











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Las plantas invasoras son una amenaza para los ecosistemas nativos, debido a que provocan la pérdida de la riqueza del suelo, hidrología, cambios en los ciclos de nutrientes, aumento en la duración e intensidad de los ciclos de fuego y ocasionalmente la pérdida de la biodiversidad nativa (Vanderhoeven et al., 2005; Ng'weno et al., 2009; Murray y Philips, 2010). Se considera que las especies invasoras son un factor importante en el cambio de los ecosistemas y la segunda amenaza más grave después de la fragmentación de los hábitats (Dukes y Mooney, 1999; Rai et al., 2012). El conocimiento de cómo crecen y se desarrollan en los diferentes tipos de vegetación es información básica para su control y/o manejo (Williams y Baruch, 2000; Arriaga et al., 2004).
El pasto [Melinis repens (Willd.) Zizka] es una gramínea invasora en los pastizales de Chihuahua. Es originario del sur de África, fue introducido por primera vez en Estados Unidos en 1866 para uso ornamental (Stokes et al., 2011). Sin embargo, a partir de 1903 se consideró una especie invasora (Stevens y Fehmi, 2009). Este pasto logra invadir exitosamente diversas áreas y se ha adaptado a las zonas áridas y semiáridas del norte de México, su principal forma de dispersión es por medio de su semilla la cual es fácilmente transportada por el viento (Possley y Maschinski, 2006; Carrillo et al., 2009). Actualmente está presente en todos los estados de la República Mexicana. La Comisión Nacional para el Conocimiento y el Uso de la Biodiversidad (CONABIO, 2012), considera este pasto como una especie invasora; sin embargo, no se tiene la suficiente información de manejo y control de esta planta.
Las semillas de M. repens son capaces de germinar lentamente bajo condiciones de precipitación irregular. Esta estrategia la utiliza para evitar la germinación total del banco de semillas en suelo al inicio de la temporada de lluvia (Stokes et al., 2011). Germinan con el paso de los días sí las condiciones siguen siendo favorables, de no ser así las semillas restantes no germinarán. Esta respuesta a la ausencia o presencia de agua le proporciona competitividad sobre los pastos nativos que comúnmente germinan rápidamente al inicio de la temporada de lluvia. De la presencia y adaptación a los cambios en el ambiente del banco de semillas dependerá la presencia de su especie (Cox y Allen, 2008). En contraste el pasto nativo Bouteloua gracilis que es dominante de las regiones de pastizales del estado de Chihuahua es una planta que presenta dificultades en su establecimiento por medio de su semilla, debido a que necesita periodos continuos de humedad de entre 8 a 10 semanas para su germinación, establecimiento y crecimiento inicial, las cuales son poco frecuentes en zonas áridas y semiáridas de México (García-Sánchez y Monroy-Ata, 2005; Moreno-Gómez et al., 2012).
La ocurrencia de sequías severas es un hecho frecuente en las regiones del estado de Chihuahua, debido a que se encuentra localizado dentro de una zona semiárida (Díaz et al., 2002), la precipitación es muy variable. En el período comprendido entre 1993 a 2004 la precipitación fue inferior al promedio histórico. La sequía que se registró a nivel estatal fue considerada como grave (Núñez et al., 2007). Entre los principales factores que afectan directamente la germinación está la disponibilidad de agua para la semilla. Esto representa una limitante en las zonas áridas y semiáridas debido a que se presentan altas temperaturas durante la época de lluvia, lo que causa altas tasas de evaporación y suelo seco (Endfield y Fernández-Tejedo, 2006). Para que una planta pueda sobrevivir después de iniciada la germinación deberá de resistir largos periodos de sequía en estas zonas (Esqueda et al., 2005).
Cada especie tiene requerimientos específicos para germinar, entre estos: agua y suelo (Cox y Martin, 1984; Evans y Etherington, 1991; McLaren et al., 2004; Zeng et al., 2005). Para que un pasto logre adaptarse a un hábitat no solo necesita disponibilidad de agua, sino también influye el tipo de suelo presente en el ecosistema (Callaway et al., 2004; Warren II et al., 2012), el cual puede presentar diversas características como: los niveles de nutrientes, difusión del agua, concentración de metales y la presencia de diferentes comunidades bióticas, que en conjunto generan un efecto en el crecimiento y establecimiento de una planta (Stanton-Geddes et al., 2012). Estas características son el aspecto clave que tienen los hábitats para diferenciarse entre ellos, cada tipo de suelo presenta distintas condiciones, por lo tanto tienen diversos microorganismos que son capaces de establecer relaciones positivas o negativas con las plantas (Berg y Smalla, 2009; Zuppinger-Dingley et al., 2011). La planta, para su desarrollo y adaptación, depende del entorno, principalmente de las interacciones entre el suelo y el clima (Stanton-Geddes et al., 2012).
La respuesta que cada planta muestra al estrés ambiental determinará su establecimiento, su dinámica vegetal, producción de semillas, crecimiento e interacciones, especialmente cuando los recursos del área son limitados y extremadamente heterogéneos (Thuiller et al., 2006). La humedad del suelo afecta la composición del banco de semillas (Collin et al., 2013). Las fluctuaciones hidrológicas en los hábitats son determinantes en la tasa de germinación, debido a que los excesos o escases de humedad pueden prevenir o retrasar este proceso (Lundholm y Stark, 2007). El objetivo del presente estudio fue evaluar la emergencia y sobrevivencia de la especie invasora M. repens en suelos provenientes de cuatro áreas del estado de Chihuahua con diferentes tipos de vegetación y diversos patrones de humedad-sequía. Dada la amplia presencia del pasto M. repens se pretende que esta investigación aporte información relevante sobre la adaptación y posible distribución de esta gramínea en varios suelos provenientes de cuatro áreas con diferentes tipos de vegetación de Chihuahua.
Materiales y métodos
El experimento se llevó a cabo en el invernadero de la Facultad de Zootecnia y Ecología de la Universidad Autónoma de Chihuahua entre mayo y junio de 2013. El trabajo comprendió la evaluación de la emergencia y sobrevivencia de la semilla M. repens en suelos provenientes de cuatro áreas con diferentes tipos de vegetación presentes en el estado de Chihuahua: matorral, pastizal mediano, pastizal halófito y bosque. Las muestras de suelo, de cada tipo de vegetación, se recolectaron del estrato de 0-30 cm. Los macetas fueron de 15 cm de diámetro y 20 cm de profundidad (4.5 kg de suelo). Se utilizaron 80 macetas, 20 por cada tipo de suelo colectado. La semilla fue recolectada en el verano del 2006. Estas 20 macetas se regaron con los cuatro patrones de humedad-sequía, donde cada patrón tenía 5 repeticiones. Fueron puestas por maceta un total de 20 semillas recolectadas. La siembra se realizó el día 11 de octubre de 2012 efectuándose el primer riego en todas las macetas. Los conteos de sobrevivencia se realizaron después de 12, 24 y 36 días posteriores al último riego, cuantificando las plantas que no estaban marchitas.
Los suelos se sometieron a análisis físicos y químicos de acuerdo con el manual IRENAT (1996) en laboratorio de UNIFRUT-Cuauhtémoc, Chihuahua (Cuadro 1). Previo a los análisis, el suelo se secó al aire y se tamizó (malla de 2 mm). Las técnicas empleadas fueron: textura por el método del hidrómetro de Bouyoucos; pH, con un potenciómetro en relación suelo-agua de 1:2.5 (p/v); materia orgánica (MO), por el método de Walkley y Black. Los cationes Ca2+ y Mg2+ se extrajeron con una solución de acetato de amonio 1N a un pH = 7 y fueron cuantificados por espectrometría de absorción atómica. Los carbonatos (CO3) se extrajeron con agua destilada y su concentración se determinó por titulación ácida. Para el nitrógeno total (Nt), se usó el método Kjeldahl; para ello, una muestra de suelo se digirió con ácido sulfúrico-salicílico y el Nt se determinó por destilación en ácido bórico y titulación con ácido sulfúrico 0.05 N. La determinación de NO3 mediante la formación de un complejo coloreado con Brucina ácida (Cuadro 1).

C.E. = conductividad eléctrica.
Cuadro 1 Caracterización del suelo proveniente de cuatro áreas con diferente tipo de vegetación en el estado de Chihuahua.
En el Cuadro 1 se presenta la caracterización física y química del suelo que se recolectó en los diferentes tipos de vegetación: matorral, pastizal halófito, pastizal mediano, bosque. La descripción se encuentra en COTECOCA (1978).
El suelo de matorral es de origen aluvial profundo, color pardo oscuro a rojizo obscuro o gris rosáceo, la textura es franco-arenoso, con grava y poca piedra en el perfil y superficie. El pH de 6.5 a 7.5.
El suelo de pastizal halófito es de origen aluvial profundo, coloración pardo grisáceo muy obscuro a negro, textura arcillo limosa, su estructura son bloques angulares a columnares, con consistencia dura, drenaje interno y escurrimiento superficial lento, el pH va de 7.8 a 10.3.
El suelo del pastizal mediano es de origen aluvial, profundo, coloración pardo obscura a pardo rojizo obscura, la textura es arcillo arenosa con grava, estructura granular, la consistencia moderadamente dura, drenaje interno y el escurrimiento es superficial medio, con un de pH de 6.5 a 7.5.
El suelo de bosque es coluvial a partir de roca ígnea, la profundidad es de media a somera, con frecuente afloramientos de roca. La textura dominante es la franco-arcillosa con grava, su estructura son bloques angulares, con consistencia dura, drenaje interno medio y escurrimientos superficiales muy rápidos, el pH es de 6.2 a 6.8.
Los patrones de lluvia que se utilizaron fueron con base en 35 años de registros climatológicos. De estos datos solo 80% de ellos fueron los utilizados para la conformación de los patrones de humedad-sequía, esto debido a que en 10% de ellos se presentaron precipitaciones muy altas donde cualquier semilla podría germinar y en el otro 10% no hubo precipitación por lo tanto no habría germinación de ninguna semilla (Esqueda et al., 2005; Moreno-Gómez et al., 2012). Las categorías de los patrones de riego fueron: pobre 6 días, baja 8, media 10 y alta 12 días de lluvia correspondientes a los registros históricos de precipitación durante la época del verano cuando se presenta el período de lluvias. En él Cuadro 2 se detalla la secuencia de riego, el número inicial muestra los días que se llevó a cabo y el número de la derecha indica los días sin riego para cada patrón. Las macetas, fueron regadas con 85 mm de agua (basado en el diámetro de la maceta) para simular 10 mm de lluvia. Esta cantidad es la mínima necesaria reportada para que pueda ser aprovechada por las plantas (Melgoza et al., 2014).
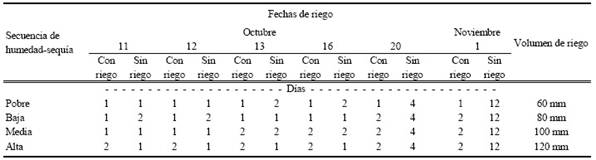
Cuadro 2 Patrones de precipitación pluvial de verano registrados en el estado de Chihuahua durante el periodo de 1965 a 1999.
Debido a que los datos obtenidos para las variables de germinación y sobrevivencia es de tipo bivariado y se expresan en proporciones, se utilizó el ajuste de un modelo para datos categóricos en los dos factores evaluados (suelos de cuatro áreas con diferente tipo de vegetación y patrones de humedad-sequía) y su interacción. De esta forma se estimó el efecto individual de cada secuencia de humedad sobre cada uno de los suelos provenientes de los tipos de vegetación. Las repeticiones consistieron de cinco macetas con veinte semillas viables cada una. La comparación de medias se realizó mediante una prueba z para proporciones en distribuciones. Posterior al análisis los datos fueron transformados a porcentajes.
Resultados y discusión
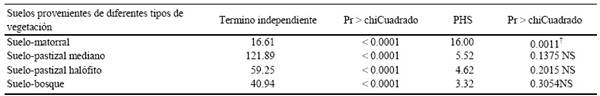
El pasto M. repens desde hace aproximadamente 30 años se encuentra presente en el estado de Chihuahua y ha invadido diversas áreas. Los resultados obtenidos en la presente investigación muestran una interacción entre los patrones de humedad-sequía y los suelos provenientes de cuatro áreas de diferente tipo de vegetación (P ≤ 0.0024) en respuesta a la tasa de germinación de M. repens (Cuadro 3).

†Efecto significativo α = 0.05; PHS = patrones de humedad-sequia; NS = no significativo.
Cuadro 3 Análisis de varianza de verosimilidad máxima para determinar efectos significativos de los tratamientos sobre la germinación de M. repens.
Emergencia de Melinis repens
M. repens es capaz de germinar en condiciones de sequía severa en cualquiera de los suelos provenientes de los tipos de vegetación evaluados. En general en este experimento, todos los tratamientos emergieron durante las dos primeras semanas posteriores al primer riego. El pasto se ha distribuido en el mundo mostrando su gran capacidad de adaptación a condiciones desfavorables de humedad y suelos pobres, convirtiéndose en invasor de zonas áridas, semiáridas y suelos erosionados (Díaz et al, 2012).
Las características físicas y químicas del suelo tienen efecto sobre la emergencia del pasto según los patrones de humedad-sequía evaluados (Esqueda et al., 2005). El pasto M. repens en su lugar de origen se encuentra en suelos con proporciones de arena que varía del 10 al 20%. El contenido de arena en suelo le permite a la planta mayor germinación, debido a las características de aeración y menor compactación (Daemane et al., 2012; Melgoza et al., 2014). Es posible que este pasto invasor se vea favorecido por la cantidad de arena presente en los suelos de la región donde el pastizal halófito tiene 13% de arena, el suelo de bosque y pastizal mediano 40% y matorral 64% de arena en su composición. Otras gramíneas de origen africano, como Eragrostis lehmanniana y E. curvula, presentes en los pastizales de Chihuahua también prefieren suelos arenosos (Cox y Martin, 1984; Esqueda et al., 2005).
La germinación es una etapa muy sensible a los cambios de humedad en el suelo. Varios autores sugieren que las semillas tienen varias respuestas a los cambios de condiciones de humedad. Una de esas estrategias menciona que la germinación disminuye cuando la humedad del suelo decrece. Esta clase de semillas tienen la capacidad de iniciar el proceso de germinación parcialmente, entrar en un estado de latencia si las condiciones son adversas y luego volver a iniciar la germinación, esto aumentará sus posibilidades de establecimiento en lugares donde la humedad en el suelo sea mínima (Evans y Etherington, 1991; Melgoza et al., 2014). La semilla de M. repens presenta esta estrategia, pues se ha observado que puede iniciar el proceso de germinación cuando se presentan las primeras lluvias, pero si las condiciones ambientales cambian y se carece de humedad suficiente en suelo, la semilla es capaz de evitar la emergencia hasta que las condiciones ambientales sean las idóneas (Carrillo et al., 2009; Díaz et al., 2012). Para que el proceso de germinación se inicie, el suelo debe de presentar condiciones de humedad próximas a -0.2 MPa. La germinación de la semilla de M. repens esta relacionada a la humedad del suelo, por lo cual se puede anticipar su posible distribución con base en los patrones de lluvia de las diferentes aéreas donde exista un banco de semillas de esta planta (Stokes et al., 2011).
La interacción entre la humedad-sequía y el suelo de vegetación proveniente del matorral tuvo efecto significativo (P ≤ 0.0011) en la emergencia de la semilla de M. repens (Cuadro 4) donde el porcentaje de germinación fue 75% en el patrón alto, 63% en el medio y 51% en bajo y pobre (Figura 1A).
†Efecto significativo α = 0.05; PHS = patrones de humedad-sequía; NS = no significativo.
Cuadro 4 Análisis de varianza de verosimilitud máxima para determinar efectos significativos de los suelos provenientes de cuatro áreas con diferentes tipos de vegetación y patrones de humedad-sequía sobre la germinación de M. repens.
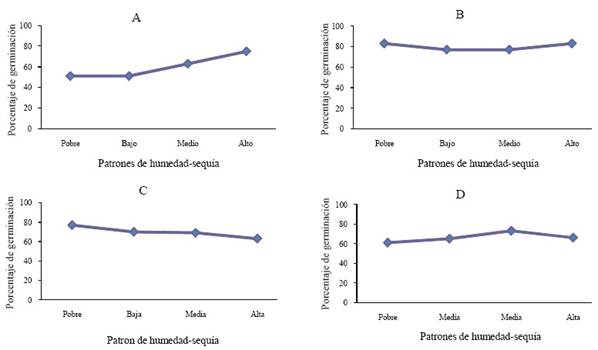
Figura 1 Porcentaje de germinación de M. repens expuesta a diferentes patrones de humedad-sequía en suelos provenientes de cuatro áreas con diferente tipo de vegetación. A) matorral; B) pastizal mediano; C) pastizal halófito; D) bosque.
El suelo proveniente del área con vegetación de pastizal mediano no mostró efecto en la interacción de germinación de la semilla y los patrones de humedad-sequía (P ≤ 0.1375) (Cuadro 4). La emergencia fue de 83% en los patrones pobre y alto, de 77% en bajo y medio respectivamente (Figura 1B). En el suelo proveniente de pastizal halófito mostró una tendencia similar (P ≤ 0.2015) (Cuadro 4). Los porcentajes de germinación para éste en los diferentes patrones fueron de 77% en pobre, 70% bajo, 69% medio y 63% en alto (Figura 1C). Este patrón de comportamiento se repite para el suelo proveniente del bosque y la interacción con los patrones de humedad-sequía (P ≤ 0.3054) (Cuadro 4), obteniendo porcentajes de germinación de 61% en pobre, 65% bajo, en medio 73% y alto de 66% (Figura 1D). Cuando el potencial de agua es bajo en el suelo, la emergencia de las semillas puede ser retrasada o acelerada dependiendo de la magnitud del déficit hídrico, afectando también la etapa de crecimiento inicial (Martínez-Ghersa et al., 1997; Cox y Allen, 2008; Fay y Schultz, 2009).
En el estado de Chihuahua 74% de la superficie es vulnerable a los impactos causados por la sequía (Núñez et al., 2007). Con los resultados encontrados se puede suponer que M. repens puede invadir diferentes áreas de vegetación tales como: matorral, pastizal mediano, pastizal halófito y bosque ubicados en la entidad, debido a que el tipo de suelo y la humedad irregular no inhibe su germinación, aunque sí provoca diferencias en la tasa de emergencia de la semilla. Sin embargo, los resultados de este trabajo muestran que el suelo proveniente del pastizal mediano puede tener mayor posibilidad de amenaza de invasión, cuando se presente cualquier tipo de precipitación, ya que, en éste se observó la mayor emergencia. Stevens y Fehmi (2009) mencionan que bajo las nuevas condiciones ambientales causadas por el cambio climático como el incremento de la temperatura y la poca precipitación, el pasto M. repens puede convertirse en un problema ecológico similar al que presenta el zacate buffel (Pennisetum ciliare (L.) Link).
Sobrevivencia de Melinis repens
La sobrevivencia de pasto M. repens después de periodos de sequía prolongados posteriores al riego, mostraron que la relación ente los suelos provenientes de cuatro áreas con diferentes tipos de vegetación y la secuencia humedad-sequía fueron significativos (P < 0.0001) en los primeros 12 días. (Cuadro 5). Tanto la germinación como el establecimiento de la semilla son etapas críticas en el ciclo de vida de las plantas (Bentsink y Koornneef, 2008). Estos dos aspectos son muy sensibles a la variabilidad ambiental y requieren condiciones favorables tanto de suelo como de agua.
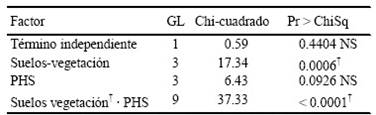
†Efecto significativo α = 0.05; NS = no significativo. PHS = patrones de humedad-sequía. Suelo-vegetación (suelos provenientes de áreas de diferentes tipos de vegetación.)
Cuadro 5 Análisis de varianza de verosimilitud máxima para determinar efectos significativos de los tratamientos sobre la sobrevivencia de M. repens a 12 días sin riego.
Es importante destacar que la tolerancia a la variabilidad de humedad del suelo también depende de la etapa del desarrollo de la planta (Fay y Schultz, 2009; Kinyua et al. 2010). La tasa de sobrevivencia en suelo proveniente de matorral y los patrones de humedad tienen interacción al doceavo día (P ≤ 0.0001) (Cuadro 6). Un 35% en la secuencia pobre, 43% en la baja, 40% en la media y 66% en el alta (Figura 2A).
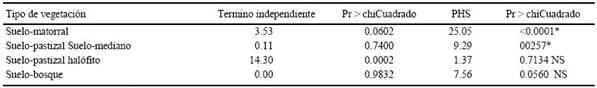
†Efecto significativo α = 0.05: PHS = patrones de humedad-sequía. NS = no significativo.
Cuadro 6 Análisis de varianza de verosimilitud máxima para determinar efectos significativos de sobrevivencia de M. repens a los 12 días en suelos provenientes de cuatro áreas con diferentes tipos de vegetación y patrones de humedad-sequía.

Figura 2 Porcentaje de sobrevivencia de M. repens expuesta a diferentes patrones de humedad-sequía en diferentes vegetaciones. A) matorral; B) pastizal mediano; C) pastizal halófito y D) bosque.
Como se muestra en el Cuadro 6 la sobrevivencia de las plantas al día 12 en el suelo colectado en pastizal mediano y los patrones de humedad-sequía tuvieron un efecto significativo en la interacción (P ≤ 0.0001). Esta variable tuvo 53% en la secuencia pobre y baja, 36% en la media y 55% en la alta (Figura 2B). El suelo proveniente de pastizal halófito no presentó un efecto significativo (P ≤ 0.7134) entre los factores evaluados en la sobrevivencia de las plantas al día 12 posterior al último riego (Cuadro 6), la sobrevivencia fue del 62% en la secuencia pobre y baja, 55% en la media y 59% en la alta (Figura 2C). En el suelo colectado en la vegetación de bosque no se observó que la interacción entre los patrones de humedad-sequía fueran significativos (P ≤ 0.0560) (Cuadro 6), esta variable se mantuvo en la secuencia pobre y baja de 54%, mientras que la alta en 38% (Figura 2D).
Se realizó un conteo de plantas sobrevivientes en el día 24 después del último riego. Sin embargo, no fue posible realizar un análisis estadístico confiable, debido a la poca sobrevivencia del pasto. Para el día 36 se observó la muerte total de las plantas de M. repens que fueron evaluadas. Stokes et al. (2011) mencionan que las plántulas de M. repens no sobreviven a períodos prolongados de sequía.
Conclusiones
De acuerdo a los patrones de humedad-sequía que se presentan en el estado de Chihuahua es factible la introducción de la gramínea invasora [Melinis repens (Willd.) Zizka] en suelos de cuatro áreas con diferente tipo de vegetación, como matorral, pastizal mediano, pastizal halófito y bosque. Su capacidad de germinar y su estrategia de detener este proceso si las condiciones no son las adecuadas para soportar periodos de precipitación irregular, la hacen una especie que puede llegar a ser una amenaza para los ecosistemas nativos del estado. Sin embargo, no solo la precipitación provoca un efecto en la germinación de esta semilla, sino también las relaciones entre la planta y el suelo pueden resultar benéficas o perjudiciales para su establecimiento. Los resultados obtenidos muestran una clara preferencia del pasto M. repens a las condiciones presentadas por el suelo proveniente de pastizal mediano, principalmente a las precipitaciones altas donde obtuvo 92% de plántulas emergidas, cabe notar que en cualquiera de los patrones de humedad-sequía esta planta obtuvo su mayor emergencia en este suelo.
En cuanto a la sobrevivencia de las plántulas emergidas, no soportan periodos largos de sequía mayores a los 24 días en ninguno de los suelos de las áreas con diferentes tipos de vegetación. Sin embargo, donde se presentó la mayor sobrevivencia fue en el suelo de bosque, probablemente debido a las condiciones que presenta este tipo de suelo. Para conocer de manera más específica los efectos que tienen los diferentes tipos de suelo en la presencia de esta planta se recomienda un estudio in situ, lo que permitirá conocer qué clase de interacciones podría promover o detener la germinación y el establecimiento de esta planta en suelos en las áreas de los diferentes tipos de vegetación presentes en el estado de Chihuahua.

