











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The genus Sargassum C Agardh (Phaeophyceae, Fucales, Sargassaceae) has a worldwide distribution, especially in tropical and subtropical regions (Noiraksar et al. 2014), including the Gulf of California (Pacheco-Ruíz et al. 1998). In this gulf, Sargassum species constitute the bulk of the fleshy macroalgal biomass, providing habitat for diverse communities of epiphytic algae and a surface canopy for understory algae, invertebrates, and fish (Norris 2010).
Sargassum species from the Gulf of California could serve as a source of alginate (Rodríguez-Montesinos et al. 2008) and could be used in agricultural fertilizers, in forage for goats and sheep, in balanced diets for chickens and shrimp, and in human nutrition (Marín-Álvarez et al. 2003; Casas-Valdez et al. 2006a, 2006b; Casas-Valdez 2009). Extracts of Sargassum have antimicrobial activity against pathogenic organisms, and hence, possess potential in the pharmaceutical industry (Muñoz-Ochoa et al. 2010). Studies of Sargassum sinicola determined some potential uses of alginate as raw material for cell immobilization in wastewater treatment and promoting plant growth (Yabur et al. 2007), and capacity to absorb heavy metal wastes from mining industries (Huerta-Díaz et al. 2007, Patrón-Prado et al. 2010). To date, Sargassum beds have not been harvested in the Gulf of California for commercial purposes (Casas-Valdez 2009). For commercial planning purposes, it is necessary to estimate biomass (Andrefouet et al. 2004).
Research on Sargassum in the Gulf of California indicates a seasonal pattern of growth and abundance. In general, the maximum growth rate and length of the thallus occur in the spring, as reported for S. sinicola (Núñez-López and Casas Valdez 1997) and S. lapazeanum (Rivera and Scrosati 2006). In spring, the highest growth rates coincide with the maximum biomass reported for species along the eastern coast of the Baja California Peninsula: S. sinicola (Espinoza-Avalos 1990), S. horridum (Muñeton and Hernández-Carmona 1993), and S. lapazeanum (Rivera and Scrosati 2006) in La Paz Bay; S. sinicola, S. herporhizum, and S. lapazeanum (Núñez-López and Casas-Valdez 1996) in Concepción Bay; and S. herporhizum, S. johnstonii, and S. sinicola from Mulegé to San Luis Gonzaga (Pacheco-Ruíz et al. 1998). Sargassum has maximum growth, length of thallus, and biomass when the water temperature is relatively cold, which occurs in spring, and the slowest growth during late summer (Espinoza-Avalos 1993).
The total biomass of Sargassum beds (in wet tons) has been estimated at several sites along the eastern coast of the peninsula: La Paz Bay (18,900 ± 928), Concepción Bay (7,250 ± 530), and from Mulegé to San Luis Gonzaga (154,559 ± 16,139) (Hernández-Carmona et al. 1990, Casas-Valdez et al. 1993, Pacheco-Ruíz et al. 1998). However, to the best of our knowledge, these are the last estimates to have been published and current estimates are necessary because the distribution and coverage may have changed in the past 2 decades.
To update the information, the location, surface canopy area, biomass, and standing crop of Sargassum beds along the southeastern coast of the Baja California Peninsula (Mexico) were surveyed and compared with previous studies. We also determined whether biomass was related to latitude. To assess the potential for sustainable exploitation, a new database of information on coverage and seasonal abundance for Sargassum, within this area, is vital.
Materials and methods
Study area
Surveys were made from La Ventana (southern La Paz Bay; 24o02´N, 109o59´W) to La Salina (27o52´N, 112o46´W) (Fig. 1). Three localities were studied: (I) Mulegé (26o56´N, 111o58´W to 27o59´N, 112o43´W), (II) Concepción Bay (26o52´N, 111o54′W to 26o 35´N, 111o 43W), and (III) La Paz Bay (24o45´N, 110o28´W to 24o08´N, 110o24´W). These sites were selected because data of Sargassum beds were available for them. The specific location of each bed within each zone was the same or at least very similar to the sites of previous studies. This made comparisons more useful. The surveys were made during spring because the previous studies of Sargassum beds were performed during spring.
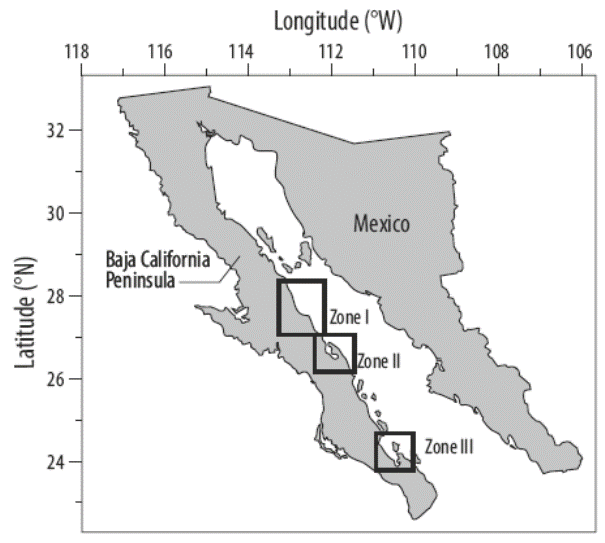
Figure 1 Study area, southeastern coast of the Baja California Peninsula (Mexico): Mulegé (zone I), Concepción Bay (zone II), and La Paz Bay (zone III).
Along the coast of the Baja California Peninsula, there are conspicuous areas where Sargassum grows, each separated from its neighbor. These beds contain a community of species, where each community can grow up to 25 m. The upper fronds reach the surface, forming a dense surface canopy. Our survey included 23 beds in zone I, 22 beds in zone II, and 27 beds in zone III.
Proportion of species and length
Since several species of Sargassum are found along this coast, the proportion of each species was determined by using a random sampling design in each bed. Three quadrats were distributed randomly. All individuals within a quadrate were collected and later identified to species, using 2 identification guides (Setchell and Gardner 1924, Dawson 1944). The Sargassum specimens were measured for length; 50 thalli of each species were measured from holdfast to apex to obtain average length in the 3 zones. Length is directly related to biomass, as described by Rivera and Scrosati (2006).
Surface canopy, biomass, and standing crop
Sargassum beds were classified as small (1 to 500 m), medium (501 to 1,500 m), or large (>1,501 m) according to their length (Hernández-Carmona et al. 1990, Casas-Valdez et al. 1993). Sargassum beds run parallel to the shoreline. Bed length is the linear distance of each bed parallel to the shore, and width is the linear distance from near-shore to faroffshore, perpendicular to the shoreline. To avoid exceeding 10% error, the width of the beds was measured every 50 m for small beds, every 100 m for medium beds, and every 200 m for large beds. Bed perimeters and surface canopy areas were calculated using the ArcView 3.2 software (ESRI 2008).
To determine average algal biomass, quadrats were used as sampling units. All Sargassum thalli in each quadrat were collected by hand and weighed (wet weight [WW]) to the nearest 0.1 kg. Ten quadrats were used for small beds, distributed randomly in the total bed area using a random number chart; 15 quadrats were used for medium beds and 20 quadrats for large beds (Hernández-Carmona et al. 1990, Casas Valdez et al. 1993). The standing crop of each Sargassum bed was determined by multiplying its average biomass by its total surface canopy area. The standing crop of each zone was calculated by adding the standing crop of beds in each zone.
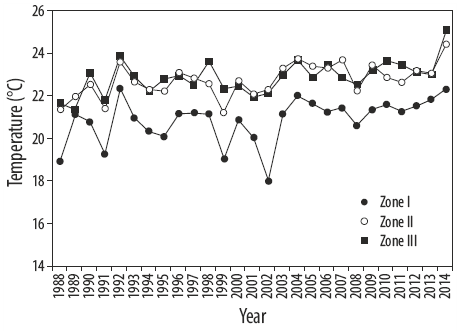
For all beds, substratum material (sand, rock, or pebbles), and depth were described for every station. From these observations, an algal bed typology was created. Sea surface temperature was obtained from satellite data for 1988 through 2014 (http://oceanwatch.pifsc.noaa.gov/). The relationships between Sargassum biomass and latitude, and seawater temperature were determined by linear correlation analysis (Cohen et al. 2003).
Spatial and temporal changes
ANOVA tests determined significant differences in the average biomass, surface canopy, standing crop, and length of the Sargassum species among the 3 zones. Differences among means were analyzed with Tukey's test, with confidence set at P < 0.05, using SAS 6.12 software. Surface canopy area, biomass, and standing crop of Sargassum beds obtained in spring of 2006 were compared with the estimated data for spring of 1988 (Hernández-Carmona et al. 1990), 1991 (Casas-Valdez et al. 1993), and 1995 (Pacheco-Ruíz et al. 1998). The area surveyed by Pacheco-Ruíz et al. (1998) extended from 26o55´N, 111o58´W to 27o57´N, 111o45´W. The area surveyed by Casas-Valdez et al. (1993) extended from 26o55´N, 111o54´W to 26o35´N, 111o45´W. The area surveyed by Hernández-Carmona et al. (1990) extended from 24o22´N, 110o12´W to 24o08´N, 110o24´W. Surveys were conducted in spring because Sargassum grows more quickly and reaches its maximum length and biomass during this season, as previously mentioned. An ANOVA test determined if there were significant differences in biomass between the years.
Results
Spatial changes
Two species in the family Sargassaceae were identified: Sargassum sinicola subsp. camouii (EY Dawson) JN Norris & Yensen and S. lapazeanum Setchell & NL Gardner. Both were found in La Paz Bay (zone III) in a proportion of 70% and 30%, respectively, of thalli. In zones I and II, only S. sinicola subsp. camouii was found.
The shortest (P = 0.00002) average thallus length of S. sinicola subsp. camouii was recorded at La Paz Bay (0.9-2.1 m) and the longest (P = 0.00002) at Mulegé (1.5-19.0 m). In zone II, thallus length ranged from 0.1 to 3.0 m (n = 2,917). The length of S. lapazeanum in zone III (0.9-1.3 m) was shorter (n = 1,947; P = 0.00002) than the length of S. sinicola subsp. camouii.
Sargassum beds were restricted to shallow waters (0.30-4.0 m) in zone III, intermediate depths (0.5-5.0 m) in zone II, and a broad range (1.0-25 m) in zone I. The holdfasts grow on stone, rocks, and pebbles along exposed and protected shores. Sargassum beds were absent on sandy bottoms.
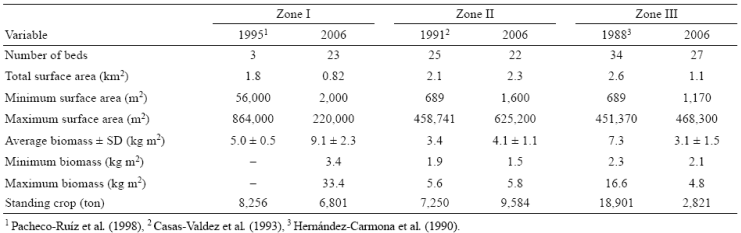
In 2006, 72 Sargassum beds were located along the southeastern coast of the peninsula. They covered a surface area of 4.22 km2, with an estimated standing crop of 19,206 t. Zone I contained 23 beds, zone II contained 22 beds, and zone III contained 27 beds (Fig. 1, Table 1). The highest average biomass was found at Mulegé (zone I), with significant difference (P = 0.000001) with respect to the other 2 zones (Table 1). The greatest surface canopy and standing crop occurred at Concepción Bay (zone II).
Table 1 Temporal changes in the number of beds, surface area, biomass, and standing crop of Sargassum spp. at Mulegé (zone I), Concepción Bay (zone II), and La Paz Bay (zone III).
In general, the average biomass of Sargassum beds increased with latitude (R2 = 0.59, P = 0.0592) (Fig. 2), that is, biomass is inversely related to water temperature. Average temperature in spring decreased with increasing latitude (Fig. 3). The average biomass of Sargassum beds decreased with increasing temperature (R2 = 0.52, P = 0.0000). The length of S. sinicola subsp. camouii was correlated with the depth of the beds (R2 = 0.71, P = 0.0000).
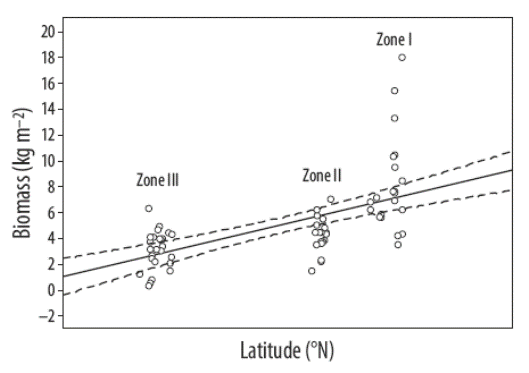
Figure 2 Relationship between Sargassum biomass and latitude along the southeastern coast of the Baja California Peninsula (dashed lines are residuals; solid line is adjusted line).
Temporal changes
Zone I
In 1995, Pacheco-Ruíz et al. (1998) found 3 large beds and the smallest covered an area of 56,000 m2. In 2006, we found 23 smaller beds and surface canopy ranged from 1,450 to 220,000 m2. The surface canopy of Sargassum beds in 1995 was 45% greater than the surface canopy in 2006 (1.8 and 0.82 km2, respectively). While the average biomass reported in 1995 (5 kg m-2 WW) was 55% lower than in 2006 (9.1 kg m-2 WW), the standing crop in 1995 (8,256 t WW) was 18% greater than in 2006 (6,801 t WW) (Table 1).
Zone II
While Casas-Valdez et al. (1993) found 25 beds in 1991, we found 22 beds in 2006. The beds at 3 sites (Buenaventura Norte, Cajede, and Tasajera) were absent in 2006. The surface canopy of beds showed high variations between 1991 and 2006: the area of some remained about the same, such as bed 29 (27,374 to 27,200 m2) and bed 24 (49,070 to 46,700 m2); the area of some increased enormously, such as bed 45 (65,446 to 625,200 m2), bed 37 (689 to 137,750 m2), and bed 25 (9,609 to 382,800 m2); and the area of some decreased significantly, such as bed 43 (458,741 to 14,950 m2), bed 39 (145,847 to 7,200 m2), and bed 31 (108,062 to 1,600 m2).
In 1991, biomass varied from 1.9 kg m-2 WW at bed 31 to 5.6 kg m-2 WW at bed 45. In 2006, biomass varied from 1.5kgm-2 WW at bed 24 to 5.8kg m-2 WW at bed 27. Average biomass was significantly higher (P = 0.00244) in 2006 (4.1 kg m-2 WW) than in 1991 (3.4 kg m-2 WW). Biomass in 2006 was higher than in 1991 at all beds, except beds 45, 44, and 24. Standing crop in 2006 was 9,584 t WW, 24% higher than in 1991 (7,250 t WW) (Table 1).
Zone III
While 34 beds were found in 1988 by Hernández Carmona et al. (1990), in 2006 we found 27 beds. The disappearance of large beds at Ensenada Verde-Puertecitos (451,370 m2), Sur de Las Ánimas (60,653 m2), and Ensenada San Gabriel at Isla Espíritu Santo (162,021 m2) is important. The total surface of Sargassum beds in 1988 was 2.6 km2, more than twice the area measured in 2006 (1.1 km2) (Table 1). The surface area of all beds was greater in 1988 than in 2006, except that of beds 68 (1988 = 24,564 m2; 2006 = 43,500 m2) and 57 (1988 = 7,436 m2; 2006 = 50,900 m2).
Average biomass in 1988 (7.3 kg m-2 WW) was significantly greater (58%) than in 2006 (3.1 kg m-2 WW) (P = 0.000001). Biomass for all beds was lower in 2006 than in 1988 (Table 1). The greatest differences occurred at bed 57 (4.1 kg m-2 WW in 2006 and 16.6 kg m-2 WW in 1988, a decline of 61%) and bed 47 (3.2 kg m-2 WW in 2006 and 11.2 kg m-2 WW in 1988, a decline of 71%). The standing crop of Sargassum in 2006 (2,821 t WW) was 85% lower than in 1988 (18,900 t WW).
Discussion
Size comparisons
Sargassum sinicola subsp. camouii was found at all sampling stations, regardless of depth, and was the dominant species in biomass. This is related to its reproductive strategies and persistence over long time periods (Espinoza-Avalos 1990). Sargassum lapazeanum was only abundant in shallow waters with rocky bottoms in zone III.
The highest average length of S. sinicola subsp. camouii was recorded in zone I, which results from higher nutrient content in northern waters (Halfar et al. 2004, 2006) and deeper sea floor. Here, S. sinicola subsp. camouii occurred at depths from 1 to 25 m. Sargassum gradually increased in average length with distance from the shore (Yeong et al. 2012). Pacheco-Ruíz et al. (1998) reported a mean length of 1.4 ± 0.4 m for this area. The length of S. sinicola subsp. camouii that we recorded ranged from 1.3 to 19 m (mean: 5.2 ± 3.1 m). Our study is the first to record a long length in this zone. A similar length was reported for S. johnstonii along the northeastern coast of the peninsula (Pacheco-Ruíz et al. 1998). The average length of S. sinicola subsp. camouii in the 3 zones was longer than the maximum length reported for the same species in Magdalena Bay (40 cm; SánchezRodríguez and Cervantes-Duarte 1999).
In Concepción Bay, the average length of S. sinicola subsp. camouii was 2.1 m and maximum length was 7 m in the spring of 1991 (Casas-Valdez et al. 1993, Núñez-López and Casas-Valdez 1997). In contrast, in 2006, the average length was 0.86 m. In 2006, the beds were located closer to the coast at depths of 0.5 to 5 m; in 1991, the beds were at depths of 0.5 to 15 m (Casas-Valdez et al. 1993). The depth at which Sargassum grows depends on the presence of hard substrata (Staehr et al. 2008, Tweedley et al. 2008).
The length of S. lapazeanum was smaller than that of S. sinicola in Concepción Bay in 1991 (Núñez-López and Casas-Valdez 1997) and at bed 74 in La Paz Bay in 2000 (Rivera and Scrosati 2006). Differences in length are largely determined by genotype characteristics (Núñez-López and Casas-Valdez 1997).
Average Sargassum biomass was lower in zones II and III in 2006 than along the northeastern coast of the peninsula (8.7-22 kg m-2 WW in Pacheco-Ruíz et al. 1998), which is a temperate area with lower sea surface temperature and higher nutrient content (Espinoza-Avalos 1993). Zones II and III were similar to other subtropical areas, such as Magdalena Bay (3.8 kg m-2 WW; Sánchez-Rodríguez and CervantesDuarte 1999). Zone I in 2006 is consistent with the 2 southern regions (8 and 9) studied by Pacheco-Ruíz et al. (1998) in 1995. The average biomass was 9.1 kg m-2 WW, higher than the earlier report (5.0 kg m-2 WW; Pacheco-Ruíz et al. 1998). This is between estimates of subtropical and temperate regions. The biomass in zone I is related to its transitional location between the north and central coastal areas of the peninsula (Espinoza-Avalos 1993). This area is dominated by upwelling currents in the Ballenas Channel, which is colder and more nutrient-rich than the southeastern coast (Álvarez Borrego 1983, Álvarez-Molina et al. 2013). According to Yeong et al. (2012), Sargassum species gradually increase in biomass with distance from the shore; in this zone Sargassum thalli can reach 25 m.
Halfar et al. (2004, 2006) measured chlorophyll a as a proxy for nutrients and temperature along the east coast of the peninsula (at 23oN, 25oN, 27oN, and 29oN) to relate these factors to carbonate production. Chlorophyll a increases northward: 0.3, 0.6, 1.6, and 2.2 mg m-3 at 23oN, 25oN, 27oN, and 29oN, respectively, which can explain the increase in biomass of Sargassum with increasing latitude.
The standing crop estimated in 2006 (this study) and in 1988 by Hernández-Carmona et al. (1990) and 1991 by Casas-Valdez et al. (1993) was lower than the standing crop calculated by Pacheco-Ruíz et al. (1998) along the northeastern coast of the peninsula, since the water temperature is cooler and the nutrient content is higher in this more northern area (Espinoza-Avalos 1993) than further south (Halfar et al. 2004, 2006).
Temporal comparisons
The difference in biomass in zone III between 1988 and 2006 could result from phenological changes in the beds caused by variations in environmental factors or a reduction of hard substrate surface. Further studies might reveal which factors lead to changes in biomass of Sargassum. Differences in standing crop between 1988 and 2006 are likely related to the disappearance of some important large beds in 2006, which yielded the lowest standing crop and lowest average biomass of the study.
Núñez-López and Casas-Valdez (1996) studied the phenological changes of Sargassum in Concepción Bay and they characterized 8 development stages (seedling, juvenile, young A and B, mature, and senescent A-C). They found that there are geographic and seasonal differences in the phenological stages of Sargassum. For instance, in spring 1991, the largest percentage of Sargassum thalli in bed 45 were young A (developing receptacles) and young B (developed receptacles) plants, while bed 32 contained mature thalli and bed 26 mostly contained senescent B stage thalli. During spring 1992, bed 45 mostly contained thalli at the young A and B stages, bed 32 contained thalli at the young A stage, and in bed 26 all thalli were juveniles. These 8 phenological stages are directly related to the quantities of fronds and, therefore, to the biomass of Sargassum (Núñez-López and Casas Valdez 1997). Differences in the biomass found in Sargassum beds at Concepción Bay between 1991 and 2006, and among the different beds in each year, can be the result of differences in the development stages of the Sargassum thalli at a specific time.
The main factor that affects the location of Sargassum is rocky substrata (Espinoza-Avalos 1993). We found that bed length was related to the surface area of hard substrata. The largest Sargassum beds occur at locations where a larger area is covered by stones, rocks, and/or pebbles (Casas-Valdez et al. 1993). The size of the beaches and hard substrate varies because they are exposed to seasonal and inter-annual dynamics, the first mainly due to wave energy (more intense in winter) and the second due to tropical storms and hurricanes. In the 3 zones, no significant coastal modifications or urbanization have occurred near the beds.
Our findings indicate that the standing crop of Sargassum in 3 zones had varied in 2006 relative to previous years. This variability must be considered to be able to achieve the sustainable exploitation of this abundant resource, which can be used as alginate source in many products, balanced diets, forage, etc. We recommend periodic monitoring of the beds to determine whether the resource is sustainable. The high variability in surface canopy, biomass, and standing crop of Sargassum beds is not related to anthropogenic activities because Sargassum beds have never been harvested along the east coast of the Baja California Peninsula.
Acknowledgments
Luis Aguilar Rosas and Raúl Aguilar Rosas† identified the Sargassum species. José Borges Souza, Ciro Arista de la Rosa, Martín Cuevas Higuera, Efraín Flores Montaño, Enrique Calvillo Espinoza, Javier Alvarez Espinoza, Jesús Espinoza Alvarez, and Pablo Simental Sigala conducted important field work. Ira Fogel (CIBNOR) provided extensive editorial services. MCV, ISR, and RNAR are EDI-IPN and COFAA-IPN fellows.

