










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkUniversidad y ciencia
versión impresa ISSN 0186-2979
Universidad y ciencia vol.25 no.1 Villahermosa abr. 2009
Artículos
Análisis de la interacción genotipo–ambiente mediante el modelo Ammi y potencial de producción de triticales forrajeros (X Triticosecale Wittm.)
AMMI analysis of genotype–environment interaction and production potential of forage triticale (X Triticosecale Wittm.)
AJ Lozano–del Rio1, VM Zamora–Villa1, L Ibarra–Jiménez2, SA Rodríguez–Herrera1, E de la Cruz–Lázaro3, M de la Rosa–Ibarra4
1 Departamento de Fitomejoramiento. Universidad Autónoma Agraria "Antonio Narro". Buenavista s/n. Saltillo 25315 Coahuila, México. Tel. (844) 4 110254. jlozrio@uaaan.mx.
2 Centro de Investigación en Química Aplicada (CIQA). Coahuila.
3 Universidad Juárez Autónoma de Tabasco. Tabasco.
4 Depto. de Botánica. UAAAN. Coahuila.
Artículo recibido: 25 de octubre de 2007
Aceptado: 13 de enero de 2009
RESUMEN
Los forrajes de corte y pastoreo son muy importantes en el norte y centro de México. Debido a la respuesta diferencial de los genotipos en cada ambiente, el desarrollo de nuevas variedades implica determinar su estabilidad de producción para su posterior liberación en una región determinada. Se evaluó la producción de materia seca por corte y acumulada en dos cortes de veintidós genotipos de triticale (X Triticosecale Wittm.) facultativos e intermedios–invernales en 14 ambientes del norte de México durante los ciclos 1996–1997 al 1999–2000. La producción se analizó con el modelo AMMI (Additive Main Effects and Multiplicative Interaction) mediante la caracterización de los genotipos por la magnitud de su interacción con los ambientes (IG*A) y su estabilidad de rendimiento. El análisis por corte discriminó los materiales de forma eficiente basado en su hábito de crecimiento, clasificando con precisión a los genotipos por su producción y estabilidad. Los genotipos 3 y 5 (facultativos) fueron recomendables para un solo corte y los genotipos 7 y 13 (intermedios invernales) fueron adecuados para cortes múltiples. El genotipo 13 (intermedio–invernal) fue el más estable y productivo para materia seca acumulada. El modelo separó efectivamente los dos tipos de hábito de crecimiento en los cortes individuales, debido a diferencias en su fenología que influyeron en su producción de materia seca. El modelo también caracterizó eficazmente los genotipos por su estabilidad, detectando en forma precisa la magnitud y signo de las interacciones presentadas por cada uno, tanto en los cortes individuales como en la producción de materia seca acumulada.
Palabras clave: Facultativo, intermedio–invernal, interacción genotipo–ambiente, estabilidad, modelo AMMI.
ABSTRACT
Forage for cut and grazing are very important in northern and central Mexico. Because of the differential response of genotypes in each environment, the development of new cultivars includes determining the production stability for future release in a certain region. Dry matter yield was evaluated per cut and accumulated in two cuts of 22 facultative and winter–intermediate triticale genotypes (X Triticosecale Wittm.) in 14 environments in northern Mexico during the cycles of 1996–1997 to 1999–2000. Single and accumulated production were analysed with the AMMI model (Additive Main Effects and Multiplicative Interaction), and the genotypes were characterised based on the extent of their interaction with the environment (IG*A) and on their yield stability. The analysis per cut efficiently identified genotypes based on their growth habit, and classified them accurately with respect to production and stability. The facultative genotypes 3 and 5 were recommended for only one cut and the winter–intermediate genotypes 7 and 13 for multiple cuts. The winter–intermediate genotype 13 was the most stable and productive for accumulated dry matter. The model effectively separated the two types of growth habit in the individual cuts in response to differences in phenology that afftected dry matter production. The AMMI model also efficiently characterised the genotypes with respect to their stability, and accurately detected the extent and sign of the interactions presented by each, both in the individual cuts and in the production of accumulated dry matter.
Key words: Facultative, winter–intermediate, genotype–environment interaction, stability, AMMI model.
INTRODUCCIÓN
El triticale (X Triticosecale Wittm.) puede utilizarse para tres fines agrícolas: a) producción de grano, b) doble propósito y c) producción de forraje, ya sea para corte ó pastoreo. Esta última modalidad en el uso de este cultivo está ganando popularidad en diversas regiones del norte y centro de México durante el ciclo otoño – invierno, debido a su potencial productivo y adecuada calidad nutritiva, la cual, en la etapa de encañe–embuche, es similar a la alfalfa (Collar & Aksland 2001). El triticale presenta también mayor tolerancia a las bajas temperaturas en comparación con la avena, cultivo tradicional de invierno. Existen varios tipos de hábito de crecimiento en este cultivo: primaverales, invernales y facultativos (Anónimo 1989a) y su clasificación se basó originalmente en los hábitos de crecimiento de los trigos (Hanson et al. 1985). Recientemente, un tipo más (intermedios–invernales) fue descrito con base a su patrón de producción y capacidad de ahijamiento (Ye et al. 2001). Las características de su crecimiento permitieron describir en forma detallada los tipos actuales de triticales. Los triticales de hábito primaveral se caracterizan por su rápido crecimiento y diferenciación, sin requerimientos de vernalización, con crecimiento inicial erecto que favorece la cosecha mecánica, con amacollamiento reducido y baja capacidad de recuperación después del corte (rebrote). Estos triticales son adecuados para un solo corte. Los tipos invernales son convenientes para cortes (o pastoreos) múltiples. Los tipos facultativos son de rápido crecimiento y diferenciación, presentan crecimiento inicial semi–postrado, amacollamiento intermedio y buena capacidad de recuperación después del corte o pastoreo, por lo que son adecuados para dos cortes (verdeos) o pastoreos. Los intermedios invernales presentan crecimiento y diferenciación medios, semipostrados, con buen ahijamiento y alta capacidad de rebrote que permite dar cortes múltiples, sin ser tan tardíos como los tipos invernales (Fox et al. 1990; Royo et al. 1995; Royo & Parés 1996; Lozano–del Río et al. 1998; Giunta et al. 1999; Royo & Blanco 1999; Ye et al. 2001).
Con base a su patrón productivo y hábito de crecimiento, en México se han desarrollado materiales de triticale para uso forrajero, principalmente para cortes múltiples o pastoreo (Lozano del Río et al. 2002a; 2002b). En este tipo de explotación es imprescindible la capacidad de rebrote de los genotipos, la cual depende principalmente del hábito de crecimiento y la etapa fenológica del corte, de las condiciones climáticas, las prácticas de manejo, la humedad y fertilidad del suelo y de la presión del corte o pastoreo, entre otras (Poysa 1985). Debido a la importancia de los forrajes de corte en la época invernal en algunas regiones, como en la Laguna y el sur de Chihuahua y de pastoreo directo, como en el Norte de Coahuila y Chihuahua, el desarrollo de variedades de este cultivo que cubran las necesidades de alguno de los tipos de explotación antes mencionados, adquiere mayor relevancia. En este orden de ideas, los ensayos de rendimiento en diferentes localidades son importantes en la selección de los mejores genotipos con el fin de: (i) utilizarlos como progenitores dentro del programa de mejoramiento ó (ii) recomendar su uso por los productores en una región determinada (Crossa et al. 1990). Por otra parte, la estabilidad de producción de un genotipo a través de diferentes ambientes se debe determinar antes de su liberación y recomendación como variedad comercial para una localidad o región determinada, ya que normalmente las variedades evaluadas en ensayos multirregionales se han comportado en forma diferencial en los diversos ambientes. Esta respuesta diferencial de los genotipos a diversos ambientes es llamada interacción genotipo–ambiente (I G*A), la cual ha sido estudiada, descrita e interpretada por medio de varios modelos estadísticos (Crossa 1990). Algunos modelos, tales como análisis de varianza ó modelos de regresión sobre medias ambientales (Yates & Cochran, 1938; Finlay & Wilkinson 1963; Eberhart & Russell 1966) y los modelos de Efectos Principales Aditivos y Multiplicativos de la Interacción (AMMI) (Gauch 1988) utilizan sólo la variable fenotípica de interés, en este caso, el rendimiento de forraje.
El modelo AMMI ha mostrado ser efectivo en el análisis de ensayos multirregionales, ya que captura una gran proporción de la suma de cuadrados de la I G*A, separando en forma precisa los efectos principales de aquellos correspondientes a la interacción (Gauch 1992). El modelo integra el análisis de varianza (ANDEVA) y el Análisis de Componentes Principales (ACP) como lo mencionaron Salmerón et al. (1996). La mayoría de los trabajos con AMMI han enfatizado la producción de grano en diversos cultivos (Ebdon & Gauch 2002) y muy pocos han evaluado la producción de biomasa. Resultados que consideren cortes múltiples no se han registrado. El análisis segmentado de la información en el estudio de la IG*A se ha realizado en trabajos anteriores como el de Verma et al. (1978), quienes utilizaron los parámetros de estabilidad de Eberhart & Russell (1966) para analizar la producción de grano. Ellos separaron los ambientes con base en sus índices (sitios de alto rendimiento y sitios de bajo rendimiento) y realizaron el análisis correspondiente para cada grupo de sitios. Además, ellos propusieron una nueva clasificación de los genotipos. Asimismo, los resultados del modelo AMMI han revelado relaciones entre características morfológicas y fenológicas que causan que los genotipos interactúen con los ambientes Zobel et al. (1988) y Gauch & Zobel (1996). Aunque el análisis AMMI sólo utiliza datos de rendimiento, se ha registrado que factores ambientales tales como la precipitación, temperatura media, altitud, latitud y fertilización nitrogenada, entre otros, frecuentemente están correlacionados con los estadísticos de la interacción del modelo (Gauch 1992; Romagosa et al. 1996; Gauch 1998). En este estudio se analizaron datos de rendimiento de materia seca, en dos cortes sucesivos de forraje con el método AMMI, con el objetivo de probar la magnitud de la IG*A en la producción de forraje seco en triticales de dos diferentes hábitos de crecimiento en un ensayo multirregional en diversas localidades de los estados de Coahuila, Chihuahua y Nuevo León.
MATERIALES Y MÉTODOS
Veinte líneas avanzadas (F10) de triticale hexaploide y dos variedades comerciales del mismo cultivo (AN–31 y AN–34) de dos diferentes hábitos de crecimiento ( facultativos e intermedios–invernales) se evaluaron en un ensayo multirregional bajo riego durante los ciclos 1996–1997 al 1999–2000, en seis localidades del Norte de México ubicadas en los estados de Coahuila (Coah.), Nuevo León (N.L.) y Chihuahua (Chih.). Las líneas avanzadas (F10) fueron generadas en el Programa de Cereales de la Universidad Autónoma Agraria Antonio Narro (UAAAN) a partir de germoplasma proveniente del Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT). Las dos variedades comerciales del mismo cultivo (AN–31 y AN–34) están clasificadas como intermedios–invernales. Debido a que no todas las localidades se evaluaron en todos los ciclos, el término ambiente se utilizó para designar la combinación particular de localidad y ciclo (Tabla 1). De igual forma, genotipo fue el término genérico usado para designar los cultivares. En cada ambiente, los experimentos se establecieron en campo bajo un diseño de bloques completos al azar con tres repeticiones. Las parcelas consistieron de ocho surcos de 5 m de longitud, espaciados a 0.17 m (6.8 m2) a una densidad de siembra de 120 kg ha–1. En todas las localidades y ciclos la fecha de siembra quedó comprendida entre la segunda quincena de septiembre y la segunda de noviembre (Tabla 1). La fertilización se hizo con base a la fórmula 120–80–00, utilizando como fuentes urea y fosfato monoamónico. Todo el fósforo y el 50 % del nitrógeno se aplicó a la siembra y el resto del nitrógeno se incorporó en el primer riego de auxilio.
En todas las localidades se realizó un corte de forraje a los 90 días después de la siembra en la etapa fenológica promedio de encañe o etapa 39 de la escala de Zadoks et al. (1974). Un marco rectangular de 0.25 × 0.50 m se colocó en forma perpendicular a las hileras centrales de la parcela (0.125 m2). El forraje se cortó manualmente con rozadera a una altura de 2 cm sobre la superficie del suelo. Después del muestreo el forraje remanente se eliminó con una cortadora mecánica y se aplicaron 40 unidades adicionales de nitrógeno para promover el rebrote y obtener un segundo corte de forraje. Este se realizó en promedio de todas las localidades a los 45 días después del primero, en la etapa fenológica promedio de embuche o etapa 45 de la escala de Zadoks et al. (1974). En ambos cortes, del forraje fresco cosechado se extrajo tomó una submuestra de 250 g, la cual se secó en una estufa a 60 ° C durante 72 horas para estimar el rendimiento de materia seca. El criterio para decidir las fechas de muestreo se basó en el sistema de manejo comercial bajo el sistema de corte en cereales de invierno en las regiones de estudio de acuerdo a: 1) producción de materia seca (>3 t ha–1) y altura de planta (>35 cm) que permita una adecuada cosecha mecánica y 2) coincidencia de la producción con los meses críticos del período invernal (diciembre – febrero). Independientemente de su hábito de crecimiento, en ninguno de los dos cortes los genotipos presentaron una etapa fenológica posterior a embuche ó inicio de espigamiento.
El rendimiento de forraje seco entre ambientes por corte y acumulado fue estimado mediante análisis de varianza (ANDEVA) combinados, para detectar la presencia de IG*A. Los tratamientos se compararon con la prueba de la Diferencia Mínima Significativa (DMS) al 0.01 de probabilidad. La comparación entre los rendimientos de materia seca por corte y el acumulado, entre los dos tipos de hábito de crecimiento, se efectuó con la prueba de T2 de Hotelling (Johnson 1998), la cual es una generalización de la prueba de t utilizada para comparar dos medias de determinada variable, con la diferencia de que considera simultáneamente todas las variables en estudio (rendimiento de forraje seco en cada corte y acumulado) de la manera siguiente:
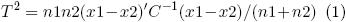
donde: n1, n2 = número de observaciones asociadas con los vectores de medias x1 y x2. C = matriz con las estimaciones conjuntas de varianza–covarianza de los n1 y n2 datos.
La significancia del valor calculado del parámetro T2 se comprobó mediante una prueba de F, cuyo valor calculado se obtuvo mediante
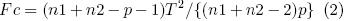
y se comparó con valores tabulados con p y (n1 + n2 – p – 1) grados de libertad, donde p representa el número de variables utilizadas en la comparación de los dos grupos de medias. Finalmente, los datos se analizaron mediante el modelo AMMI en cada uno de los dos cortes (14 ambientes) y el acumulado de ambos. El modelo del método AMMI fue el siguiente:
Donde Yij = rendimiento del i–ésimo genotipo en el j–ésimo ambiente, µ= media general, gi = desviaciones de la media de los genotipos, ej = desviaciones de las medias de los ambientes, λk = valor singular para cada Análisis de Componentes Principales (ACP), n = número de componentes principales (ACP) retenidos en el modelo, αik,  = calificación del ACP para el k–ésimo eje del i–ésimo genotipo y j–ésimo ambiente respectivamente y Rij = residual del modelo.
= calificación del ACP para el k–ésimo eje del i–ésimo genotipo y j–ésimo ambiente respectivamente y Rij = residual del modelo.
Las calificaciones del ACP para los ambientes y genotipos se expresaron como unidades del eigenvector correspondiente por la raíz cuadrada del eigenvalor (Zobel et al. 1988). La suma de cuadrados de la interacción genotipo–ambiente se subdividió en ejes del ACP, donde el eje k posee g + e – 1 – 2k grados de libertad, donde g y e representan el número de genotipos y ambientes, respectivamente. Normalmente, en el modelo son retenidos solamente los dos primeros componentes principales (CP). Los ejes (CP) no significativos son enviados al residual. En este modelo, las calificaciones asignadas a los genotipos y a los ambientes pueden tomar valores positivos o negativos con respecto al componente principal. Los genotipos considerados estables fueron aquellos que poseen valores cercanos a cero (menor interacción). Los valores mayores indican una mayor interacción con los ambientes y dependiendo del signo se realizó una descripción más amplia de los genotipos. Las diferencias estadísticas registradas en la interacción genotipo–ambiente en los ANDEVA's, se analizaron con el modelo AMMI, en el cual, etiquetando los genotipos se etiquetaron por su hábito de crecimiento (F = facultativo y + = intermedio–invernal) para explicar con mayor precisión las tendencias observadas y determinar el grado de estabilidad de la producción de los genotipos a través del ciclo. El análisis bajo este modelo se realizó mediante SAS, versión 6 (Anónimo 1989b).
RESULTADOS
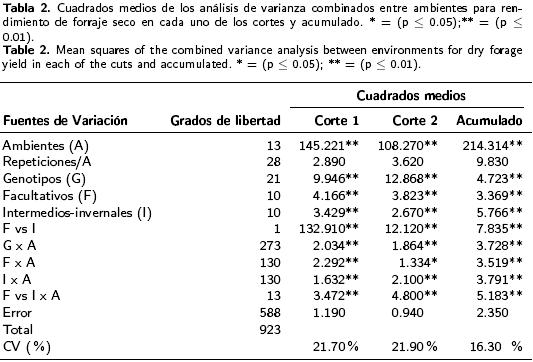
El rendimiento de materia seca en los cortes 1 y 2 y el acumulado de ambos registró diferencias estadísticas (p ≤ 0.01) para los efectos principales de ambientes, genotipos y para la interacción genotipo * ambiente, con coeficientes de variación considerados aceptables para este tipo de variables (Tabla 2). El efecto principal de genotipos y el contraste entre los dos hábitos de crecimiento registraron diferencias altamente significativas tanto dentro de cada tipo como entre ellos (p ≤ 0.01). La diferencia en producción entre facultativos e intermedios–invernales resultó significativa (Tabla 2), aunque esta diferencia disminuyó conforme se consideró el segundo corte o el acumulado. En lo referente a la partición de la IG*A se confirmó que todos los tipos de triticales presentaron interacción con el ambiente (p < 0.01), aunque de manera diferencial de acuerdo a este contraste (Tabla 2).
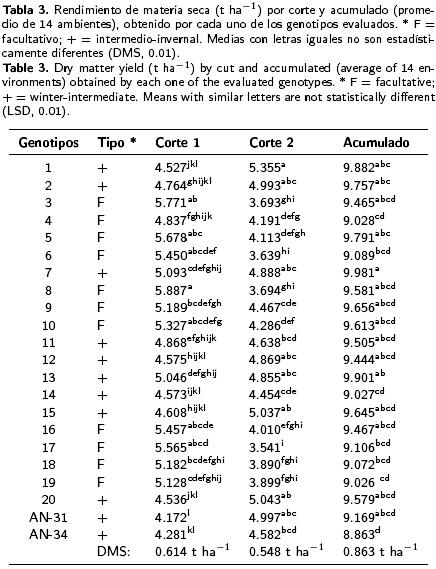
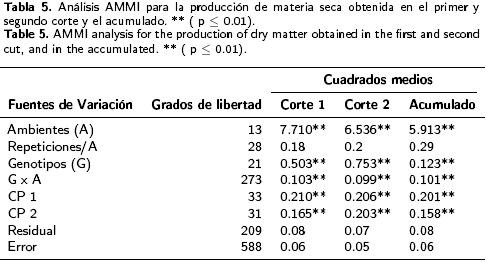
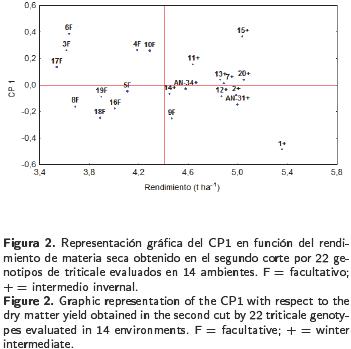
Las comparaciones entre las medias de los genotipos durante el primer corte revelaron que los rendimientos promedios a través de ambientes variaron entre 4.172 y 5.887 t ha–1. En el segundo corte, los rendimientos promedios oscilaron variaron 3.541 y 5.355 t ha–1 , mientras que la productividad promedio de forraje seco acumulado varió entre 8.863 y 9.981 t ha–1 (Tabla 3). Los genotipos 7, 13 y 1 de hábito intermedio–invernal registraron el mayor rendimiento biológico acumulado en los dos cortes, con 9.981, 9.901 y 9.882 t ha–1 de forraje seco, respectivamente. En el primer corte, el rendimiento de forraje seco en los tipos facultativos (F) fue significativamente mayor (> 16.3%) que el de los tipos intermedio–invernal (+) (T2 de Hotelling; p < 0.01). Lo inverso sucedió en el segundo corte, en donde los tipos intermedio–invernales produjeron mayor cantidad de forraje seco que los facultativos (> 23.6 %). No obstante, entre los dos tipos en la producción acumulada de forraje seco no se registraron diferencias estadísticas, pero sí entre genotipos del mismo hábito de crecimiento (Tabla 4). En el análisis efectuado para el primer corte de forraje, el modelo AMMI reportó diferencias altamente significativas entre ambientes, genotipos y la IG*A. Los CP 1 y 2 explicaron un 43 % de la interacción (Tabla 5) y permitieron clasificar como estables a los genotipos 3, 5, 10 y 19 (F) y 14 y AN–34, (+), los cuales presentaron valores del CP cercanos a cero (Figura 1). Independientemente de la estabilidad mostrada por estos genotipos, al graficar el rendimiento promedio de los mismos contra el primer eje del análisis de componentes principales (CP 1), mediante el trazo de una línea en el eje de las X para representar la media general de producción (Figura 1), los tipos facultativos, y específicamente los genotipos 3, 5 y 10 (F) mostraron significativamente mayor rendimiento que los genotipos 14 y AN–34 (+), confirmando el patrón productivo entre hábitos de crecimiento registrado en la prueba de T2 para el primer corte (Tabla 4). Excepto los genotipos 7 y 13, los tipos + mostraron rendimientos menores que la media, debido a su desarrollo más lento al inicio del ciclo de crecimiento. Aunque estable, el genotipo 14 (+) presentó un rendimiento significativamente menor (Tabla 3), el cual fue similar al de los genotipos 20 y 12 (+), aunque este último con una mayor interacción. Por su rendimiento destacaron los genotipos 3, 5 y 8 (F), aunque en este último la interacción resultó negativa (Figura 1). El grupo formado por los genotipos 9, 10, 18 y 19 (F) y 7 y 13 (+) presentaron rendimientos similares, pero difirieron en la magnitud de sus interacciones, ya que 7 y 9 fueron positivas y 13 y 19 negativas, pero menores que las del genotipo 18, lo cual evidenció la variabilidad presente entre los genotipos (Figura 1). Al analizar la producción del segundo corte, el AM–MI reportó diferencias estadísticas (p ≤ 0.01) entre ambientes, genotipos y la interacción, así como los CP 1 y 2, que juntos explicaron un 48 % de la interacción (Tabla 5). Los tratamientos 2, 7, 12, 13, 14, 20, AN–34 (+) y 5 (F) resultaron estables dada la menor magnitud de sus interacciones. Los genotipos identificados con los números 1, 15, 11 (+) y 9 (F), aunque no estables, presentaron rendimientos iguales o superiores a la media y pudieran considerarse para regiones particulares.

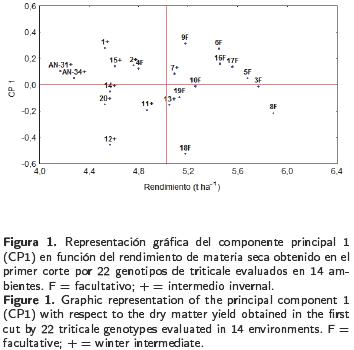
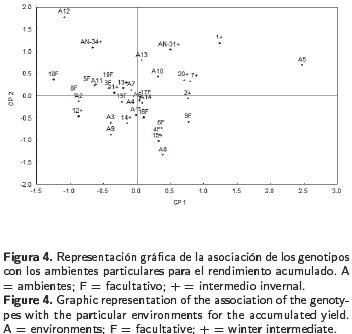
En contraste al primer corte, los genotipos intermedio–invernales (+) mostraron mayor rendimiento y estabilidad que los facultativos (F), que aparecen a la izquierda de la línea trazada en el eje X que representa el rendimiento promedio (Figura 2), lo cual fue demostrado por la prueba de T2 de Hotelling (Tabla 4). El grupo formado por los tratamientos 1 y AN–31 (+) presentó rendimientos estadísticamente iguales al tratamiento 15 (+), pero con interacciones de diferente signo. Al analizar la información del rendimiento acumulado en los dos cortes, el método AMMI reportó diferencias altamente significativas entre ambientes, genotipos y la interacción, así como para los CP 1 y 2, con los cuales se explicó un 42% de la interacción (Tabla 5) y se calificó a los genotipos 16, 17, 19 (F) y 13 y 14 (+) como estables (Figura 3). Los genotipos 1, 7 y 13 (+) destacaron por su rendimiento. Los dos primeros con grandes interacciones positivas. La producción del genotipo 18 (F) fue inferior a la media, con mayores interacciones negativas. Los testigos comerciales AN–31 y AN–34 (+) resultaron contrastantes, ya que AN–31 rindió en forma similar a los genotipos 4, 6 y 17 (F), con interacciones positivas, mientras que AN–34 registró el menor rendimiento acumulado (Tabla 3) e interacciones negativas. Al graficar el rendimiento de los genotipos contra el primer componente, no se observó una separación de los mismos de acuerdo a su hábito de crecimiento, ya que ambos tipos no fueron significativamente diferentes entre sí, lo que fue confirmado por la prueba de T2 Hotelling (Tabla 4), aunque con una pequeña ventaja de producción para los tipos intermedio–invernales (9.523 vs 9.354 t ha–1 ). El modelo AMMI también permitió realizar una doble representación de genotipos y ambientes en un mismo gráfico, lo que a su vez permitió asociar genotipos con ambientes particulares. Las asociaciones del rendimiento acumulado de los genotipos con respecto a su interacción con los ambientes revelaron que el genotipo más estable fue el 17, de tipo facultativo, el cual mostró una estrecha asociación con los ambientes 4, 6, 7 y 14, que fueron los que menos contribuyeron a la IG*A; por el contrario, ambientes como el 5, 12 y 8 aportaron más a la IG*A (Figura 4). Algunos genotipos registraron asociaciones específicas con ambientes como la mostrada por la variedad AN–34 en el ambiente 12, o los genotipos 4, 6 y 15 en el octavo ambiente. La variedad comercial AN–31 y los genotipos 2, 7 y 20 se asociaron más a los ambientes 10 y 13.
DISCUSIÓN
La relación significativa detectada entre ambientes confirmó las diferencias entre las localidades, principalmente a lo que se refiere a clima y tipo de suelo. Las diferencias estadísticas registradas entre los genotipos confirmaron la distinta fenología de los materiales de acuerdo a su hábito de crecimiento, mientras que el efecto significativo estimado en la interacción genotipo–ambiente demostró la repuesta diferencial de los genotipos al evaluarse en diferentes condiciones ambientales. Los genotipos evaluados parecen diferenciarse más en sus patrones de producción a través de cortes sucesivos (Lozano–del Río et al. 1998), y no en su potencial de producción total de forraje a través del ciclo de cultivo. De esta manera, genotipos de diferente hábito de crecimiento quedaron incluidos en los mismos grupos de significancia. Las diferencias entre los hábitos de crecimiento fueron detectadas en forma más precisa por la utilización de la prueba de T2 de Hotelling. En este estudio, el comportamiento productivo de los genotipos en cada uno de los cortes estuvo principalmente determinado por su hábito de crecimiento, que influyó en su patrón de producción. Esto conllevó a que los genotipos facultativos produjeran en el primer corte mayor cantidad de materia seca que los tipos intermedio – invernales. Esta superioridad detectada en el primer corte por los tipos facultativos, posiblemente se debió a que presentaron una etapa fenológica promedio más avanzada que los tipos invernales (facultativos: etapa promedio=39–43; intermedios–invernales: etapa promedio=31–33) (Zadoks et al. 1974). Los tipos facultativos también mostraron una buena capacidad de rebrote, pero de menor magnitud que los intermedios invernales, que se han caracterizado por un mayor ahijamiento (Lozano et al. 1998; Giunta et al. 1999; Royo & Blanco 1999; Ye et al. 2001), característica que les permitió producir mayor cantidad de materia seca en el segundo corte. Después de un corte o pastoreo, los triticales invernales producen más biomasa y área foliar que los tipos facultativos, debido a su mayor capacidad de amacollamiento (Royo & Romagosa 1996).
Holiday (1956a) y Scott & Hines (1991) concordaron también en que al avanzar la edad de las plantas en las gramíneas se elevaron los puntos de crecimiento, lo cual favoreció su remoción por los cortes o pastoreos y disminuyó el número de vástagos o hijuelos que sobreviven. En base al aspecto fisiológico del rebrote, el alargamiento de los culmos ó tallos en los cereales al avanzar la edad de las plantas, como señalaron Holiday (1956a; 1956b) y Scott & Hines (1991), provocó que por efecto del corte se eliminaran algunos puntos de crecimiento, lo que disminuyó el número de vástagos o hijuelos que sobrevivieron y esto afectó la producción de biomasa. Los tipos facultativos, en etapa fenológica más avanzada, fueron más afectados en su rebrote que los tipos invernales, que registraron una mayor capacidad de amacollamiento y una etapa fenológica menos avanzada. El manejo de triticales para los diferentes tipos de explotación (un solo corte y cortes o pastoreos múltiples) requiere del conocimiento del hábito de crecimiento de la variedad y de la etapa fenológica en la época del corte, además de la longitud del período de crecimiento después del corte y su capacidad de rebrote, ya que existe diversidad genética para estas características (Sharrow 1990; García del Moral 1992).
Los resultados de este trabajo sugieren que en forma general, el hábito de crecimiento y la capacidad de rebrote de los tipos en estudio, favoreció a los intermedios invernales, los cuales se caracterizan por poseer una mayor capacidad de recuperación después del primer corte o pastoreo, que depende de la etapa en que sean cortados o pastoreados, ya que a menor etapa, mayor capacidad de rebrote (Royo et al. 1995; Royo & Romagosa 1996; Lozano–del Río et al. 1998; Royo & Blanco 1999; Ye et al. 2001). Este hábito de crecimiento mostró en general interacciones positivas, o negativas pero pequeñas, salvo los genotipos 12 y AN–34, lo que indicó, en términos generales, que los tipos intermedios– invernales son los más adecuados para cortes múltiples. En cambio, los facultativos resultaron adecuados para un solo corte, debido a que presentaron una mayor acumulación de biomasa en el período inicial en comparación con los tipos intermedio–invernales. En este sentido, la información generada por el modelo AMMI en este estudio en los cortes individuales concordó con lo mencionado por Ebdon & Gauch (2002), quienes señalaron que algunas características morfológicas o fenológicas de las plantas tienen algún efecto causal sobre la interacción de los genotipos con los ambientes.
El análisis segmentado (por corte) con el modelo AMMI clasificó en forma precisa a los genotipos por la magnitud de su interacción con el ambiente. La separación de los dos tipos de hábitos de crecimiento con respecto a su patrón de producción en los análisis individuales por corte se debió a diferencias en la etapa fenológica de cada tipo. El modelo fue también eficiente en la caracterización de los genotipos por su estabilidad al analizarse la producción acumulada, lo cual permitió discriminar en forma gráfica la magnitud y el signo de la interacción de cada uno de ellos, independientemente de su hábito de crecimiento y constituye una buena herramienta en el estudio de la interacción genotipo–ambiente en la evaluación de genotipos en ensayos en localidades múltiples.
Con respecto a su patrón de producción, los genotipos facultativos se recomiendan para un solo corte. Los genotipos 3, 5, 10 y 19 fueron los más apropiados para las diferentes zonas donde se llevó a cabo el estudio, dado sus altos niveles de productividad y estabilidad. Los tipos intermedio–invernales son más recomendables para cortes o pastoreos múltiples, aunque pudieran existir genotipos facultativos que pueden adaptarse a esta condición en determinados ambientes, tal como lo demostró el elevado promedio de producción de materia seca obtenido por los genotipos 5 y 9 (F). Estos resultados apoyan también la posibilidad de diseñar mezclas forrajeras que combinen los diferentes patrones productivos para obtener una producción más uniforme a través de los cortes, favorecida por el posible efecto de relevo de los distintos tipos estudiados.
LITERATURA CITADA
Anónimo (1989a) NRC National Research Council. Triticale: A promising addition to the world's cereal grains. National Academy Press, Washington, D.C. 105 pp. [ Links ]
Anónimo (1989b) SAS/STAT User's guide. Version 6. Fourth edition. SAS Institute Inc. Cary. 943 pp. [ Links ]
Collar C, Aksland G (2001) Harvest effects on yield and quality of winter forage. Proc. 31st California Alfalfa and Forage Symposium. , Ca. U.C. Cooperative Extensión. University of California, Davis. Modesto: 133–142. [ Links ]
Crossa J (1990) Statistical analysis of multilocation trials. Advances in Agronomy 44: 55–85. [ Links ]
Crossa JL, Gauch HG, Zobel RW (1990) Additive main effects and multiplicative interaction analysis of two international maize cultivar trials. Crop Science 30: 493–500. [ Links ]
Ebdon JS, Gauch HG (2002) Additive main effect and multiplicative interaction analysis of national turfgrass performance trial: I. Interpretation of genotype × environment interaction. Crop Science 42: 489–496. [ Links ]
Eberhart SA, Russell WA (1966) Stability parameters for comparing varieties. Crop Science 6: 36–40. [ Links ]
Finlay KW, Wilkinson GN (1963) The analysis of adaptation in a plant breeding program. Australian Journal of Agricultural Research 14: 742–754. [ Links ]
Fox PN, Skovmand B, Thompson BK, Braun HJ, Cormier R (1990) Yield and adaptation of hexaploid spring triticale. Euphytica 47: 57–64. [ Links ]
García del Moral LF (1992) Leaf area, grain yield and yield components following forage removal in triticale. Journal of Agronomy and Crop Science 168: 100–107. [ Links ]
Gauch HG (1988) Model selection and validation for yield trials with interaction. Biometrics 44: 705–715. [ Links ]
Gauch HG (1992) Statistical analysis of regional yield trials: AMMI analysis of factorial designs. Elsevier. New York. 278 pp. [ Links ]
Gauch HG (1998) MATMODEL Version 2.1: AMMI and related analyses for two–way data matrices. Microcomputer Power. Ithaca. 59 pp. [ Links ]
Gauch HG, Zobel RW (1996) AMMI analysis of yield trials. In: M S Kang and H G Gauch, Jr. (eds) Genotype by environment interaction. CRC Press. New York. 416 pp. [ Links ]
Giunta F, Motzo R, Deida M (1999) Grain yield analysis of a triticale (X Triticosecale Wittmack) collection grown in a Mediterranean environment. Field Crops Research 63: 199–210. [ Links ]
Hanson H, Borlaug NE, Anderson RG (1985) Trigo en el tercer mundo. El Batán, México. CIMMYT. D.F. 166 pp. [ Links ]
Holiday R (1956a) Fodder production from winter– sown cereals and its effect upon grain yield. Field Crops Abstracts 9: 129–135 [ Links ]
Holiday R (1956b) Fodder production from winter– sown cereals and its effect upon grain yield (concluded). Field Crops Abstracts 9: 207–213 [ Links ]
Johnson DE (1998) Applied multivariate methods for data analysis. Duxbury Press. Belmont. 567 pp. [ Links ]
Lozano–del Río AJ, Zamora VM, Solís HD, Mergoum M, Pfeiffer WH (1998) Triticale forage production and nutritional value in the northern region of México. In: Proc. 4th International Triticale Symposium.Jul. 26–31, Red Deer, Alberta, Canadá. pp: 140–142. [ Links ]
Lozano–del Río AJ, Colín–Rico, Pfeiffer WH, Mergoum M, Hede A, Reyes–Valdés MH (2002a) Registration of "TCLF–AN–31" Triticale. Crop Science 42: 2214–2215. [ Links ]
Lozano–del Río AJ, Colín–Rico, M Pfeiffer WH, Mergoum M, Hede A, Reyes–Valdés MH (2002b) Registration of "TCLF–AN–34" Triticale. Crop Science 42: 2215–2216. [ Links ]
Poysa VW (1985) Effect of forage harvest on grain yield and agronomic performance of winter triticale, wheat and rye. Canadian Journal of Plant Science 65: 879–888. [ Links ]
Romagosa I, Ullrich SE, Han F, Hayes PM (1996) Use of the additive main effects and multiplicative interaction in QTL mapping for adaptation in barley. Theoretical Applied Genetics 93: 30–37. [ Links ]
Royo C, Parés D (1996) Yield and quality of winter and spring triticales for forage and grain. Grass and Forage Science 51: 449–455. [ Links ]
Royo C, Romagosa I (1996) Effect of forage removal at the first detectable node stage on the growth of winter and spring triticale. Grass and Forage Science 51: 170–179. [ Links ]
Royo C, Blanco R (1999) Growth analysis of five spring and five winter triticale genotypes. Agronomy Journal 91: 305–311. [ Links ]
Royo C, Soler C, Romagosa I (1995) Agronomical and morphological differentiation among winter and spring triticales. Plant Breeding 114: 413–416. [ Links ]
Salmerón ZJJ, Cabañas BC, Chávez J, Valenzuela MV (1996) Agrupación de ambientes de temporal y genotipos de avena con el modelo AMMI. Revista Fitotecnia Mexicana. 19: 151–162. [ Links ]
Scott WR, Hines SE (1991) Effect of grazing on grain yield of winter barley and triticale: the position of the apical dome relative to the soil surface. New Zealand Journal Agricultural Research. 34: 177–184. [ Links ]
Sharrow SH (1990) Defoliation effects on biomass yield components of winter wheat. Canadian Journal of Plant Science 70: 1191–1194. [ Links ]
Verma MA, Chahal GS, Murty BR (1978) Limitations of conventional regression analysis: a proposed modification. Theoretical Applied Genetics. 53: 89–91. [ Links ]
Yates F, Cochran WG (1938) The analysis of groups of experiments. Journal of Agricultural Science (Cambridge) 28: 556–580. [ Links ]
Ye CWE, Díaz SH, Lozano AJ, Zamora VM, Ayala MJ (2001) Agrupamiento de germoplasma de triticale forrajero por rendimiento, ahijamiento y gustosidad. Técnica Pecuaria en México 39 (1): 15–30. [ Links ]
Zadoks JC, Chang TT, Konzak CF (1974) A decimal code for the growth stages of cereals. Eucarpia Bulletin 7: 42–52. [ Links ]
Zobel RW, Wright MJ, Gauch HG (1988) Statistical analysis of a yield trial. Agronomy Journal 80: 388–393. [ Links ]

