











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The ocean surface layer (0-100 m) off the Baja California Peninsula is influenced by the southern end of the California Current (CC). This region is considered a transition zone (Roden 1971) due to the confluence of water masses with different physicochemical properties: Subarctic Water that is transported southward by the CC, Tropical Surface Water, and Subtropical Surface Water that flows from the areas south and southwest of the peninsula (Hickey 1998, Durazo 2015). Biological diversity and the physicochemical variability in the region are modulated by the interactions and boundaries between water masses and their spatiotemporal variability (Bograd and Lynn 2003, Gaxiola-Castro et al. 2008). For example, at the seasonal scale, the CC is more intense and influences the entire region off Baja California during spring, whereas during autumn-winter the weakening of this current allows waters of tropical and subtropical origin to enter the region. This variability divides the peninsula into 2 regions: one with subarctic characteristics to the north of Punta Eugenia and one with warmer conditions to the south of Punta Eugenia (Durazo 2009, 2015). This seasonal change in environmental conditions is reflected in the variations of phytoplankton biomass and production and of zooplankton groups (Lavaniegos and Ohman 2003, Gómez-Ocampo et al. 2017a).
Phytoplankton biomass and production off Baja California have different spatiotemporal scales. High chlorophyll a (Chla, mg·m-3) values were reported for spring as a result of intensified wind stress, which causes upwelling of nutrient-rich water along the peninsula (Durazo 2015). The mesoscale eddies, fronts, and meanders that occur as a result of friction between currents have caused changes in biomass and phytoplankton groups (Espinosa-Carreon et al. 2004, 2012). At the interannual scale, events such as the subarctic water intrusion in 2002-2006, the Blob, and the 2015-2016 El Niño caused phytoplankton biomass and production to decline (Espinosa-Carreón et al. 2015, Gómez-Ocampo et al. 2017b). Therefore, understanding changes in phytoplankton biomass and production in the region off Baja California at these scales and their possible implications on the food web is essential to determine oceanic carbon flux and changes in the upper trophic levels, such as zooplankton, small pelagic fish, and/or fish larvae.
In the present analysis all data collected between 1997 and 2016 by the IMECOCAL (Spanish acronym for Mexican Investigations of the California Current) program are used. The analysis will allow a more precise evaluation of average conditions and seasonal variability in phytoplankton biomass and production in the region. For this reason, the aim of this article is to characterize the spatiotemporal variability in the water column using Chla and primary production data obtained during the IMECOCAL oceanographic campaigns. The results are used to discuss the main seasonal and interanual processes that impact the pelagic ecosystem in this region.
Materials and methods
The data used for this study were collected during approximately 60 cruises that were carried out along the station grid that covers the study area (Fig. 1). Water samples for the Chla (mg·m-3) analysis were collected with 5-L Niskin bottles (General Oceanics) at standard depths (0, 10, 20, 50, 100, 150, and 200 m). Each sample was analyzed with the fluorescence method (Yentsch and Menzel 1963, Holm-Hansen et al. 1965). Primary production (PP, mg C·m-3·h-1) was determined in situ with the 14C method (Steeman-Nielsen 1952) in samples collected between 11:00 and 13:00 local time at 6 irradiance levels (100%, 50%, 30%, 20%, 10%, and 1% surface irradiance) and incubated for ~2 h in the water column. The results of both biological variables were vertically integrated using the trapezoidal rule to estimate integrated Chla (Chla int, mg·m-2) from the surface to 100 m depth and integrated PP (PPint, mg C·m-2·h-1) from the surface to the euphotic zone depth (1% surface irradiance).
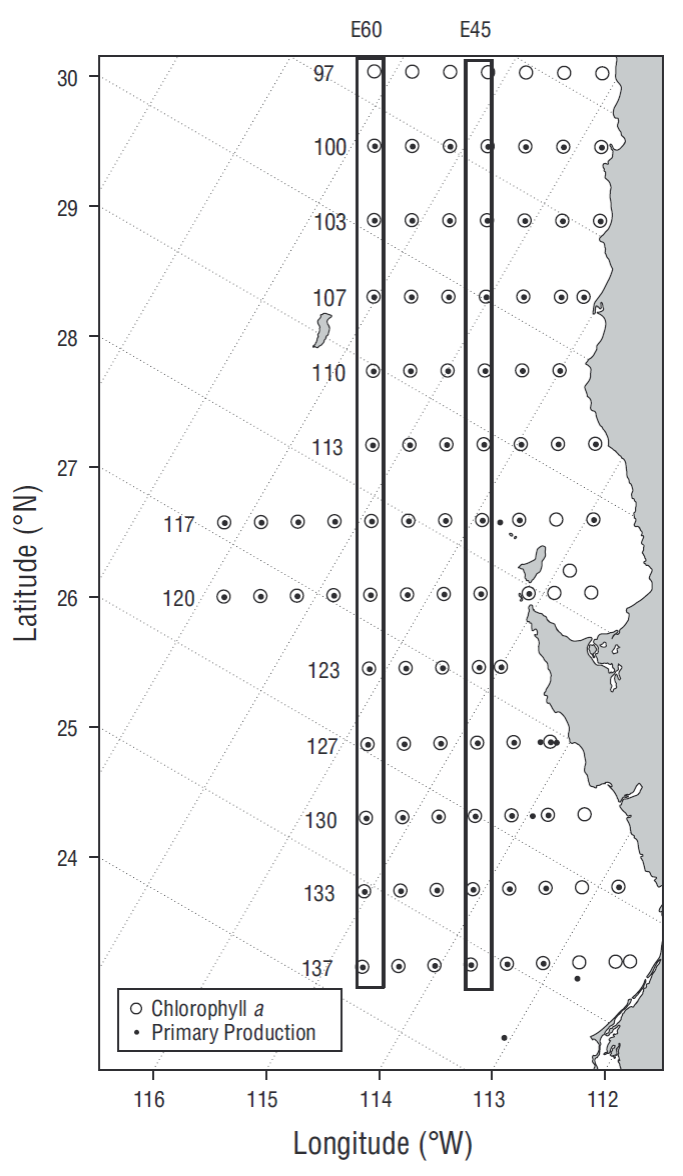
Figure 1 Hydrographic lines and stations of the IMECOCAL program where chlorophyll a (○) and primary production (•) were measured. Boxes indicate the transects of stations 45 (E45) and stations 60 (E60), representative of transition and oceanic conditions, respectively.
In order to represent seasonal variation of productivity in the water column, seasonal composites (because for some seasons there was only one datum) of Chla int and PPint at the IMECOCAL sampling stations were calculated for winter (December, January, and February), spring (March, April, and May), summer (June, July, and August), and autumn (September, October, and November). Interannual variations were analyzed using the time series of the Chla int and PPint anomalies, which were defined as the observed value minus the average value calculated from 1997 to 2016. These variations were associated with 3 indexes representative of the interannual variability in the Pacific Ocean: the Pacific Decadal Oscillation (PDO; Mantua et al. [1997], http://jisao.washington.edu/pdo/PDO.latest), the Multivariate El Niño/ Southern Oscillation (ENSO) Index (MEI, https://www.esrl.noaa.gov/psd/enso/mei/rank.html), and the North Pacific Gyre Oscillation index (NPGO, Di Lorenzo et al. [2008]).
Results
Average conditions
Productivity in the water column was represented by integrated Chla and PP values (Fig. 2 a, b). The highest Chla int values (~150 mg·m-2) were observed along the coastal zone of the study area, between Vizcaíno Bay and Punta Baja (28ºN-30ºN, Fig. 2a). The oceanic zone shows the lowest biomass values (<50 mg·m-2) in the water column. However, a site with slightly higher values (~70 mg·m-2) than the ones observed for the oceanic zone is detected off Punta Eugenia (28ºN). The highest PPint values (150 mg C·m-2·h-1, Fig. 2b) in the region off Baja California occurred to the south and to the north of Punta Eugenia (28ºN), and to a lesser degree in the coastal zone (~100 mg C·m-2·h-1). Average PPint values were lower (<50 mg C·m-2·h-1) in the oceanic zone, except at one high productivity site off Punta Eugenia (~125 mg C·m-2·h-1).
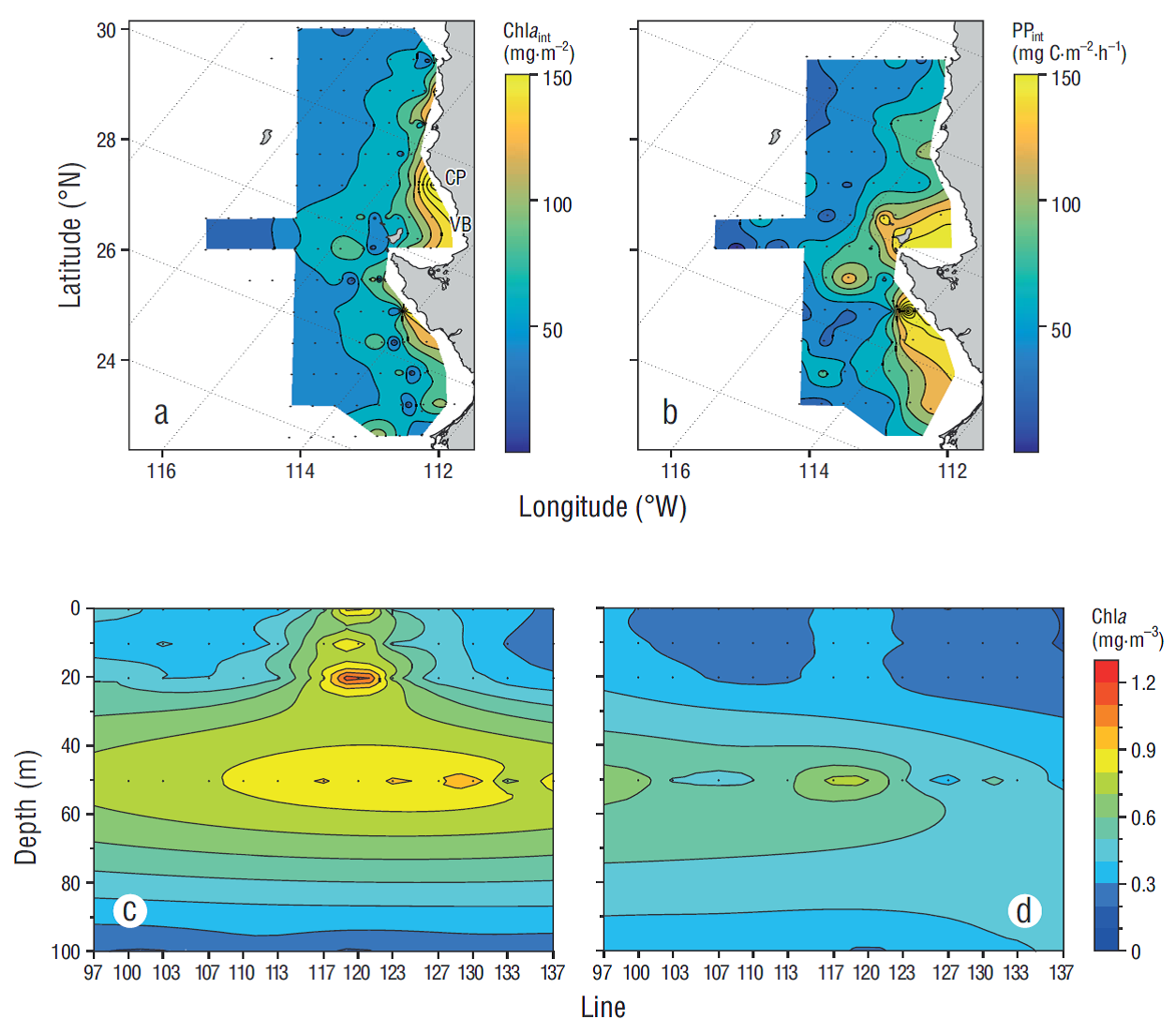
Figure 2 Mean horizontal distribution of in situ integrated chlorophyll a (Chla int, a) and primary production (PPint, b), and vertical mean conditions (c, d) for chlorophyll a (Chla) from the surface to 100 m depth. The vertical distributions correspond to station 45 (E45, c) and station 60 (E60, d) of each IMECOCAL line. Geographic references are Vizcaíno Bay (VB) and Canoas Point (CP). Mean conditions were estimated with at least 2 samples per station.
Average Chla values across stations 45 of each transect (E45) are highest at 20 m depth on line 117 (~1.3 mg·m-3), which is located off Vizcaíno Bay (28ºN). The Chla maximum (~0.8 mg·m-3) was generally located at 50 m depth. Likewise, on stations 60 of each transect (E60) (Fig. 2d) the highest Chla value (~0.9 mg·m-3) is observed at 50 m depth on line 117. In addition, the formation of a boundary or a division between the northern (97-123) and southern (127-137) lines is observed. This division occurrs around lines 117 and 120, and separates the water column into 2 zones, a mesotrophic (0.2 mg·m-3 < Chla < 1 mg·m-3) zone in the northern portion and an oligotrophic (Chla < 0.2 mg·m-3) zone in the southern portion, according to the classification proposed by Kahru and Mitchell (2002).
Seasonal variations of phytoplankton biomass and production
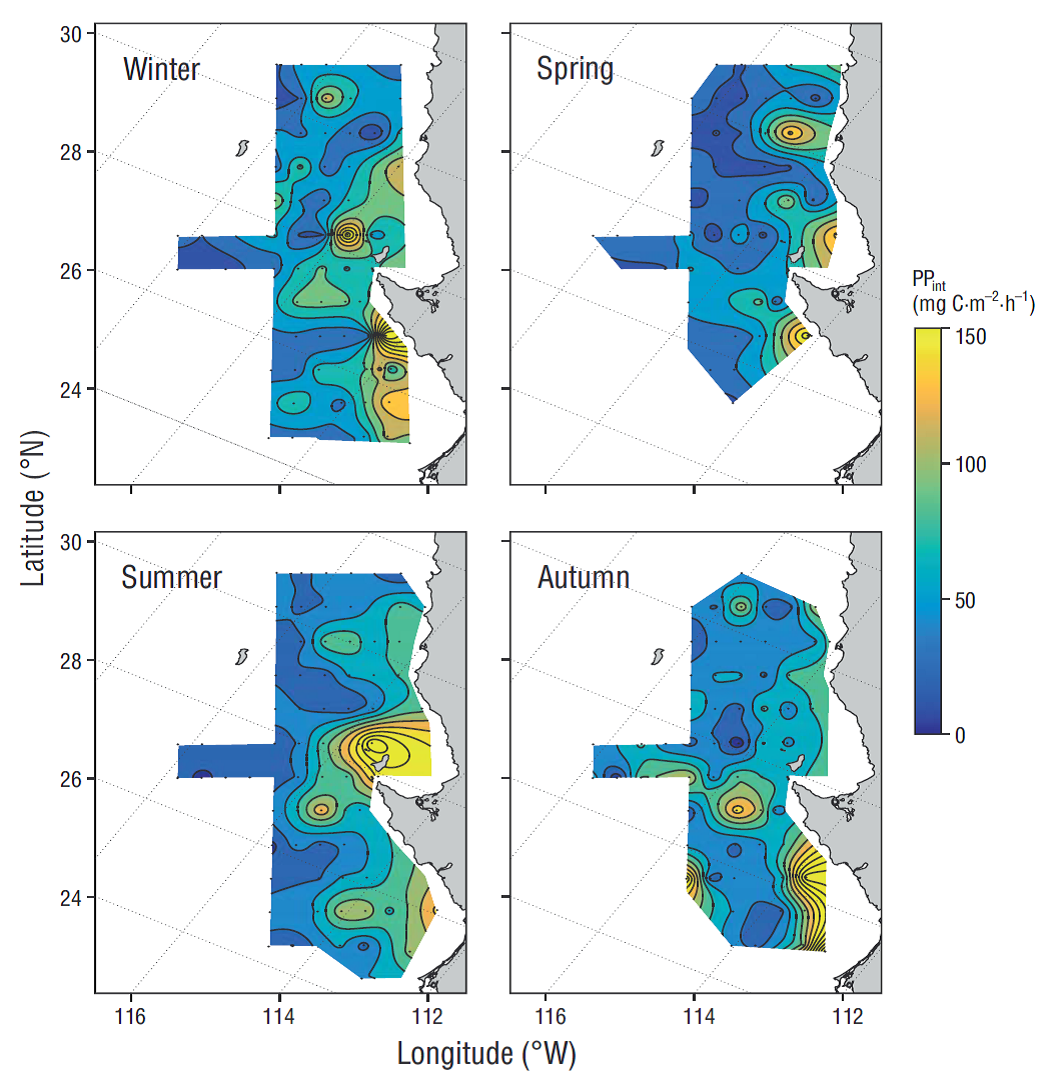
The estimated phytoplankton biomass and production seasonal averages estimated for the IMECOCAL area (Figs. 3, 4) showed that the highest values of phytoplankton biomass integrated in the water column occurred during winter and spring along the coastal zone and in some parts of the oceanic zone (Fig. 3). In winter a site with outstandingly high Chla int values (~300 mg·m-2) was observed off Punta Eugenia. In spring the highest Chla int values (~300 mg·m-2) occurred to the north of the study area. During summer and autumn, integrated biomass values were similar in the coastal and oceanic zones (<100 mg·m-2). The seasonal variation in the study area was higher for PPint (Fig. 4) than for Chla int. In winter the highest PPint values (~150 mg C·m-2·h-1) were observed to the south of Punta Eugenia (28ºN). In spring PPint maxima (~150 mg C·m-2·h-1) occurred in the region to the north of Punta Eugenia. During summer, the highest PPint values (~150 mg C·m-2·h-1) occurred in Vizcaíno bay, whereas in the region to the south and off Punta Eugenia the highest values occurred during autumn.
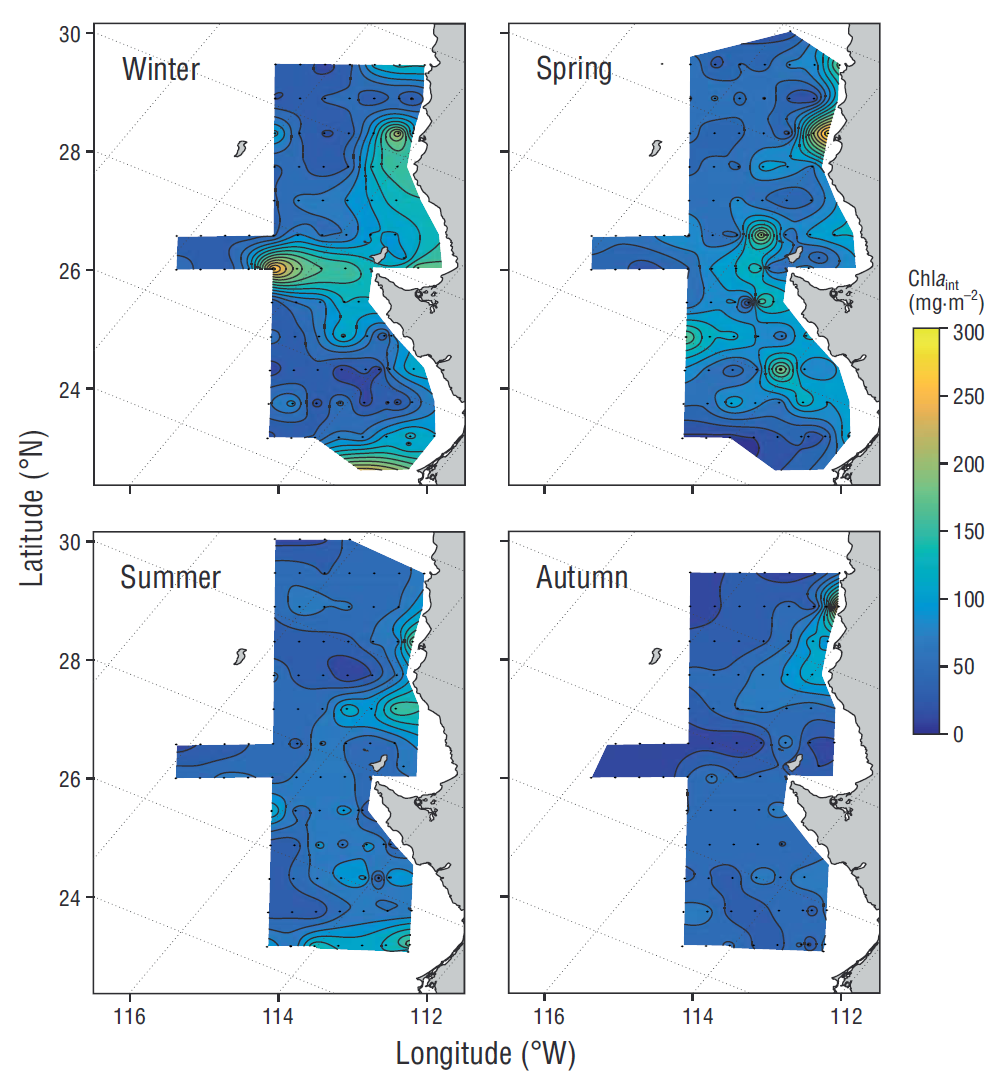
Figure 3 Seasonal variability of integrated chlorophyll a (Chla int) in winter, spring, summer, and autumn.
Phytoplankton biomass and production anomalies (1997-2016)
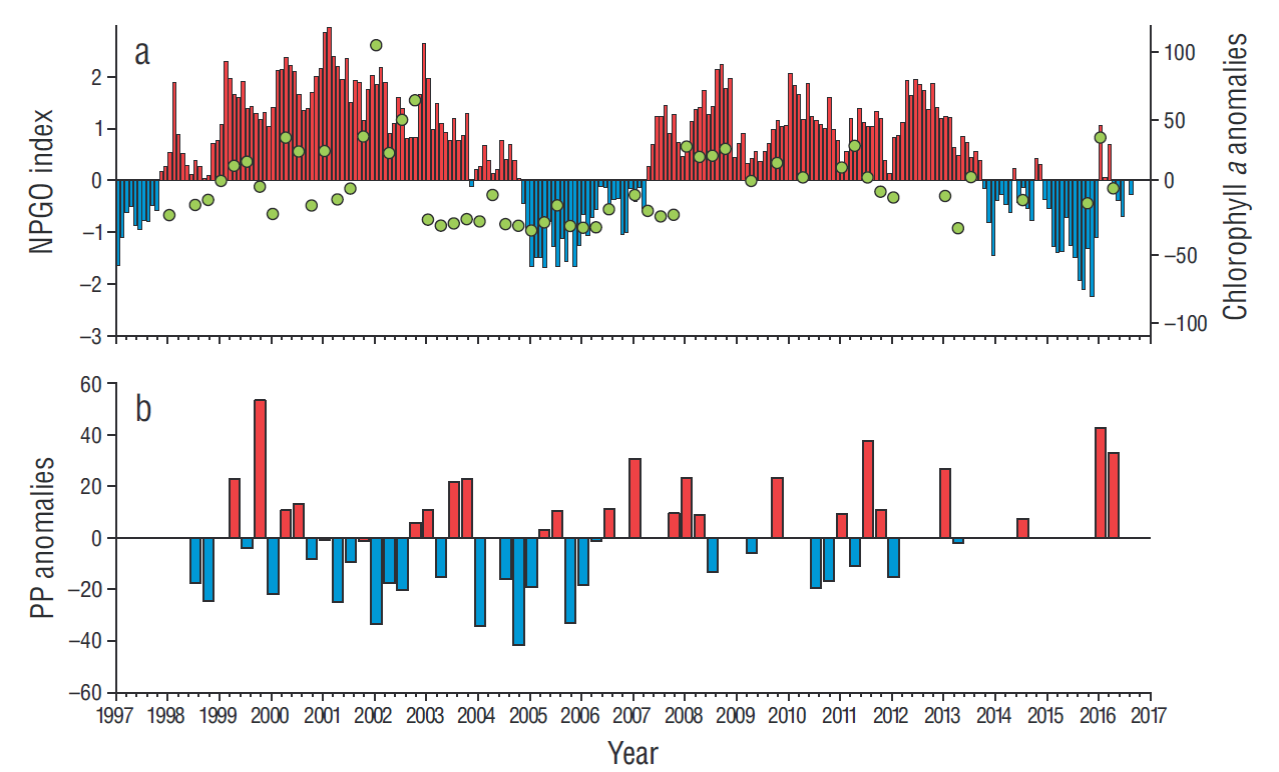
Notable interannual variations of phytoplankton biomass and production integrated in the water column were observed throughout the 20 years of IMECOCAL surveys. Phytoplankton biomass decreased during various periods: El Niño in 1998, the subarctic water intrusion from 2002 to 2007, the Blob, and the 2015-2016 El Niño. The largest declines in Chla concentrations were observed during the subarctic water intrusion (approximately -30 mg·m-2) and El Niño events (approximately -20 mg·m-2). In contrast, the 1999 and 2008-2010 La Niña events and the intensified upwelling during 2002 increased phytoplankton biomass by up to 90 mg·m-2 (Fig. 5a). Unlike biomass, phytoplankton production (Fig. 5b) did not exhibit clear responses to interannual events. However, noticeable periods of PPint increase were observed during the 1999 and 2008 La Niña events, in late 2014 during the transition period between the Blob and the 2015-2016 El Niño event, and in late 2016 during the ENSO neutral period. The largest PPint decrease occurred immediately after the 1998 El Niño event (-30 mg C·m-2·h-1) and during most of the period of subarctic water intrusion (up to -40 mg C·m-2·h-1 below the average).
Discussion
Gaxiola-Castro et al. (2010) used measurements from 11 years of surveys by the IMECOCAL program to characterize average conditions and the seasonal variability of Chla, Chla int, and PPint in the region off Baja California. In the present study, in addition to refining these averages using in situ measurements from 20 years, our aim was to analyze horizontal productivity using phytoplankton production and biomass integrated in the water column, and the vertical distribution of Chla in 2 representative transects off Baja California. We also analyze the effect of the different large-scale events that occurred in 2010-2016 on productivity. Since phytoplankton is the main component in the marine trophic food web, this type of study becomes relevant to understand the regional variations of upper trophic levels.
High productivity was observed in the Vizcaíno Bay coastal zone and in the oceanic zone off Punta Eugenia (28ºN). Vizcaíno Bay is characterized as a mesotrophic environment influenced by CC waters, coastal upwelling events off Punta Canoas (29ºN), and an anticyclonic surface eddy that advects the cool, nutrientrich surface water into the bay (Martínez-Fuentes et al. 2016). The subarctic conditions conveyed by the CC, and the upwelling of nutrient-rich waters sustain the high productivity in this region. In addition, outside the bay, there are 2 cyclonic structures separated by Punta Eugenia (Durazo 2015), which raise isopycnals in the oceanic zone and thus increase nutrient availability for phytoplankton. Therefore, the area covering Vizcaíno Bay and waters near Punta Eugenia is a physically dynamic environment that favors the regional high productivity observed in PPint and Chla int distributions (Fig. 2).
Water column productivity in the oceanic zone was divided into 2 regions, a mesotrophic one covering the northernmost lines and an oligotrophic one covering the southernmost lines, with a marked maximum at station 45 of line 117. These 2 productivity regions are a result of the influence of subarctic water transported by the CC throughout the year in the area north of Punta Eugenia (Durazo 2015), and of the input of nutrient-rich waters from the southern mouth of Vizcaíno Bay and coastal upwelling (Martínez-Fuentes et al. 2016). In contrast, the area to the south of Punta Eugenia is characterized by the seasonal influence of warm waters of tropical and subtropical origin (Durazo 2015). Furthermore, the semipermanent cyclonic gyre off Punta Eugenia could have favored the high productivity site found in this area, which can be observed by the surface chlorophyll maximum at station 45 of line 117 and in the Chla int and PPint average distribution. The high productivity observed off Punta Eugenia could be the result of upwelling waters produced by the effect of the headland (Largier et al. 2006). On the other hand, the regional division into productivity areas is consistent with the 2 regions proposed by Durazo (2015), one with subarctic characteristics to the north of Punta Eugenia and another one with warm water influence to the south.
In the coastal zone, phytoplankton biomass maxima, as seen by Chla int in the water cloumn, occurred in spring and minima occurred in autumn. The observed maxima took place during the period of most intense coastal upwelling in the IMECOCAL region (Durazo 2015). Wind stress along the coast prompts upwelling of nutrient-rich subsurface waters, which favors phytoplankton growth and biomass increase. During autumn upwelling weakens and the influence of warm waters of subtropical and tropical origin to the north of Punta Eugenia causes biomass to decline because the nutricline deepens and the input of nutrients to the euphotic zone decreases. These results concur with the results obtained by Gaxiola-Castro et al. (2010) with 10 years of phytoplankton biomass and production data analysis for the IMECOCAL area. Nevertheless, the seasonal analysis of Chla int and PPint spatial distributions allowed us to identify high productivity zones and their association with ocean circulation processes.
The seasonal variability of phytoplankton production in the water column showed a different pattern from that observed for biomass. The highest PPint values occurred during spring and summer to the north of Punta Eugenia, whereas to the south of this location the highest values occurred during winter and autumn. Furthermore, during summer and autumn, the cyclonic gyre off Punta Eugenia affected this zone and increased productivity. During spring and summer, the turbulent, nutrient-rich environment originated by coastal upwelling, mainly to the north of Punta Eugenia (Durazo 2015, Martínez-Fuentes et al. 2016), favors the presence of larger phytoplankton cells (20-200 μm) such as diatoms (Hinz et al. 2012). On the contrary, during autumn and winter the warm and oligotrophic environment prompted by the intrusion of tropical and subtropical waters in the area to the south of Punta Eugenia provides ideal conditions for the presence of smaller phytoplankton cells (e.g., Picoplankton 0.2-2.0 μm and nanoplankton 2.0-20 μm) (Barber and Hiscock 2006). The differences in phytoplankton cell sizes have been associated with the magnitudes of 2 photosynthetic parameters: maximum utilization coefficient at low light conditions (αB) and maximum photosynthetic rate (P m B ) (Coté and Platt 1983). A P m B increase could be associated with the photoacclimation of small phytoplankton cells (e.g., cyanophytes and prochlorophytes) to high irradiances, as has occurred in highly stratified environments where the thermocline and the mixed layer are shallow. Geider et al. (1993) observed P * m changes between different phytoplankton cell sizes and reported that large diatoms showed low P * m at the same growth irradiance. Moreover, Gómez-Ocampo et al. (2017a) observed higher P B m in the IMECOCAL area during high stratification periods. This could explain PPint maxima to the south of the IMECOCAL area during autumn and winter (Fig. 4d, a). On the whole, oceanic circulation in the IMECOCAL area, which is characterized by the water masses in it, determines phytoplankton habitat and favors the presence of various phytoplankton cell sizes with different photosynthetic efficiencies.
The largest declines in phytoplankton biomass and production integrated in the water column occurred during the subarctic water intrusions and El Niño events. In the 20-year time frame, the period of subarctic water intrusion was always associated with the largest productivity decline off Baja California. This could be due to the high water-column stratification and pycnocline deepening that occurred during this period, particularly in the oceanic zone (Gómez-Ocampo et al. 2017a). Both conditions limited nutrient input in the euphotic zone, and this was reflected in the reduction of phytoplankton biomass and primary production rates. However, the effect of zooplankton grazing on low chlorophyll values should not be discarded because zooplankton volumes increased from 2003 to 2013 (Lavaniegos et al. 2015). The other period with decreased productivity occurred during the Blob and El Niño 2015-2016, and it was consistent with that reported by Gómez-Ocampo et al. (2017b), who analyzed satellite-derived chlorophyll data and modeled PP data for the southern end of the CC. Unlike in the period of subarctic water intrusion, during this period stratification was high but the pycnocline was shallower. This could explain the less drastic, yet high, productivity decline during this period. Moreover, zooplankton volume decreased drastically during the Blob; hence, grazing pressure was discarded as an influential factor in the decline of chlorophyll concentrations (McClatchie et al. 2016).
In order to associate atmospheric and oceanic processes with productivity anomalies, correlations between the inter-annual indexes (NPGO, PDO, and MEI) and the time series of Chla int and PPint anomalies were calculated for the analyzed period. Correlations with MEI and PDO, though significant, were low (<0.3, results not shown). On the other hand, the highest correlation (~0.5, 95% confidence interval) occurred between NPGO and the time series of Chla int anomalies. The high correlation in the area was previously reported for Chla int anomalies (Gaxiola-Castro et al. 2008) and zooplankton (Lavaniegos et al. 2015). Negative NPGO values are associated with the weakening of the CC and strengthening of the Aleutian Low (weakening winds along the coast), which altogether reduce the input of cool, subarctic waters, weak coastal upwelling (Chhak and Di Lorenzo 2007, Di Lorenzo et al. 2008), and consequently nutrient input into the euphotic zone. Lavaniegos et al. (2015) observed significant correlations between sea surface salinity anomalies and NPGO in 4 regions off Baja California, with the presence of more saline waters during the positive NPGO phases, probably due to the strengthening of coastal upwelling. During the 2 periods of highly reduced productivity, the phase of the NPGO was negative (Fig. 5). This correlation indicated that atmospheric processes originating in the North Pacific modulate productivity off the Baja California Peninsula.

