junto con P. durangensis y P. teocote es común que se encuentren asociadas entre sí (García y González, 1998; Márquez-Linares et al., 1999) y han sido documentadas como las más frecuentes en los bosques templados (Silva-Flores et al., 2014). Jiménez et al. (2001) al analizar la estructura de un bosque en el norte de México, encontraron que las distintas especies de Quercus se ubicaron de manera mínima en el estrato II y en mayor proporción en el estrato III; coincidiendo con Villavicencio et al. (2005), que señalan que las especies del género Quercus son las dominantes en el estrato bajo. Cabe señalar que todas las especies arbóreas registradas en el área de estudio tuvieron presencia en el estrato inferior (III).
El área basal total aumentó en las tres mediciones; sin embargo, el cambio no fue significativo, el estrato bajo (III) fue el de mayor aumento. Uno de los mejores indicadores para medir el grado de recuperación del bosque es el área basal (Norden et al., 2009). Diversos autores señalan que el área basal de los bosques secundarios incrementa de forma asintótica a través del tiempo y tiende a ser semejante a los bosques primarios (Guariguata y Ostertag, 2002; Ruschel et al., 2009).
No se presentaron cambios significativos en composición de especies en el área de estudio en las tres mediciones, la especie P. durangensis resultó ser la especie de mayor importancia ecológica; sin embargo, al analizar el IVI por estratos, resultó que Q. sideroxyla fue la de mayor relevancia en el estrato III. Estos resultados concuerdan con Delgado et al. (2016) quienes registraron a P. durangensis y Q. sideroxyla como especies de mayor valor ecológico en bosques de la región aledaños al área de estudio.
Las copas de los árboles compiten por recursos, especialmente con individuos vecinos, siendo la competencia por espacio de crecimiento y radiación, algunos de los factores más importantes que determinan el adecuado crecimiento y buen desarrollo de un árbol (Saavedra-Romero et al., 2016). El área de copa total no presentó cambios significativos entre los diferentes años de medición, pero esta fue mayor a 10,000 m2 ha-1; es decir superior a 100% de la superficie muestreada, lo que significa una sobreposición de copas. El estrato III presentó el mayor aporte y aumento en las mediciones. López-Hernández et al. (2017) registraron un valor menor (8463 m2 ha-1) en un bosque templado del centro de México.
El área de estudio presentó un aumento en volumen que no fue significativo (197.89 a 245.57 m3 ha-1), siendo los estratos II y III los que presentaron el mayor incremento; en éstos se concentraron el mayor número de individuos de Pinus y Quercus. Autores como Návar-Cháidez y González-Elizondo (2009), señalan que el volumen aumenta con el índice de mezcla de Pinus y Quercus y con el incremento en la densidad en bosques de Durango, México. Corral-Rivas et al. (2015) registraron valores menores (181.7 m3 ha-1) en bosques mixtos e irregulares de Durango, México.
La distribución de clases diamétricas presentó una forma de J invertida en las tres mediciones, encontrándose gran cantidad de individuos en las categorías diamétricas menores que disminuyen en los mayores diámetros, dichos individuos jóvenes reemplazarán a árboles maduros en un futuro (Arruda et al., 2011; Hernández-Stefanoni et al., 2011). Este tipo de distribuciones diamétricas es la mejor garantía de la supervivencia de la comunidad forestal, ya que los individuos de tallas superiores son eliminados ocasionalmente y sustituidos sin dificultad por individuos de las categorías diamétricas inferiores (Lamprecht, 1990). La desigualdad en los tamaños diamétricos es un efecto asociado a procesos competitivos que llevan a cabo las especies durante las diferentes etapas de desarrollo del rodal (Olvera-Vargas y Figueroa-Rangel, 2012).
La riqueza total de especies en las tres mediciones no presentó cambios significativos (S=8 en promedio en cada sitio). El estrato I fue el que tuvo menor número, mientras que el III, al agrupar mayor número de individuos, permitió que el número de especies aumentara. Condit et al. (1996), mencionan que las especies se acumulan en función del número de individuos muestreados. Los valores encontrados en este estudio fueron mayores a las que López-Hernández et al. (2017) registraron en un bosque de Puebla, México (11).
Por otro lado, la complejidad biológica en el área de estudio no presentó cambios significativos (H´=1.57), esta disminuyó ligeramente en los estratos I y II a través del tiempo, mientras que en el III se presentó el mayor valor que fue constante en las tres mediciones. López-Hernández (2017) registraron valores menores (H´=1.37) a lo registrado en este estudio. Por otro lado, se han reportado valores de H´=1.51 y H´=1.87 en Áreas Naturales Protegidas del estado de Durango (Vargas Larreta, 2013; Domínguez et al., 2018). En bosques con poca perturbación y mayor edad, generalmente se presenta alta diversidad de árboles y especies arbustivas (Onaindia et al., 2004).
Conclusiones
La comparación realizada permitió determinar que la vegetación arbórea de la comunidad evaluada se mantiene sin cambios respecto a la diversidad de especies, mientras que el área basal y el volumen tienden a aumentar. La composición de especies en el área estudiada fue muy similar en las tres mediciones, se registraron 16 pertenecientes principalmente a las familias Pinaceae y Fagaceae, siendo el género Pinus el de mayor representatividad. La altura máxima aumentó en promedio 1.5 m cada 5 años. De acuerdo con los valores del índice vertical (A) existe alta uniformidad, no hay cambios considerables en la distribución de especies en los tres estratos de altura a través del tiempo, y el estrato III tiene mayor heterogeneidad. Las especies con mayor contribución estructural son P. durangensis y Q. sideroxyla, la importancia de la primera recae en los estratos I y II, mientras que la segunda presenta mayor relevancia en el III. La cobertura fue mayor a 100% y aumentó conforme pasó el tiempo, indicando una sobreposición de copas, presentando la mayor contribución el estrato III. Los estratos II y III presentaron el mayor aumento en volumen, reflejo de la densidad existente en estos.
La distribución de clases diamétricas presentó una forma de tipo J invertida, donde el mayor número de individuos se localiza en el estrato III, lo que garantiza la supervivencia de esta comunidad forestal. Los valores de riqueza y complejidad biológica no mostraron cambios significativos, pero tienden a disminuir con el paso del tiempo, aun así, son altos en comparación con otros bosques similares. Las especies registradas se distribuyeron de diferente forma en los estratos, siendo el estrato III el más diverso.
Lo obtenido en esta investigación sirve como referencia para el desarrollo de planes de manejo silvícola, aportando información respecto a los cambios existentes en el bosque y sus estratos a través del tiempo.

