










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de fitopatología
versión On-line ISSN 2007-8080versión impresa ISSN 0185-3309
Rev. mex. fitopatol vol.36 no.1 Texcoco ene./abr. 2018
https://doi.org/10.18781/r.mex.fit.1706-5
Review articles
The genus Bacillus as a biological control agent and its implications in the agricultural biosecurity
1 Instituto Tecnológico de Sonora, 5 de Febrero 818 Sur, Colonia Centro CP. 85000, Ciudad Obregón, Sonora, México.
2 Campo Experimental Norman E. Borlaug, CP. 85000, Ciudad Obregón, Sonora, México.
3 CONACYT-Instituto Tecnológico de Sonora, 5 de Febrero 818 Sur, Colonia Centro CP. 85000, Ciudad Obregón, Sonora, México.
The genus Bacillus is widely distributed in agro-systems, being one of its main applications the control of diseases in agricultural crops. The present review describes and analyzes the genus Bacillus, and its main mechanisms of action, such the excretion of antibiotics, toxins, siderophores, lytic enzymes and Induced systemic resistance, focused on its ability to be used as biocontrol agent of pests and diseases in plants; as well as its use in formulations of biopesticides, which have been incorporated into Integrated Pest Management programs. In addition, the Bacillus genus is analyzed in terms of agricultural biosecurity, as well as the principal criteria for the effective selection of biocontrol agents, considering strains not pathogenic to humans, and that do not negatively impact the microbial communities of agro-ecosystems, as a side-effect by its non-specific biological activity against a particular plant pathogen.
Key words: biocontrol; phytopathogens; biopesticides
El género Bacillus se encuentra ampliamente distribuido en los agro-sistemas y una de sus principales aplicaciones es el control de enfermedades de cultivos agrícolas. En la presente revisión se describe y analiza al género Bacillus, y sus principales mecanismos de acción, tales como la excreción de antibióticos, toxinas, sideróforos, enzimas líticas e induciendo la resistencia sistémica, enfocado en su capacidad para ser utilizado como agente de control biológico de plagas y enfermedades en plantas; así como su uso en la formulación de bioplaguicidas, que han sido incorporados a los programas de Manejo Integrado de Plagas y Enfermedades. Además, se analiza el uso del género Bacillus en la agricultura bajo un enfoque de bioseguridad agrícola, así como los principales criterios indispensables de selección de agentes de control biológico promisorios, considerando cepas no patogénicas para el ser humano, y que no impacten negativamente a las comunidades microbianas de los agroecosistemas, como efecto secundario por su actividad biológica no específica contra un fitopatógeno en particular.
Palabras clave: biocontrol; fitopatógenos; bioplaguicidas
Agriculture is a decisive activity in worldwide economic, social and environmental development, since it contributes to 80% of the food consumed globally (FAO, 2014). However, approximately 20 to 30% of the agricultural production is affected by pests and diseases, with fungi, bacteria, nematodes, viruses and insects being the main causal agents (Pérez-García et al., 2011). In this way, in order to fight diseases in crops, chemical compounds have been widely used, including insecticides, fungicides and nematicides. However, despite the success in the application of these compounds in the increase of worldwide agricultural activity, their excessive use has had a negative impact on soils, ecosystems and human health, i.e. captan, a fungicide used in Mexico, the use of which has been prohibited in other countries due to its carcinogenic effect (Arellano-Aguilar and Rendón, 2016).
On the other hand, the incidence of pests and diseases can be minimized by the use of cultural practices such as crop rotation and the system of planting in separate plots; however, these are not entirely efficient (Sainju et al., 2016). Thus, the Integrated Pest and Disease Management (IPDM) is a crucial alternative in the development of a sustainable agriculture that guarantees global food security, which uses the combination of biological, cultural and chemical methods in a compatible way (from Olivar et al., 2008). The IPDM highlights the use of biological control agents (BCA) as a sustainable alternative to mitigate the negative effects in the productivity and quality of the crops caused by different diseases, reducing the resistance of phytopathogenic organisms, and reducing the contamination of soils and water tables. This helps produce innocuous foods and reduces the costs of agricultural production (Reyes et al., 2015).
The use of BCA began in the early 19th Century, using live organisms or their metabolites to mitigate diseases in agricultural crops (Badii et al., 2000). In recent decades, the effect of a large diversity of rhizospheric microorganisms in the control of phytopathogenic organisms has been reported, since the rhizosphere represents the first line of defense of the plant against edaphic phytopathogenic organisms, thus avoiding their establishment in the roots (Tejera-Hernández et al., 2011). This has led to the study of multiple mechanisms used by the BCA to control the growth, development and infection of phytopathogenic organisms in diverse economically important crops; some of the most important of these mechanisms include: a) contact mechanisms such as hyperparasitism and predation (Chen et al., 2016); b) the production of compounds with low molecular weights with a direct effect on the growth of the antibiotic pathogen (phenazines, 2,4-diacetylfluoroglucinol, cyclic lipopeptides), lytic enzymes (chitnases, glucanases, proteases), products of unregulated residues (ammonia, carbon dioxide, hydroxide cyanide) (Yan et al., 2011; Zhang et al., 2016; Jaaffar et al., 2017; Piechulla et al., 2017) and c) indirect mechanisms from the competition for space and nutrients (consumption of leached-exudates, production of siderophores, induction to the systemic response in plants through the production of phytohormones and molecular patterns) (Pal and Gardener, 2006; Yu et al., 2011; Chowdhury et al., 2015). These action mechanisms have been observed in the microbial strains used as BCA, in which the Bacillus genus has been widely studied due to its high abundance and diversity in agro-systems (soil, water and plant), with a significantly higher population in comparison to other microbial genera, and also due to its diverse metabolic capabilities, with an outstanding capability to produce antibiotics and other antimicrobial and antifungal metabolites (Tejera-Hernández et al., 2011). For example, several molecular studies have revealed that a significant percentage of the genome of strains of the Bacillus genus is related to the production of secondary metabolites related to the control of phytopathogens, i.e. 8.5 and 4% of the genome of strains B. amyloliquefaciens FZB42 and B. subtilis 168, respectively (Raaijmakers and Mazzola, 2012). This justifies the wide use of strains of this bacterial genus as BCA for the control of diseases that affect agricultural crops. However, the efficiency of strains without risks to the biosecurity of the Bacillus genus used as BCA is maximized by their effective introduction to the agro-system, inducing their biological control mechanisms. Therefore, this study focuses on the analysis and discussion of the Bacillus genus, its main action mechanisms against plant diseases, as well as its implementation, without risks to biosafety with the development of biopesticides, since the scientific foundation of these bioproducts on its action mechanisms, environmental and biosafety implications, and the formulation of biopesticides are decisive for the development of a sustainable agriculture.
Classification
The Bacillus genus was first reported by Cohn (1872), who described it as heat-resistant, endospore-producing bacteria. The species of Bacillus belong to the bacteria kingdom; Phylus Firmicutes; Class Bacilli; Order Bacillales and Family Bacillaceae (Maughan and van der Auwera, 2011). Currently, the genus includes over 336 species, which, because of their genetic similarity, can be classified into different groups, the most important of which are a) the group of B. cereus, related to pathogenicity, which includes B. cereus-anthracis-thuringiensis; b) the environmental bacilli, which are characteristically present in different habitats, such as the group of Bacillus subtilis, composed of B. subtilis-licheniformis-pumilus; c) the group of B. clausii-halodurans; and d) the group that includes Bacillus sp. NRRLB-14911-coahuilensis (Alcaraz et al., 2010; LPSN, 2016).
Ecology
The Bacillus species are widely distributed worldwide due to their ability to form endospores, a characteristic that provides them with resistance and boosts its isolation in several habitats, both water and terrestrial ecosystems, and even in environments under extreme conditions (Tejera-Hernández et al., 2011). However, soils are considered the main reservoir of this bacterial genus, since most Bacillus species are saprophytic and are able to use the large diversity of organic substrates in the soil, making this a complex matrix for the establishment of a large genetic and functional diversity of microbial species (McSpadden, 2004). For this reason, the multiple Bacillus species can grow in soil, the cultivable counts are found in the range of log 3 to log 6 per gram of fresh weight of soil, essentially in genetically similar species to the group of B. subtilis and B. cereus (Vargas-Ayala et al., 2000). Regardless of rRNA studies in soil contradicting the relative abundance of cultivable and un-cultivable species of this bacterial genus (Kumar et al., 2011).
Under a focus aimed at agricultural sustainability, few investigations have aimed at understanding the specific population diversity and dynamics of Bacillus in rhizospheric soils, in which the bacterial communities that inhabit the rhizosphere respond particularly to the soil fertility and the exudates of plant roots, which vary with the phenology and genotype of the plant (de Souza et al., 2015), and therefore bacteria that interact with plants and present associative or endophytic capacities, or symbiotic processes to adapt to the rhizospheric conditions are acknowledged as potential microbial inoculants. Rudrappa et al. (2008) reported that the production of B. subtilis FB17 biofilm is a rhizosphere colonization mechanism, due to its attraction to the L-malic acid secreted by the roots of Arabidopsis thaliana and induced by the foliar pathogen Pseudomonas syringae pv tomato. On the other hand, Kumar et al. (2011), López-Fernández et al. (2016) and Selim et al. (2016) point out that diverse species of Bacillus may reside in internal tissues of grape (Vitis vinífera) and cotton plants (Gossypium barbadense L). These characteristics play a crucial role in the development, colonization and function of Bacillus, stimulating their relationship with the host plant, the biological control characteristics of which are strengthened.
Main characteristics
Some of the main characteristics of the Bacillus genus are its optional aerobic or sometimes anaerobic growth, Gram-positive, bacillary morphology, flagellar motility, and variable size (0.5 to 10 μm), its optimal growth being in a neutral pH, with a wide interval of growth temperatures, although most species are mesophilic (temperature between 30 and 45 °C), its metabolic diversity related to the promotion of plant growth and control of pathogens (Tejera-Hernández et al., 2011); other features that stand out are its capacity to produce endospores (oval or cyllindrical) as a mechanism of resistance to diverse types of stress (Calvo and Zúñiga, 2010; Layton et al., 2011; Tejera-Hernández et al., 2011) (Figure 1).
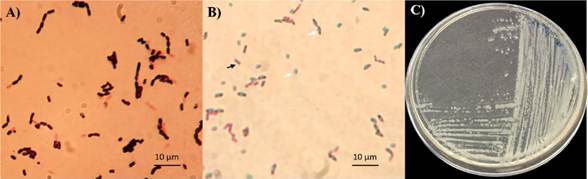
Figure 1 Morphological characteristics of Bacillus sp. TE3 belonging to the Collection of Native Edaphic and Endophytic Microorganisms (www.itson.edu.mx/COLMENA). A) Bacillary cells, Gram positive; B) endospores (white arrow) and bacillary cells (black arrow); and C) macroscopic morphology of strain TE3.
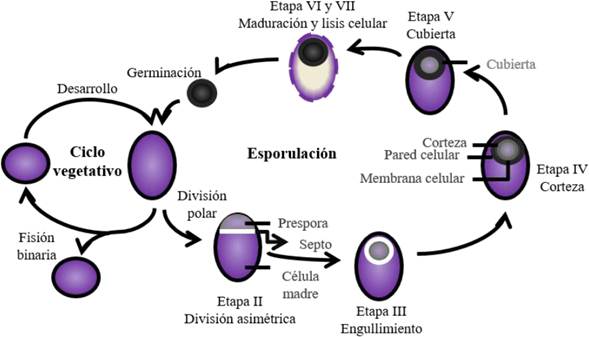
The presence of endospores confers the Bacillus genus its capacity to spread and its prevalence in ecosystems. These form during the second phase of its life cycle, which is composed of a phase of vegetative growth and a phase of sporulation (Figure 2). During the first stage, the bacteria grows exponentially by binary fission, since it is found in a medium with favorable conditions for its growth. The second phase begins as a strategy for survival in the presence of some type of stress (high population density, lack of nutrients, external factors such as salinity, temperature, pH, and others). In this way, the vegetative cell begins the formation of the endospore, which implies an asymmetric cell division, giving way to the formation of two compartments: a mother cell and the immersion of a prespore. Later, the prespore is swallowed, forming a cell within the mother cell. During the later stages, the prespore is covered by protective layers (protein components, peptidoglycan and a wall that resides below it formed by germ cells), followed by the dehydration and the final maturation of the prespore. Finally, the mother cell is lysed by programmed cell death, releasing the endospore. The endospore can remain viable in the environment until the conditions become favorable to begin their metabolic processes and generate a vegetative cell (Errigton 2003; Tejera-Hernández et al., 2011; CALS; 2016). Due to this, the formation of endospores resistant to heat and desiccation is an important characteristic for the formulation of biotechnological products based on strains of this bacterial genus (Pérez-García et al., 2011).
The Bacillus genus as a Biological Control Agent
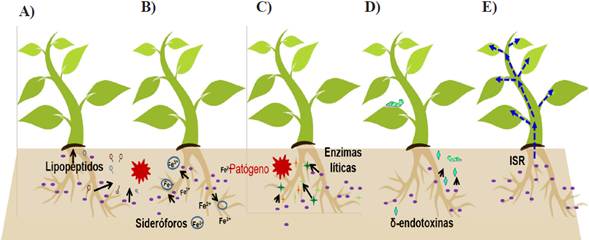
Large diversity of species of the Bacillus genus have proven to have antagonistic activity against diverse phytopathogenic microorganisms in agricultural crops, such as maize, rice, fruit trees, and others (Wang et al., 2014; Li et al., 2015). The study of this ability of Bacillus began with the discovery of insecticidal activity of Cry proteins produced by B. thuringiensis; currently, several species of the Bacillus genus (B. subtilis, B. pumilus, B. amyloliquefaciens and B. licheniformis) are widely studied to mitigate the incidence of diseases of importance to agriculture (Raaijmakers and Mazzola, 2012). Some of the main ways in which these strains avoid the establishment and development of phytopathogenic organisms is through different mechanisms, which include A) the excretion of antibiotics, B) siderophores, C) lytic enzymes, D) toxins and E) inducing the systemic resistance of the plant (ISR) (Figure 3) (Layton et al., 2011; Tejera-Hernández et al., 2011).
Production of lipopeptides
One of the most important characteristics of the Bacillus genus is its capacity to produce a large variety of antibiotics capable of inhibiting the growth of phytopathogenic agents, including non-ribosomal cyclic peptides, which have been the most widely studied. Lipopeptides (LPs) structurally consist of a cyclic peptide joined to a chain of β-hydroxy or β-amino fatty acids (Figure 4-A), classified into 3 different families (iturines, fengicines and surfactines), according to their amino acid sequences and length of the fatty acid (Ongena and Jaques, 2008; Falardeau et al., 2013). These molecules are synthesized by multienzymatic complexes, known as nonribosomal peptide synthetase (NRPS), which are independent to messenger RNA (Chowdhury et al., 2015). The families of iturines, fengicines and surfactines have been widely studied for their antibacterial and antifungal activity (Meena and Kanwar, 2015). The antimicrobial activity of these LPs occurs because of their interaction with the cytoplasmic membrane of bacterial or fungal cells, causing the formation of pores and an osmotic imbalance, which triggers the cell death of the phytopathogenic microorganisms (Figure 4-B) (Aranda et al., 2005; Gong et al., 2006). However, recent reports claim that some lipopeptides can have multiple action mechanisms, altering cell processes such as intracellular calcium homeostasis, energetic metabolism and RNA processing (Zhang et al., 2016).
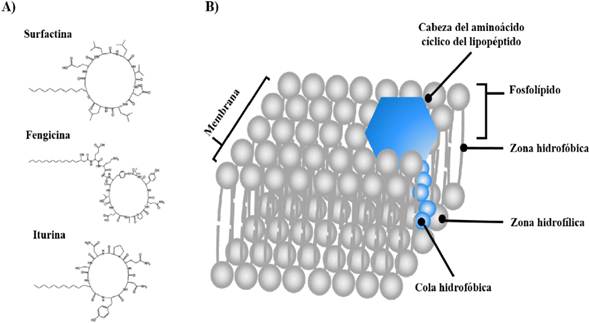
Figure 4 Lipopeptides as metabolites involved in the biological control of plant pathogens. A) Representatives of the family of lipopeptides, B) Action mechanism of lipopeptides. Modified from Ongena and Jaques (2008); Bayer (2016).
The role of the LPs has become evident in the growth inhibition in phytopathogenic microorganisms. Touré et al. (2004) identified several isoforms of lipopeptides in extracts produced from liquid cultures of B. subtilis GA1. Later, in a test for co-inoculation of strains GA1 and Botrytis cinerea in apple fruits, they identified the presence of fengicines and iturines in inhibitory concentrations, showing the activity of these lipopeptides in-situ. Likewise, genetic expression and mutagenesis analyses have contributed significantly to the understanding the role of lipopeptides in the microbial activity of the Bacillus genus against phytopathogenic microorganisms (Xu et al., 2013). For example, Li et al. (2014) mutated the gene of the phosphopantetheine transferase (sfp) in the strain SQR9 of B. amyloliquefaciens, crucial for the functioning of non-ribosomal peptide synthetase (NRPS), and therefore the synthesis of lipopeptides. This mutation resulted in a phenotype lacking antifungal activity against diverse agriculturally important phytopathogens, including Verticillium dahliae, Sclerotinia sclerotiorum, Fusarium oxysporum, Rhizoctonia solani, Fusarium solani, Phytophthora parasiticaThe efficient control of diseases by the biological control agents requires these to be able to establish in, and interact with, their host plant (Pérez-García et al., 2011). Likewise, along with antibiosis, lipopeptides have an influence on the establishment of Bacillus with the regulation of cell processes such as motility and the formation of biofilms (Choudhary and Jhori, 2009; Xu et al., 2013). The latter has been observed in the mutant of B. subtilis 6051, deficient in the production of surfactines, producing an irregular biofilm during colonization, as well as the reduction in its capacity to control infections caused by Pseudomonas syringae pv. tomato in Arabidopsis thaliana (Bais et al., 2004). Similarly, the role of lipopeptides in the motility of Bacillus has been proven with the study of deficient mutants in the production of lipopeptides, i.e. Bacillus subtilis 168 deficient of the gen sfp gene, displayed a loss in motility in semi-solid agar, although it was restored when purified lipopeptides were added to the culture medium (Kinsinger et al., 2003). Due to the role of the lipopeptides in several cell processes (antibiosis, formation of biofilms and motility), its isolated study on the motility in situ of Bacillus is complex.
Despite lipopeptides not being the only metabolites of the antibiome of the Bacillus genus involved in the biological control of phytopathogens, they have been proposed as the most efficient metabolites for this biological activity, due to its ecological role and antimicrobial capability, which explains their use in the search and selection of promissory BCA of this genus (Cawoy et al., 2015; Mora et al., 2015).
Production of lytic enzymes
The production of enzymes involved in the degradation of the cell wall of phytopathogenic agents is one of the most widely reported biological control mechanisms, in particular against pathogens of fungal origins. The fungal cell is made up of glycoproteins, polysaccharides and other components that vary according to the fungal species (Bowman and Free, 2006). The fraction of polysaccharides can comprise up to 80% of the cell wall of fungi, particularly chitin (~ 10 - 20%) and glucane (~ 50 - 60%), which are composed of residues of beta-1,3-glucose and beta-1,4-N-acetylglucosamine, respectively (Bowman and Free, 2006; Latgé, 2007). These polymers play a crucial role in the firmness of the cell wall by a wide network of glycosidic bonds. Therefore, the interference in these bonds can deteriorate the cell wall of phytopathogenic fungi, causing their lysis and cell death.
The production of lytic enzymes such as chitinases (EC 3.2.1.14) and β-glucanases (EC 3.2.1.6 and EC 3.2.1.39) excreted by the BCA, including the Bacillus genus, have displayed an inhibitory effect against pathogens of fungal origins (Compant et al., 2005). These enzymes are responsible for the degradation of the main polysaccharides that make up the cell wall of the fungi, by the hydrolysis of their glycosidic bonds (Figure 5). There are currently diverse scientific studies that report the role of these enzymes in the in vitro antifungal activities obtained from strains of the Bacillus genus (Kishore et al., 2005; Liu et al., 2010; Shafi et al., 2017). For example, Yan et al. (2011) demonstrated the role a chitinase in the control of Rhizoctonia solani by B. subtilis SL-13, evaluating the inhibitory activity of the purified enzyme, confronted with this pathogen. Chien-Jui et al. (2004) cloned the gene chiCW of B. cereus 28-9 on Escherichia coli DH5α and showed that the purified products presented a high inhibitory activity in the germination of Botrytis elliptica spores. Likewise, Martínez-Absalón et al. (2014) observed that when placing liquid B. thuringiensis UM96 supernatants in the specific inhibitor of chitinases, alosamidine, they lost their ability to inhibit the growth of Botrytis cinerea, thus showing the importance of chitinases on the biological control activity of the strain. Similarly, different authors have reported β-glucanases as important components in the activity for the biological control strains of the Bacillus genus; Aktuganov et al. (2007), reported that β-1,3-glucanases are the main lytic enzymes involved in the in vitro control of Bipolaris sorokiniana by Bacillus sp. 739, and in studies with B. amyloliquefaciens MET0908, with the aid of scanning electronic microscopy, the lytic activity of a β -1,3-glucanases on the Colletotrichum lagenarium hyphae was observed. Also, genetic engineering studies focused on the increase of antifungal activity of Bacillus strains have successfully achieved the insertion of codifying genes into lytic enzymes, such as gene ChiA of B. subtilis F29-3 in B. circulans, obtaining a more aggressive phenotype against Botrytis elliptica (Chen et al., 2004). In addition, Zhang et al. (2012) significantly increased the antifungal activity of Burkholderia vietnamiensis P418 in the face of different fungal phytopathogens by the chromosomal insertion of the gene Chi113, from a B. Subtilis strain.
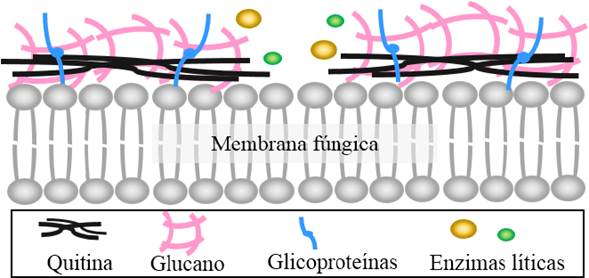
Figure 5 Degradation of the cell walls of plant pathogenic fungi by lytic enzymes. Modified from Moebius et al. (2014).
Production of siderophores
Iron (Fe) is an essential nutrient for important cell functions, such as in redox reactions of proteins with cofactors (Fe-S), in the electron transportation chain, and catalyzing vital enzyme reactions, such as those involving hydrogen, oxygen and nitrogen (Faraldo-Gómez and Sansom, 2003). Fe is found in nature mostly in a ferric form (Fe3+) with a low solubility, making its use impossible for some live beings (Aguado-Santacruz et al., 2012). In response to the iron restriction in the environment, some microorganisms have developed diverse receptor protein structures with low molecular weights and high affinity to iron, called siderophores, facilitating the capturing of Fe+3 (Thyagarajan et al., 2017). Siderophores are secondary metabolites that act as iron sequestrants or chelators, as a consequence of their high dissociation constant due to this metal, fluctuating between 1022 and 1055. This allows for the formation of Fe3+- siderophore complexes, so siderophore-producing microorganisms, using a specific receptor located in the membrane, can use it using two mechanisms: 1) directly with the Fe3+- siderophore complex through the cell membrane, or 2) reducing extracellularly to Fe2+ complexes (Neilands, 1995) (Figure 6-A). Siderophores, based on their main chelating group, are classified into: hydroxamates (using hydroxamic acids), catecholates (containing catechol rings), carboxylates, phenolates, and in a combination of two or more of these groups (Wilson et al., 2016) (Figure 6-B). A wide diversity of strains with the capacity of biological control belonging to the Bacillus genus have shown the ability to synthesize siderophores, regulating the concentration of iron in the medium through its chelation (Fe3+-siderophore), causing this metal to become unavailable for pathogenic microorganisms, which depend highly on this element for growth (Scharf et al., 2014). On the other hand, the formation of such complexes does not affect the development of the plants, since most of them can grow in lower iron concentrations than required by these biological control agents; likewise, some plants have the ability to use these microbial complexes, increasing their bioavailability to this element (Aguado-Santacruz et al., 2012).
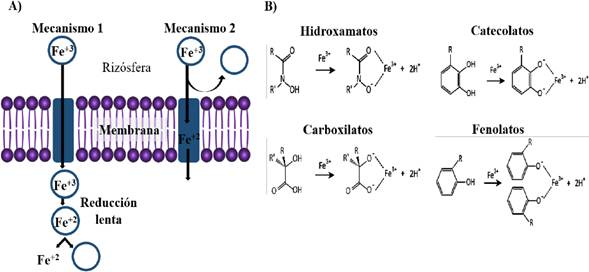
Figure 6 Siderophores as a mechanism for the inhibition of plant pathogens. A) Capture and solubilization of iron, B) main chelating structures of siderophores. Modified from Neilands (1995); Wilson et al. (2016).
In this way, several species of the Bacillus genus have been reported for their ability to control plant diseases by the secretion of siderophores, limiting the growth and colonization of iron-dependent phytopathogenic microorganisms (Fgaier and Eberl, 2011). Yu et al. (2011) showed, in co-culture tests in chrome azurol sulfonate (CAS) agar slides that B. subtilis CAS15 strongly antagonized the growth of 15 pathogenic fungi belonging to the genera of Fusarium, Colletotrichum, Pythium, Magnaporthe and Phytophthora, with inhibition rates in the interval of 19 to 94%, attributing this effect to the catecholate siderophores (Bacillibactin), identified by ESI-MS and DHB. Similarly, May et al. (2001) reported the potential of strains of the Bacillus genus on the production of siderophores by the analysis of genes involved in the synthesis of Bacillibactin (BB), particularly in B. subtilis, observing that the mutant strain JJM405 in gene dhb, presented a limited production of BB, while ATCC21332 (wild strain) presented a higher production of this siderophore.
Production of δ-endotoxins
δ-endotoxins, produced particularly by Bacillus thuringiensis (Bt), are protein parasporal bodies made up of polypeptide units of different molecular weights, from 27 to 140 kDa. There are currently 300 reported holotypes of Bt toxins, classified inti 73 Cry and 3 Cyt families (Porcar and Juárez, 2004; Xu et al., 2014). Bt toxins are produced during the sporulation phase; the Cry (crystal) protein is known for its toxic effects in an objective organism (most belong to the order of insects); likewise, Cyt (cytolitic) protein has been related to toxic effects on a large variety of insects, particularly diptera; however, its cytotoxicity has been tested on mammal cells (Soberón and Bravo, 2007).
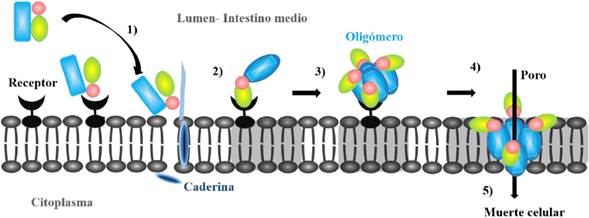
Cry proteins are widely used for their efficiency in the biological control of insects, the action mechanism of which begins once the Cry proteins are processes proteolytically through proteases present in the host’s middle intestine, separating a section of amino acids in the N-terminal region and in the C-terminal end (depending on the nature of the Cry protein), thus releasing active and toxic fragments that interact with the receptor proteins present in the intestine cells (Figure 7). These fragments are acknowledged by specific receptors in the membrane and inserted through the cadherin (1), giving rise to a series of signals for the formation of a pre-porous oligomeric structure (2-3), and consequently lytic porous (4), which triggers an osmotic imbalance, which destroys the intestinal epithelium and the consequent cell death (5) (Portela-Dussán et al., 2013; Xu et al., 2014). Several authors have reported the potential of Cry proteins in the toxicity of agriculturally important pests. For example, Niedmann and Meza-Basso (2006) point out that the tomato leafminer (Tuta absoluta) causes losses of between 60 and 100% of the crops that are not treated with insecticides, thus showing the potential of Bt, highlighting that in tests run on tomato leaves with added Cry protein concentrates extracted from strains LM-11, LM-12, LM-14 and LM-33 of Bt, mortality was reduced to between 20 and 60% of the T. Absoluta larvae, which would suggest the reduction in losses of up to 12 and 60%. On the other hand, Vázquez-Ramírez et al. (2015) highlight the prevailing role Bt strains can take against the pest of the fall armyworm (Spodoptera frugiperda), on biotests carried out with Cry protein extracts obtained from strains LBIT-13, LBIT-44, LBIT-383, LBIT-418 and LBIT-428 of Bt, showed that they all had a toxic effect on the fall armyworm, although strains LBIT-13 and LBIT-418 showed a high toxicity towards S. frugiperda, with CL50 of 137.2 and 197.2 ng cm-2, respectively, in comparison with the commercial standard HD-1 (CL50 of 142 ng cm-2).
Induced systemic response
Throughout evolutionary history, plants have developed mechanisms to defend themselves against the invasion of pathogenic organisms (bacteria, fungi, nematodes, insects, etc.). These mechanisms are latent and are activated by stimuli during the interaction with pathogenic agents. In general terms, these mechanisms are known as systemic acquired resistance (SAR) and become activated, not only in the site of the infection, but systemically in other tissues (Pieterse et al., 2014).
SAR is activated by stimuli perceived mainly by two receptors, the PRRs (pattern recognition receptors) and NB-LRRs (nucleotide-binding-leucine-rich repeat) (Pieterse et al., 2014). The former perceives cell components such as fungal chitin or flagellins (PAMPs o MAMPs, pathogen o microbe associated molecular patterns) triggering the first line of defense, known as PTI (PAMP-triggered immunity). In pathogens with mechanisms for the evasion of PTI, a second line of defense is activated, which perceives virulence effector proteins via receptors NB-LRRs (Boller et al., 2009). SAR depends on the signaling of salicylic acid (SA), which activates PR (pathogenesis-related) genes by codifying many of them to PR proteins with antimicrobial ability (e.i. PR1) (Vlot et al., 2009).
Similarly to SAR, the systemic response in plants may be induced (ISR, induced systemic resistance) by chemical signals (elicitors) produced by beneficial microorganisms (Pérez-Montaño et al., 2014) (Figure 3-E). Despite SAR and ISR being considered synonymous due to the similarity between the mechanisms (Pieterse et al., 2014), signaling in ISR depends on the jasmonic acid and ethylene (Pieterse, 1998The protection of agriculturally important crops (tomato, peppers, bean, rice, etc.) by inducing the systemic response using strains of the Bacillus genus has been documented (Akram et al., 2016; Choudhary and Johri, 2009; Wang et al., 2013; Yi et al., 2013). Bacillus produces a large diversity of eliciting molecules that induce a systemic response in plants, including lipopeptides (Chowdhury et al., 2015), phytohormones (Ryu et al., 2003) and volatile compounds (Kim et al., 2015). The latter activate PR genes, which protect from the invasion of pathogenic genes. This has been observed in tobacco plants, where PR2 codifies for one β-1,3 glucanase and PR3 codifies for one chitinase, they were activated in response to volatile compounds of Bacillus sp. JS, conferring resistance to Rhizocronia solani and Phytophthora nicotianae (Kim et al., 2015). Along with PR genes, Bacillus activates other plant protection mechanisms, which include structural changes in the cell wall with the accumulation of lignin (Singh et al., 2016) or the production or secondary metabolites such as flavonoids, phytoalexins, auxins or glucosinolates in general (Pretali et al., 2016). Despite the plant protection mechanisms induced by elicitors being activated by different metabolically routes, a microorganism can activate multiple protection mechanisms. This has been observed in wheat plants, where after the inoculation of Bacillus amyloliquefaciens B-16, the production of multiple PR proteins and secondary metabolites such as gallic acid and ferulic acid was induced, conferring resistance against Bipolaris sorokiniana (Singh et al., 2016).
The Bacillus genus and pesticides
The use of chemical pesticides as the main method for the control of pests and diseases in agriculture has helped increase agricultural productivity significantly in the past decades. However, their excessive use has created resistance to these compounds by phytopathogenic microorganisms, while having harmful effects on human health and the environment. This shows the need to generate efficient and environmentally friendly alternatives to reduce the use of synthetic products in order to achieve an efficient and sustainable control of diseases in agricultural crops (Pérez-García et al., 2011).
Biological control is an important part of the management of pests and diseases, and it consists of the use of living organisms to reduce and maintain the abundance of a pest or pathogen below the levels of economic damage. The potential of this alternative is based on an efficient control of a pest or disease in the middle and long terms, compatible with a low environmental risk and a sustainable production. In this way, the management of pests and diseases can be carried out by several methods: the use of synthetic pesticides, crops modified genetically to resist pests, biological control or the combination of one or more of these strategies. Biopesticides are a particular group of tools for the protection of crops used in the MIPE. Although there is not a formal definition for this term, a biopesticide refers to an agent produced at a massive scale, from a living microorganism or a natural product, and sold for the control of pests or diseases in plants (this definition covers most of the products classified as biopesticides, within the countries of the Organization for the Economic Co-operation and Development, OCDE, 2009). Biopesticides can be classified into three groups, depending on the active substance: i) biochemical products, that mainly comprise secondary metabolites of plants and microorganisms; ii) semi-chemical products, mostly made up of pheromones; and iii) microorganisms, which include bacteria, viruses, fungi, and protozoa (Chandler et al., 2011).
Of the commercially available microbial pesticides, Bacillus is the most widely exploited in agricultural biotechnology, with 85% of the bacterial products, due to its large metabolic versatility that allow it to perform a biological control of pests and diseases through diverse mechanisms. Also, this bacterial genus is able to produce endospores, which are the main active ingredient of the formulae, and confer to them - as a property - a greater viability in time (Ongena and Jaques, 2008).
One of the biological control mechanisms that has been most exploited in the biopesticides market is the ability of the Bacillus genus of producing δ-endotoxins. This mechanism was the milestone in the development of the first microbial biopesticide for the biological control of lepidoptera, produces from B. thuringiensis var. kurstaki HD-1. As mentioned earlier, B. thuringiensis (Bt) produces Cry proteins (Bt- δ endotoxins) during spore formation, which is capable of producing lysis in cells of the digestive tract when consumed by susceptible insects. Strain HD-1 is one of the most studied strains, since it characteristically carries varieties of cry antilepidoptera genes: cry1Aa, cry1Ab, cry1Ac, cry2Aa, cry2Ab and cry1Ia (Höfte and Whiteley, 1989; Sauka and Benintende, 2008).
Currently, B. Thuringiensis-based products account for 75% of the biopesticides sold globally (Olson, 2015), with a substantial impact on the Mexican biopesticide market since 1980. In Mexico, the use of Bt-based formulae is an efficient formula for the control of insects and it accounts for between 4 and 10% of the total of insecticides used for maize, cotton and vegetable crops (Tamez et al., 2001). Likewise, other species of the Bacillus genus, such as: B. subtilis, B. pumilus, B. licheniformis and B. amyloliquefaciens, stand out for their successful implementation in commercial formulations, and those developed mainly for the control of fungal diseases (Table 1). For example, Galindo et al., (2015) developed the first Mexican wide-spectrum biofungicide “Fungifree AB”, produced with viable B. subtilis 83 spores. This bacteria is a natural antagonist to diverse phytopathogens, used to prevent at least 8 pathogens of different etiologies: Colletotrichum, Erysiphe, Leveillula, Botrytis, Sphaerothecamacularis, in over 20 agriculture crops; They even indicate that the success of its formula resides, not only in the support provided by scientists, but also in the publication of its results in a journal read by professionals in agribusiness, thus allowing the linked that bonded crop exporting companies in search for sustainable alternatives that could allow them to control phytopathogens (i. e. Colletotrichum gloeosporioides).
Table 1 Bacillus as an active ingredient in commercial formulations.
| Agente de Control Biológico |
Producto (Año) |
Patógeno | Cultivos | Empresa |
| B. pumilus QST2808q | Ballad Plus (2007) | Erysiphe sp., Puccinia spp., Pyricularia sp., Rhizoctonia sp., Tilletia sp., Xanthomonas spp, entre otros. | Gramíneas, oleaginosas, entre otros. | AgraQuest |
| B. subtilis QST713r, s | Serenade ASO (2017) | Pythium spp., Rhizoctonia spp., Fusarium spp., Phytophthora sp., entre otros. | Frutales, hortalizas, entre otros. | Bayer CropScience |
| B. subtilis 83t | Fungifree AB (2012) | Colletotrichum sp., Leveillula sp., Botrytis sp., entre otros | Frutales, hortalizas | Agro & biotecnia |
| B. subtilis var. amyloliquefaciens FZB24u, v | Taegro 2 (2014) | Rhizoctonia sp., Fusarium sp., Pythium sp., Botrytis sp., entre otros. | Diversas frutas, plantas ornamentales, entre otros. | ISAGRO |
| B. licheniformis SB3086w, x | EcoGuard-GN (2013) | Colletotrichum sp., Sclerotinia sp., Rhizoctonia sp., entre otros. | Plantas ornamentales, entre otros. | Novozymes |
| B. thuringiensis var kurstakiy, z | DiPel WG (2007) | Cydia sp., Otiorhychus sp., Spodoptera sp, entre otros. | Frutales, hortalizas, entre otros. | Valent BioSciences |
(qAgraQuest, 2007; rBayer CropScience, 2016; sEPA, 2004; tGalindo et al., 2015; uISAGRO, 2017; vEPA, 2014; wNovozymes, 2017; xEPA, 2013; yValent BioSciences, 2017; zEPA, 2007).
In recent years, the chemical pesticide production market has declined 2% every year, while the production of biopesticides presents an annual increase of 20% (Cheng et al., 2010). There are several reasons for the increasing interest towards microbial biopesticides, including the limited development of resistance of pathogenic organisms to them, a reduction in the rate of discovery of new insecticides, a greater public perception of the dangers related to synthetic pesticides, the highest number of studies on the specificity of microbial pesticides, improvements in production, the technology of formulation and dissemination, as well as the interaction with producers and regulation bodies. In this way, biopesticides account for a small fraction of the global market focused on crop protection: 5% (approximate value of $3 billion USD). However, biopesticides are expected to have a Compound Annual Growth Rate (CAGR) of at least 8.64% by 2023, estimating a value of over $4.5 billion USD (Olson et al., 2013; Olson, 2015). It is worth mentioning that the transition and integration of the use of biopesticides in current agricultural practices should comply with the following requirements: a) effectiveness against the pest or disease; b) compatibility with other control methods; c) low or no environmental impact; d) long-lasting effect on the medium; e) economy, from a cost/ benefit viewpoint; f) technical feasibility of its use; and g) acceptance by producers and society in general. Therefore, the use of biopesticides offers an opportunity to stimulate the development and modernization of current agricultural practices, with the aim of contributing to food security under the scope of biosecurity.
Discussion and perspectives of biosecurity and biodiversity in the use of the Bacillus genus in agro-systems
The species of the Bacillus genus have a great metabolic and functional diversity, promoting its wide use in agriculture. In this sense, the groups of Bacillus cereus and Bacillus subtilis are the most widely used. However, in terms of biosecurity, they should be studied broadly before being used as biological control agents in the field. The group of Bacillus subtilis, which includes important species for agriculture, such as B. subtilis, B. licheniformis and B. pumilus, are not traditionally considered as pathogens for humans, and B. subtilis has even been granted the status of QPS (Qualified Presumption of Safety) by the European Food Security Authority (EFSA, 2015). However, there are some isolated cases of intoxications form digestive manifestations, such as the one reported by Pavic et al. (2005), pointing out B. subtilis and B. licheniformis as causal agents of the intoxication outbreak in a kindergarten caused by the consumption of powdered milk, which contained these bacterial species. On the other hand, in the group of Bacillus cereus made up of species such as B. cereus, B. thuringiensis, B. anthracis, B. mycoides, B. seudomycoides, B. cytotoxicus and B. weihenstephanensis, some strains have been identified as pathogens for humans (Ceuppens et al., 2013; Kim et al., 2016). Among these, B. cereus has been identified in a large diversity of foods (dairy products, fresh vegetables and others), causing important worldwide epidemiological crises, and in some cases, even death by emetic and diarrheal infections (Oh et al., 2012; Kim et al., 2016; Glasset et al., 2016). Also, B. thuringiensis strains have recently been related to intoxications caused by the ingestion of contaminated foods (Oh et al., 2012).
The virulence of these species has been mainly related to the presence of two toxins, hemolysin BL (HBL) and the nonhemolytic enteric toxin (NHE), which form a protein complex (Kim et al., 2016). Other toxins have also been identified in pathogenic strains, including cytotoxin K (cytK), enterotoxin FM (entFM), enterotoxin S (entS) and enterotoxin T (bceT). In strains that produce the emetic toxin (toxin of high resistance to thermal treatments, extreme pH values and the activity of proteases), virulence has been related to the presence of the dodeca depsipeptide synthesized by non-ribosomal peptide synthases (NRPS) codified by genes ces, found in type pXO1 plasmids. Likewise, products of other genes, such as hemolysin A (hlyA), hemolysin II and III (hlyI, hlyII), cereolysin A and B (cerA, cerB), and the pleitropic transcription factor (pclR) are involved in the pathogenicity of these strains (Ceuppens et al., 2013).
Historically, the classification and differentiation of species in the Bacillus cereus group has been carried out using gene 16S RNAr and other characteristics such as i) virulence (B. cereus), ii) content of plasmids (B. anthracis and B. thuringiensis), iii) growth conditions (B. cytotoxicus and B. weihenstephanensis) and iv) morphological characteristics (B. mycoides and B. seudomycoides). However, in widely related species such as B. cereus, B. anthracis and B. thuringiensis, the differentiation using virulence factors and contents of plasmids is limited, due to its loss and transference during the evolutionary history of these species (Hoffmaster et al., 2006; Liu et al., 2015). Recent comparative studies with complete genomes using dDDH (digital DNA: DNA hybridization) showed the distribution of cry genes and type pXO plasmids in members of this group, showing the low correlation between the phylogenetic position and the presence or absence of these plasmids (Liu et al., 2015). The above study also showed the low resolution of the multilocus sequence typing (MLST) for the differentiation at the level of species.
In this way, due to the high metabolic versatility with agricultural application shown by members of the B. Cereus group, particularly B. cereus and B. thuringiensis, the correct identification and the determination of its virulence for the human being is decisive for the selection and commercialization of biological control agents of these species. Although the study of comparative genomics using complete genomes is the only accurate alternative for the classification and determining its virulence, it is important to consider that they are costly tools for the discrimination of pathogenic strains during the primary process of selection of potential biological control agents.
On the other hand, in agro-systems, soil is a dynamic matrix that houses a large amount (~ 1x109 cells/gram of soil) and diversity (1x104 species/ gram of soil) of microorganisms (Curtis et al., 2002). This edaphic microbiota plays a very important ecological part, offering several eco-systemic services, such as i) social and ecological sustainability, ii) adaptation to, and mitigation of, climate change, iii) biotechnological resource for humanity, iv) cycling of water and nutrients, and v) food security, mainly by the cycling of nutrients (van der Heijden et al., 2008), and the boosting of plant growth through the production of phytohormones, solubilization of nutrients (Hayat et al., 2010) and avoiding the establishment of phytopathogenic agents (Compant et al., 2005). In this way, the use of biopesticides has acquired great relevance in the agricultural sector, offering a sustainable alternative, focused on increasing crop production. This generally implies the application of large populations of the microorganism of interest with the aim of promoting its establishment and colonization. However, this practice may cause disturbances in the microbial communities of the agro-systems (Trabelsi et al., 2013), particularly when inoculating biological control agents, since their biological activity is not specific or selective for the phytopathogenic agent in question, which may cause unpredictable changes in the microbial structure of said agro-systems. It is therefore important to evaluate the impact of the inoculation of biological control agents on the structure and composition of the microbial communities in agro-systems, to guarantee the ecological balance, as well as the desired biological effect.
Different studies have shown the impact of the inoculation of strains with the ability for biological control on microbial communities in different crops. For example, Li et al. (2015) evaluated the effect of the B. subtilis strain B068150 on the microbial communities in the rhizosphere of cucumber plants using the DGGE (denaturing gradient gel electrophoresis) technique. The study was carried out using three different soil types, with no significant changes in the microbial diversity related to the cucumber rhizosphere, after the inoculation of strain B068150. On the other hand, You et al. (2016) reported that with the inoculation of B. subtilis Tpb55, not only was the Phytophthora parasitica infection reduced, but significant changes were also observed in the microbial community related to the rhizosphere of a tobacco plantation, with an ANDRA (Amplified ribosomal 16S rDNA restriction analysis), highlighting that the relative abundance of some communities was favored, essentially in those belonging to the dominant fila: Acidobacteria and Proteobacteria, in an increase of 2 and 10% respectively in regard to the control, yet reducing by at least 50% the relative abundance of the communities belonging to Planctomycetes, Nitrospirae, Bacteroidetes and Chloroflexi, which may be involved in an important activity for plant development. In this way, each BCA is a particular organism that performs its action in a specific manner, in which the studies of each microbial strain chosen must be explored in depth in order to acquire further knowledge on the way to strengthen its biological control with an effective formulation, considering aspects of ecological risk and biosafety for the agro-system.
Conclusions
The negative impacts of chemical pesticides on the environment has been widely documented, include health damages, resistance to compounds by phytopathogens, soil and water pollution, and produce the need to develop sustainable alternatives to protect agricultural soils against pathogens, for example, biological control agents.
The Bacillus genus presents a large metabolic diversity involved in the biological control of phytopathogens; given this, the academic and industrial sectors have concentrated on generating commercial formulae for their use in the field, and in the description of the main action mechanisms involved in this effect (competition for space and nutrients, antibiosis, production of lytic enzymes, secretion of toxins, inducing host resistance). However, it is unusual for a single action mechanism to be used by said antagonist for the suppression of phytopathogens in situ. In this way, the knowledge of the mechanisms with which the antagonist exerts its action is decisive for both the guarantee of its effect and for the development of commercial formulations, the success of which lies in the creation of microenvironments that boost their biological activity without stimulating the growth of the pathogen. Currently, several commercial formulations consider strains of the Bacillus genus as an active ingredient, due to its ability to colonize, reproduce easily and its high persistence related to the formation of endospores, with the latter being a characteristic of interest, since it allows it to live under conditions of abiotic stress, making its production and long-term storage easier. On the other hand, after the inoculation of biopesticides, it is possible to observe a dual effect on crops due to the action of biological control agents, mitigating phytopathogens and indirectly promoting plant growth with the improvement of the plant’s health. However, it is necessary to carry out basic studies in depth, which integrate other MIPE strategies (cultural practices, chemical control), as well as in the correct identification of pathogenic strains for humans, such as Bacillus cereus NS Bacillus anthracis, which are even potentially effective strains for the control of phytopathogens. The potential risk of these species may be detected with taxonomy studies, β-hemolytic activity, detection with defined virulence molecular markers, guaranteeing the use of biologically safe strains for agriculture. In addition, studies on the ecological impact of the introduction of a BCA to agro-systems must be developed, since under certain circumstances, they can cause changes in the microbial communities with agro-ecologically unpredictable results. Finally, the success of the use of biopesticides will depend largely on the innovation, research, marketing strategies and dissemination of results amongst producers and the bodies in charge of making decisions related to regulation and the use of this type of technologies.
Acknowledgements
The authors would like to thanks and acknowledge the support given by Dr. Angélica Herrera for her suggestions for the improvement of this manuscript. The authors also wish to thank the National Science and Technology Council for the funding of the project 253663 “Fortalecimiento de la infraestructura del Laboratorio de Biotecnología del Recurso Microbiano del ITSON para la creación de COLMENA: Colección de Microrganismos Edáficos y Endófitos Nativos, para contribuir a la seguridad alimentaria regional y nacional (Fortifying the Infrastructure of the ITSON Microbial Resource Biotechnology Laboratory for the creation of COLMENA: Collection of Native Edaphic and Endophytic Microorganisms, to contribute to the regional and national food security)” and project 257246 “Interacción trigo x microorganismos promotores del crecimiento vegetal: identificando genes con potencial agro-biotecnológico (Interaction Wheat x Plant Growth Promoting Microorganisms: Identifying Genes with Agro-biotechnological Potential)”.
REFERENCES
AgraQuest. 2007. BALLAD PLUS, Biofungicide. Disponible en línea: http://fs1.agrian.com/pdfs/Ballad_Plus_(30_May_2007)_Label.pdf [ Links ]
Aguado-Santacruz GA, Moreno-Gómez B, Jiménez-Francisco B, García-Moya E y Preciado-Ortiz RE. 2012. Impacto de los sideróforos microbianos y fitosidéforos en la asimilación de hierro por las plantas: una síntesis. Revista Fitotecnia Mexicana. 35:9-21. Disponible en línea: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0187-73802012000100004 [ Links ]
Akram W, Anjum T and Ali B. 2016. Phenylacetic Acid Is ISR Determinant Produced by Bacillus fortis IAGS162, Which Involves Extensive Re-modulation in Metabolomics of Tomato to Protect against Fusarium Wilt. Frontiers in Plant Science. 7:1-12. https://doi.org/10.3389/fpls.2016.0049Aktuganov GE, Galimzyanova NF, Melent’ev AI and Kuz’mina L. 2007. Extracellular hydro-lases of strain Bacillus sp. 739 and their involvement in the lysis of micromycete cell walls. Microbiology. 76:413-120. http://dx.doi.org/10.1134/S0026261707040054 [ Links ]
Alcaraz LD, Moreno-Hagelsieb G, Eguiarte LE, Souza V, Herrera-Estrella L and Olmedo G. 2010. Understanding the evolutionary relationships and major traits of Bacillus through comparative genomics. BMC Genomics. 11:332. http://dx.doi.org/10.1186/1471-2164-11-332 [ Links ]
Aranda FJ, Teruel JA and Ortiz A. 2005. Further aspects on the hemolytic activity of the antibiotic lipopeptide iturin A. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1713:51-56. http://dx.doi.org/10.1016/j.bbamem.2005.05.003 [ Links ]
Arellano-Aguilar O y Rendón OJ. 2016. La huella de los plaguicidas en México. E. Martínez. Greenpeace México A. C. Las Flores 35 Col. Pueblo de Los Reyes, C.P. 04330, Coyoacán, México. 39 p. Disponible en línea: http://www.greenpeace.org/mexico/Global/mexico/Graficos/2016/comida-sana/Plaguicidas_en_agua_ok_EM.pdf [ Links ]
Badii MH, Tejada LO, Flores AE, Lopez CE y Quiróz H. 2000. Historia, fundamentos e importancia. Pp: 3-17. In: Badii MH, Flores AE y Galán LJ (eds.). Fundamentos y Perspectivas de Control Biológico. UANL, Monterrey. [ Links ]
Bais HP, Fall R and Vivanco JM. 2004. Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiology. 134:307-319. http://dx.doi.org/10.1104/pp.103.028712 [ Links ]
Bayer AG. 2016. Serenade, Fungicida Biológico, Mejorando la Protección de Cultivos. Bayer Crop Science. Folleto Serenade. Santiago, Chile. 5p. [ Links ]
Bayer CropScience. 2016. Serenade ASO. Disponible en línea: http://www.cropscience.bayer.cl/upfiles/etiquetas/SERENADE__ASO_20170517.pdf [ Links ]
Boller T and Felix G. 2009. A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annual Review Plant Biology. 60:379-406. http://dx.doi.org/10.1146/an-nurev.arplant.57.032905.105346 [ Links ]
Bowman SM and Free SJ. 2006. The structure and synthesis of the fungal cell wall. BioEssays 28:799-808. http://dx.doi.org/10.1002/bies.20441 [ Links ]
CALS, College of Agriculture and Life Sciences. 2016. Bacterial Endospores. Department of Microbiology. Cornell University. Ithaca, Nueva York 14850, EE. UU. Disponible en línea: https://micro.cornell.edu/research/epulopiscium/bacterial-endospores [ Links ]
Calvo P y Zúñiga D. 2010. Caracterización fisiológica de cepas de Bacillus spp. aisladas de la rizósfera de papa (Solanum tuberosum). Ecología Aplicada. 9:31-39. http://dx.doi.org/10.21704/rea.v9i1-2.393 [ Links ]
Cawoy H, Debois D, Franzil L, De Pauw E, Thonart P and Ongena M. 2015. Lipopeptides as main ingredients for inhibition of fungal phytopathogens by Bacillus subtilis/amyloliquefaciens. Microbial Biotechnology. 8:281-295. http://dx.doi.org/10.1111/1751-7915.12238 [ Links ]
Ceuppens S, Boon N and Uyttendaele M. 2013. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles. FEMS Microbiology Ecology. 84:433-450. https://dx.doi.org/10.1111/1574-6941.12110 [ Links ]
Chandler D, Bailey AS, Tatchell GM, Davison G, Graves J and Grant WP. 2011. The development, regulation and use of biopesticides for integrated pest management. Philosophical Transactions of the Royal Society B, Biological Sciences. 366:1987-1998. http://dx.doi.org/10.1098/rstb.2010.0390 [ Links ]
Chen CY, Wang YH and Huang CJ. 2004. Enhancement of the antifungal activity of Bacillus subtilis F29-3 by the chitinase encoded by Bacillus circulans chiA gene. Canadian Journal of Microbiology. 50:451-454. http://dx.doi.org/10.1139/w04-027 [ Links ]
Chen YY, Chen PC, Tsay TT. 2016. The biocontrol efficacy and antibiotic activity of Streptomyces plicatus on the oomycete Phytophthora capsica. Biological Control. 98:34-42. http://dx.doi.org/10.1016/j.biocontrol.2016.02.011 [ Links ]
Cheng XL, Liu CJ and Yao JW. 2010. The Current Status, Development Trend and Strategy of the Bio-pesticide Industry in China. Hubei Agricultural Sciences. 49:2287-2290. Disponible en línea: http://en.cnki.com.cn/Article_en/CJFDTOTAL-HBNY201009086.htm [ Links ]
Chien-Jui H, Tang-Kai W, Shu-Chung C and Chao-Ying C. 2004. Identification of an Antifungal Chitinase from a Potential Biocontrol Agent, Bacillus cereus 28-9. Journal of Biochemistry and Molecular Biology. 38:82-88. http://dx.doi.org/10.5483/BMBRep.2005.38.1.082 [ Links ]
Choudhary DK and Johri BN. 2009. Interactions of Bacillus spp. and plants with special reference to induced systemic resistance (ISR). Microbiological Research. 164:493-513. http://dx.doi.org/10.1016/j.micres.2008.08.007 [ Links ]
Chowdhury SP, Hartmann A, Gao X and Borriss R. 2015. Bio-control mechanism by root-associated Bacillus amyloliquefaciens FZB42 - a review. Frontiers in Microbiology. 6:780. https://doi.org/10.3389/fmicb.2015.00780 [ Links ]
Cohn F. 1872. Untersuchungen Über Bakterien. Beitrage zur Biologie Pflanz. 1:127-1224. [ Links ]
Compant S, Duffy B, Nowak J, Clément C and Barka EA. 2005. Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Applied Environmental Microbiology. 71:4951-4959. http://dx.doi.org/10.1128/AEM.71.9.4951-4959.2005 [ Links ]
Curtis TP, Sloan WT and Scannel JW. 2002. Estimating prokaryotic diversity and its limits. PNAS USA. 99:10494-10499. http://dx.doi.org/10.1073/pnas.142680199 [ Links ]
de Olivar CG, Castillo CCE, Cañizales BLM y Olivar R. 2008. Control Biológico: Una herramienta para el desarrollo sustentable y sostenible. Academia Trujillo Venezuela. 7:50-74. Disponible en línea: http://erevistas.saber.ula.ve/index.php/academia/article/view/6030/5831 [ Links ]
de Souza R, Ambrosini A and Passaglia LMP. 2015. Plant growth-promoting bacteria as inoculants in agricultural soils. Genetics and Molecular Biology. 38:401-419. http://dx.doi.org/10.1590/S1415-475738420150053 [ Links ]
Environmental protection agency, EPA. 2004. Office of Pesticide Programs Biopesticides and Pollution Prevention Division (7511C). Disponible en línea: https://www3.epa.gov/pesticides/chem_search/ppls/070127-00012-20140207.pdf [ Links ]
Environmental protection agency, EPA. 2004. United States environmental protection agency. Disponible en línea: https://www3.epa.gov/pesticides/chem_search/ppls/069592-00012-20040902.pdf [ Links ]
Environmental protection agency, EPA. 2007. Office of Pesticide Programs Biopesticides and Pollution Prevention Division (7611C). Disponible en línea: https://www3.epa.gov/pesticides/chem_search/ppls/000004-00252-20070112.pdf [ Links ]
Environmental protection agency, EPA. 2013. Office of chemical safety and pollution prevention. Disponible en línea: https://www3.epa.gov/pesticides/chem_search/ppls/070127-00003-20130906.pdf [ Links ]
Errigton J. 2003. Regulation of endospore formation in Bacillus subtilis. Nature Reviews Microbiology. 1:117-126. http://dx.doi.org/10.1038/nrmicro750 [ Links ]
European Food Safety Authority, EFSA. 2015. Statement on the update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA. 2: Suitability of taxonomic units notified to EFSA until March 2015. EFSA Journal. 13:4138. http://dx.doi.org/10.2903/j.efsa.2015.4138 [ Links ]
Falardeau J, Wise C, Novitsky L and Avis TJ. 2013. Ecological and mechanistic insights into the direct and indirect antimicrobial properties of Bacillus subtilis lipopeptides on plant pathogens. Journal of Chemical Ecology. 39:869-878. http://dx.doi.org/10.1007/s10886-013-0319-7 [ Links ]
FAO, Food and Agriculture Organization of the United Nations. 2014. El estado mundial de la agricultura y la alimentación. FAO. Viale delle Terme di Caracalla 00153 Roma, Italia. Disponible en línea: http://www.fao.org/3/a-i4040s.pdf [ Links ]
Faraldo-Gómez JD and Sansom MS. 2003. Acquisition of side-rophores in gram-negative bacteria. Nature Reviews Molecular Cell Biology. 4:105-116. http://dx.doi.org/10.1038/nrm1015 [ Links ]
Fgaier H and Eberl HJ. 2011. Antagonistic control of microbial pathogens under iron limitations by siderophore producing bacteria in a chemostat setup. Journal of Theoretical Biology. 273:103-114. http://dx.doi.org/10.1016/j.jtbi.2010.12.034 [ Links ]
Galindo E, Serrano-Carreón L, Gutiérrez CR, Balderas-Ruíz KA, Muñoz-Celaya AL, Mezo-Villalobos M y Arroyo-Colín J. 2015. Desarrollo histórico y los retos tecnológicos y legales para comercializar Fungifree AB®, el primer biofungicida 100% mexicano. TIP Revista Especializada en Ciencias Químico-Biológicas. 18:52-60. http://dx.doi.org/10.1016/j.recqb.2015.05.005 [ Links ]
Glasset B, Herbin S, Guillier L, Vignaud M, Grout J, Pairaud S and Ramarao N. 2016. Bacillus cereus -induced food-borne outbreaks in France, 2007 to 2014: epidemiology and genetic characterisation. Eurosurveillance. 21:1-11. https://dx.doi.org/10.2807/1560-7917.es.2016.21.48.30413 [ Links ]
Gong M, Wang JD, Zhang J, Yang H, Lu XF, Pei Y and Cheng JQ. 2006. Study of the Antifungal Ability of Bacillus subtilis Strain PY-1 in vitro and Identification of its Antifungal Substance (Iturin A). Acta Biochimica et Biophysica Sinica. 38:233-240. http://dx.doi.org/10.1111/j.1745-7270.2006.00157.x [ Links ]
Hayat R, Ali S, and Amara U. 2010. Soil beneficial bacteria and their role in plant growth promotion: a review. Annals of Microbiology. 60:579-598. http://dx.doi.org/10.1007/s13213-010-0117-1 [ Links ]
Hoffmaster AR, Hill KK, Gee JE, Marston CK, De B K, Popovic T, Sue D, Wilkins PP, Avashia SB, Drumgoole R, Helma CH, Ticknor LO, Okinaka RT and Jackson PJ. 2006. Characterization of Bacillus cereus isolates associated with fatal pneumonias: strains are closely related to Bacillus anthracis and harbor B. anthracis virulence genes. Journal of Clinical Microbiology. ca44:3352-3360. https://dx.doi.org/10.1128/jcm.00561-06 [ Links ]
Höfte H and Whiteley H. 1989. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiological Reviews. 53:242-255. Disponible en línea: http://mmbr.asm.org/content/53/2/242.long [ Links ]
ISAGRO. 2017. Isagro, TAEGRO 2, biofungicide - Product Training. Disponible en línea: http://www.isagro-usa.com/assets/taegro-2-training-presentation-usa-final-2017-05-02.pdf [ Links ]
Jaaffar AKM, Parejko JA, Paulitz TC, Weller DM and Thomashow LS. 2017. Sensitivity of Rhizoctonia Isolates to Phenazine-1-Carboxylic Acid and Biological Control by Phenazine-Producing Pseudomonas spp. Phytopathology. 107:692-703. http://dx.doi.org/10.1094/PHYTO-07-16-0257-R [ Links ]
Kim JS, Lee J, Lee CH, Woo SY, Kamg H, Seo SG and Kim SH. 2015. Activation of pathogenesis-related genes by the rhizobacterium, Bacillus sp. JS, which induces systemic resistance in tobacco plants. Plant Pathology Journal. 31:195-201. http://dx.doi.org/10.5423/PPJ. NT.11.2014.0122 [ Links ]
Kim MJ, Han JK, Park JS, Lee JS, Lee SH, Cho JI and Kim KS. 2016. Various Enterotoxin and Other Virulence Factor Genes Widespread Among Bacillus cereus and Bacillus thuringiensis Strains. Journal of Microbiology and Biotechnology. 25:872-879 https://dx.doi.org/10.4014/jmb.1502.02003 [ Links ]
Kim Y, Kim H, Kim K, Chon J, Kim D and Seo K. 2016. High Occurrence Rate and Contamination Level of Bacillus cereus in Organic Vegetables on Sale in Retail Markets. Foodborne Pathogens and Disease. 13:656-660. https://dx.doi.org/10.1089/fpd.2016.2163 [ Links ]
Kinsinger RF, Shirk MC and Fall R. 2003. Rapid surface motility in Bacillus subtilis is dependent on extracellular surfactin and potassium ion. Journal of Bacteriology. 185:5627-5631. Disponible en línea: http://jb.asm.org/content/185/18/5627.long [ Links ]
Kishore GK, Pande S and Podile AR. 2005. Biological Control of Late Leaf Spot of Peanut (Arachis hypogaea) with Chitinolytic Bacteria. Phytopathology. 95:1157-1165. http://dx.doi.org/10.1094/PHYTO-95-1157 [ Links ]
Kumar A, Prakash A and Johri BN. 2011. Bacillus as PGPR in Crop Ecosystem. Bacteria in Agrobiology: Crop Ecosystems. 37-59. http://dx.doi.org/10.1007/978-3-642-18357-7_2 [ Links ]
Latgé JP. 2007. The cell wall: a carbohydrate armour for the fungal cell. Molecular Microbiology. 66:279-290. http://dx.doi.org/10.1111/j.1365-2958.2007.05872.x [ Links ]
Layton C, Maldonado E, Monroy L, Corrales LC y Sánchez LC. 2011. Bacillus spp.; perspectiva de su efecto biocontrolador mediante antibiosis en cultivos afectados por fitopatógenos. Revista NOVA Publicación Científica en Ciencias Biomédicas. 9:177-187. http://dx.doi.org/10.22490/24629448.501 [ Links ]
Li B, Li Q, Xu Z, Zhang N, Shen Q and Zhang R. 2014. Response of beneficial Bacillus amyloliquefaciens SQR9 to different soilborne fungal pathogens through the alteration of antifungal compounds production. Frontiers in Microbiology. 5:636. http://dx.doi.org/10.3389/fmicb.2014.00636 [ Links ]
Li L, Ma J, Ibekwe AM, Wang Q and Yang C. 2016. Cucumber Rhizosphere Microbial Community Response to Biocontrol Agent Bacillus subtilis B068150. Agriculture. 6:1-15. https://dx.doi.org/10.3390/agriculture6010002 [ Links ]
Li Y, Gu Y, Li J, Xu M, Wei Q and Wang Y. 2015. Biocontrol agent Bacillus amyloliquefaciens LJ02 induces systemic resistance against cucurbits powdery mildew. Frontiers Microbiology. 6:883. http://dx.doi.org/10.3389/fmicb.2015.00883 [ Links ]
Liu D, Cai J, Xie C, Liu C and Chen Y. 2010. Purification and partial characterization of a 36-kDa chitinase from Bacillus thuringiensis subsp. colmeri, and its biocontrol potential. Enzyme Microbial Technology. 46:252-256. http://dx.doi.org/10.1016/j.enzmictec.2009.10.007 [ Links ]
Liu Y, Lai Q, Göker M, Meier-kolthoff JP, Wang M, Sun Y and Shao Z. 2015. Genomic insights into the taxonomic status of the Bacillus cereus group. Scientific Reports. 5:1-11. http://dx.doi.org/10.1038/srep14082 [ Links ]
López-Fernández S, Compat S, Vrhovsek U, Bianchedi PL, Sessitsch A, Pertot I and Campisano A. 2016. Grapevine colonization by endophytic bacteria shifts secondary metabolism and suggests activation of defense pathways. Plant and Soil. 405:155-175. http://dx.doi.org/10.1007/s11104-015-2631-1 [ Links ]
LPSN, List of Prokaryotic names with Standing in Nomenclature. 2016. Genus Bacillus. Microbiology Society. Charles Darwin House, 12 Roger St, London WC1N 2JU, United Kingdom. http://www.bacterio.net/bacillus.html (consulta, mayo 2017) [ Links ]
Martínez-Absalón S, Rojas-Solís D, Hernández-León R, Prieto-Barajas C, Orozco-Mosqueda MC, Peña-Cabriales JJ, Sakuda S, Valencia-Cantero E and Santoyo G. 2014. Potential use and mode of action of the new strain Bacillus thuringiensis UM96 for the biological control of the gray mold phytopathogen Botrytis cinerea. Biocontrol Science Technology. 24:1349-1362. http://dx.doi.org/10.1080/09583157.2014.940846 [ Links ]
Maughan H and van der Auwera G. 2011. Bacillus taxonomy in the genomic era finds phenotypes to be essential though often misleading. Infection, Genetics and Evolution. 11:789-797. http://dx.doi.org/10.1016/j.meegid.2011.02.001 [ Links ]
May JJ, Wendrich TM and Marahiel MA. 2001. The dhb operon of Bacillus subtilis encodes the biosynthetic template for the catecholic siderophore 2,3-dihydroxybenzoate-glycine-threonine trimeric ester bacillibactin. Journal of Biological Chemistry. 276:7209-7217. http://dx.doi.org/10.1074/jbc.M009140200 [ Links ]
Mc Spadden GBB. 2004 Ecology of Bacillus and Paenibacillus spp. in Agricultural Systems. Phytopathology. 94:1252-1258. http://dx.doi.org/10.1094/PHYTO.2004.94.11.1252 [ Links ]
Meena KR and Kanwar SS. 2015. Lipopeptides as the Antifungal and Antibacterial Agents: Applications in Food Safety and Therapeutics. BioMed Research International. 2015:1-9. http://dx.doi.org/10.1155/2015/473050 [ Links ]
Moebius N, Üzüm Z, Dijksterhuis J, Lackner G and Hertweck C. 2014. Active invasion of bacteria into living fungal cells. Microbiology and infectious disease. 1:1-20. http://dx.doi.org/10.7554/eLife.03007 [ Links ]
Mora I, Cabrefiga J and Montesinos E. 2015. Cyclic Lipopeptide Biosynthetic Genes and Products, and Inhibitory Activity of Plant-Associated Bacillus against Phytopathogenic Bacteria. PLoS One. 10: e0127738. http://dx.doi.org/10.1371/journal.pone.0127738 [ Links ]
Neilands JB. 1995. Siderophores: structure and function of microbial iron transport compounds. Journal of Biological Chemistry. 270:26723-26726. http://dx.doi.org/10.1074/jbc.270.45.26723 [ Links ]
Niedmann LL y Meza-Basso L. 2006. Evaluación de Cepas Nativas de Bacillus thuringiensis Como una Alternativa de Manejo Integrado de la Polilla del Tomate (Tuta absoluta Meyrick; Lepidoptera: Gelechiidae) en Chile. Agricultura Técnica. 66:235-246. http://dx.doi.org/10.4067/S0365-28072006000300002 [ Links ]
Novozymes. 2017. EcoGuard-GN, BioFungicide. Disponible en línea: http://www.kellysolutions.com/ere-newals/documentsubmit/KellyData%5CNC%5Cpesticide%5CProduct%20Label%5C70127%5C70127-3%5C70127-3_ROOTS_ECOGUARD_GN_BIOFUNGI-CIDE_12_9_2010_2_33_14_PM.pdf [ Links ]
OECD, Organization for Economic Co-operation and Development. 2009. Series on pesticides no. 44. Report of Workshop on the Regulation of Biopesticides: Registration and Communication Issues. Disponible en línea: http://www.oecd.org/env/ehs/pesticides-biocides/ENV-JM-MONO(2009)19-ENG.pdf [ Links ]
Oh, M., Ham, J. and Cox, J. M. 2012. Diversity and toxigenicity among members of the Bacillus cereus group. International Journal of Food Microbiology, 152:1-8. https://dx.doi.org/10.1016/j.ijfoodmicro.2011.09.018 [ Links ]
Olson S, Ranade A, Kurkjy N, Pang K and Hazekamp C. 2013. Green Dreams or Growth Opportunities: Assessing the Market Potential for “Greener” Agricultural Technologies. Lux Research Inc, Boston, MA, USA. Disponible en línea: https://portal.luxresearchinc.com/research/tidbit/15753 [ Links ]
Olson S. 2015. An analysis of the biopesticide market now and where it is going. Outlooks on Pest Management. 26:203-206. http://dx.doi.org/10.1564/v26_oct_04 [ Links ]
Ongena M and Jacques P. 2008. Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends Microbiolgy. 16:115-125. http://dx.doi.org/10.1016/j.tim.2007.12.009 [ Links ]
Pal KK and Gardener BM. 2006. Biological Control of Plant Pathogens. The Plant Health Instructor. 1:1-25. http://dx.doi.org/10.1094/PHI-A-2006-1117-02 [ Links ]
Pavic S, Brett M, Petric N, Lastre D, Smoljanovic M and Atkinson M. 2005. An outbreak of food poisoning in a kindergarten caused by milk powder containing toxigenic Bacillus subtilis and Bacillus licheniformis . Archiv Fur Lebensmittelhygiene. 56:20-22. Disponible en línea: https://www.tib.eu/en/search/id/BLSE%3ARN163847095/An-outbreak-of-food-poisoning-in-a-kindergarten/ [ Links ]
Pérez-García A, Romero D and de Vicente A. 2011. Plant protection and growth stimulation by microorganisms: biotechnological applications of Bacilli in agriculture. Current Opinion in Biotechnology. 22:187-193. http://dx.doi.org/10.1016/j.copbio.2010.12.003 [ Links ]
Pérez-Montaño F, Alías-Villegas RA, Bellogín RA, del Cerro P, Espuny MR, Jiménez-Guerrero I, López-Baena FJ, Ollero FJ and Cubo T. 2014. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiological Research. 169:325-336. http://dx.doi.org/10.1016/j.micres.2013.09.011 [ Links ]
Piechulla B, Lemfack MC and Kai M. 2017. Effects of discrete bioactive microbial volatiles on plants and fungi. Plant, Cell & Environment. 40: 2042-2067. http://dx.doi.org/10.1111/pce.13011 [ Links ]
Pieterse CMJ, Zamioudis C, Berendsen RL, Weller DM, Van Wees SC and Bakker PA. 2014. Induced Systemic Resistance by Beneficial Microbes. Annual Review of Phytopathology, 52:347-375. http://dx.doi.org/10.1146/annurev-phyto-082712-102340 [ Links ]
Pieterse CMJ. 1998. A Novel Signaling Pathway Controlling Induced Systemic Resistance in Arabidopsis. The Plant cell. 10:1571-1580. https://doi.org/10.1105/tpc.10.9.1571 [ Links ]
Porcar M y Juárez V. 2004. Aislamiento y establecimiento de una colección de Bacillus thuringiensis. Pp:69-100. In: Bravo A y Cerón J (eds). Bacillus thuringiensis en el control biológico. Universidad Nacional de Colombia, Bogotá. [ Links ]
Portela-Dussán DD, Chaparro-Giraldo A y López-Pazos SA. 2013. La biotecnología de Bacillus thuringiensis en la agricultura. Revista NOVA Publicación Científica en Ciencias Biomédicas. 11:87-96. http://dx.doi.org/10.22490/24629448.1031 [ Links ]
Pretali L, Bernardo L, Butterfield TS, Trevisan M and Lucini L. 2016. Botanical and biological pesticides elicit a similar Induced Systemic Response in tomato (Solanum lycopersicum) secondary metabolism. Phytochemistry, 130:56-63. https://doi.org/10.1016/j.phytochem.2016.04.002Raaijmakers JM and Mazzola M. 2012. Diversity and Natural Functions of Antibiotics Produced by Beneficial and Plant Pathogenic Bacteria. Annual Reviews of Phytopathology. 50:403-424. http://dx.doi.org/10.1146/annurev-phyto-081211-172908 [ Links ]
Reyes A, Ricón G, López L, Martínez ZE y Quiñones E. 2015. Lucha entre microbios: una herramienta para el control de enfermedades de plantas. Revista Digital Universitaria UNAM. 16:2-15. Disponible en línea: http://www.revista.unam.mx/vol.16/num11/art92/ [ Links ]
Rudrappa T, Czymme KJ, Paré PW and Bais HP. 2008. Root-Secreted Malic Acid Recruits Beneficial Soil Bacteria. Plant Physiology. 148:1547-1556. http://dx.doi.org/10.1104/pp.108.127613 [ Links ]
Ryu C-M, Hu C-H, Reddy MS and Kloepper JW. 2003. Different signaling pathways of induced resistance by rhizobacteria in Arabidopsis thaliana against two pathovars of Pseudomonas syringae. New Phytologist. 160:413-20. http://dx.doi.org/10.1046/j.1469-8137.2003.00883.x [ Links ]
Sainju UM, Lenssen AW, Allen BL, Stevens WB and Jabro J. 2016. Nitrogen balance in response to dryland crop rotations and cultural practices. Agriculture, Ecosystems and Environment. 233:25-32. http://dx.doi.org/10.1016/j.agee.2016.08.023 [ Links ]
Sauka DH y Benintende GB. 2008. Bacillus thuringiensis: generalidades. Un acercamiento a su empleo en el biocontrol de insectos lepidópteros que son plagas agrícolas. Revista argentina de microbiología. 40:124-140. Disponible en línea: http://www.scielo.org.ar/pdf/ram/v40n2/v40n2a13.pdf [ Links ]
Scharf DH, Heinekamp T and Brakjage AA. 2014. Human and Plant Fungal Pathogens: The Role of Secondary Metabolites. PLoS Pathogens. 10(1):e1003859. http://dx.doi.org/10.1371/journal.ppat.1003859 [ Links ]
Selim HMM, Gomaa NM and Essa AMM. 2016. Application of endophytic bacteria for the biocontrol of Rhizoctonia solani (Cantharellales: Ceratobasidiaceae) damping-off disease in cotton seedlings. Biocontrol Science and Technology. 27:81-95. http://dx.doi.org/10.1080/09583157.2016.1258452 [ Links ]
Shafi J, Tian H and Ji M. 2017. Bacillus species as versatile weapons for plant pathogens: a review. Biotechnology and Biotechnological Equipment. 31:446-459. http://dx.doi.org/10.1080/13102818.2017.1286950 [ Links ]
Singh UB, Malvivya D, Wasiullah, Singh S, Imran M, Pathak N, Alam M, Rai JP, Singh RK, Sarma Bk, Sharma PK and Sharma AK. 2016. Compatible salt-tolerant rhizosphere microbe-mediated induction of phenylpropanoid cascade and induced systemic responses against Bipolaris sorokiniana (Sacc.) Shoemaker causing spot blotch disease in wheat (Triticum aestivum L.). Applied Soil Ecology: 108:300-306. http://dx.doi.org/10.1016/j.aps-oil.2016.09.014 [ Links ]
Soberón M y Bravo A. 2007. Las toxinas Cry de Bacillus thuringiensis: modo de acción y consecuencias de su aplicación. Biotecnología. 14:303-314. Disponible en línea: http://www.ibt.unam.mx/computo/pdfs/libro_25_aniv/capitulo_27.pdf [ Links ]
Tamez GP, Galán WLJ, Medrano RH, García GC, Rodríguez PC, Gómez FRA y Tamez GGRS. 2001. Bioinsecticidas: su empleo, producción y comercialización en México. Ciencia UANL. 4:143-152. Disponible en línea: http://www.redalyc.org/pdf/402/40240205.pdf [ Links ]
Tejera-Hernández B, Rojas-Badía MM y Heydrich-Pérez M. 2011. Potencialidades del género Bacillus en la promoción del crecimiento vegetal y el control de hongos fitopatógenos. Revista CENIC Ciencias Biológicas. 42:131-138. Disponible en línea: http://www.redalyc.org/pdf/1812/181222321004.pdf [ Links ]
Thyagarajan SI, Ramanathan G, Singaravelu S, Kandhasamy S, Perumal PT and Sivagnam UT. 2017. Microbial Siderophore as MMP inhibitor: An interactive approach on wound healing application. Wound Medicine. 17:7-14. http://dx.doi.org/10.1016/j.wndm.2016.12.002 [ Links ]
Touré Y, Ongena M, Jacques P, Guiro A and Thonart P. 2004. Role of lipopeptides produced by Bacillus subtilis GA1 in the reduction of grey mould disease caused by Botrytis cinerea on apple. Journal of Applied Microbiology. 96: 1151-1160. http://dx.doi.org/10.1111/j.1365-2672.2004.02252.x [ Links ]
Trabelsi D and Mhamdi R. 2013. Microbial Inoculants and Their Impact on Soil Microbial Communities: A Review. BioMed Research International. ID 863240:1-11. http://dx.doi.org/10.1155/2013/863240 [ Links ]
Valent BioSciences. 2017. DiPel WG (Bacillus thuringiensis). Disponible en línea: http://www.cropscience.bayer.cl/upfiles/etiquetas/Dipel_WG.pdf [ Links ]
Van der Heijden M, Bardgett R and van Straalen N. 2008. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecology Letters. 11: 296-310. http://dx.doi.org/10.1111/j.1461-0248.2007.01139.x [ Links ]
Vargas-Ayala R, Rodríguez-Kábana R, Morgan-Jones G, Mclnroy JA and Kloepper JW. 2000. Shifts in Soil Microflora Induced by Velvetbean (Mucuna deeringiana) in Cropping Systems to Control Root-Knot Nematodes. Biological Control. 17:11-22. https://doi.org/10.1006/bcon.1999.0769 [ Links ]
Vázquez-Ramírez MF, Rangel-Nnúñez JC, Ibarra JE y del Rincón-Castro MC. 2015. Evaluación como agentes de control biológico y caracterización de cepas mexicanas de Bacillus thuringiensis contra el gusano cogollero del maíz Spodoptera frugiperda. Interciencia. 40:397-402. Disponible en línea: http://www.redalyc.org/pdf/339/33938675006.pdf [ Links ]
Vlot AC, Dempsey DA and Klessig DF. 2009. Salicylic acid, a multifaceted hormone to combat disease. Annual Review of Phytopathology. 47:177-206 http://doi.org/10.1146/annurev.phyto.050908.135202 [ Links ]
Wang W, Chen LN, Wu H, Zang H, Yang Y, Xie S and Gao X. 2013. Comparative proteomic analysis of rice seedlings in response to inoculation with Bacillus cereus. Letters in Applied Microbiology. 56(3):208-215. http://dx.doi.org/10.1111/lam.12035 [ Links ]
Wang X, Wang L, Wang J, Jin P, Liu H and Zheng Y. 2014. Bacillus cereus AR156-Induced Resistance to Colletotrichum acutatum Is Associated with Priming of Defense Responses in Loquat Fruit. PLos ONE. 9(11):e112494. http://dx.doi.org/10.1371/journal.pone.0112494 [ Links ]
Wilson BR, Bogdan AR, Miyazawa M, Hashimoto K and Tsuji Y. 2016. Siderophores in Iron Metabolism: From Mechanism to Therapy Potential. Trends in Molecular Medicine. 22:1077-1090. http://dx.doi.org/10.1016/j.molmed.2016.10.005 [ Links ]
Xu C, Wang BC, Yu Z and Sun M. 2014. Structural Insights into Bacillus thuringiensis Cry, Cyt and Parasporin Toxins. Toxins. 6:2732-2770. http://dx.doi.org/10.3390/toxins6092732 [ Links ]
Xu Z, Shao J, Li B, Yan X, Shen Q and Zhang R. 2013. Contribution of bacillomycin D in Bacillus amyloliquefaciens SQR9 to antifungal activity and biofilm formation. Applied Environmental Microbiology. 79:808-815. http://dx.doi.org/10.1128/aem.02645-12 [ Links ]
Yan L, Jing T, Yujun Y, Bin L, Hui L and Chun L. 2011. Biocontrol Efficiency of Bacillus subtilis SL-13 and Characterization of an Antifungal Chitinase. Chinese Journal Chemical Engineering. 19:128-134. http://dx.doi.org/10.1016/s1004-9541(09)60188-9 [ Links ]
You C, Zhang C, Kong F, Feng C and Wang J. 2016. Comparison of the effects of biocontrol agent Bacillus subtilis and fungicide metalaxyl - mancozeb on bacterial communities in tobacco rhizospheric soil. Ecological Engineering. 91:119-125. http://dx.doi.org/10.1016/j.ecoleng.2016.02.011 [ Links ]
Yu X, Ai C, Xin L and Zhou G. 2011. The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper. European Journal of Soil Biology. 47:138-145. http://dx.doi.org/10.1016/j.ejsobi.2010.11.001 [ Links ]
Zhang B, Qin Y, Han Y, Dong C, Li P and Shang Q. 2016. Comparative proteomic analysis reveals intracellular targets for bacillomycin L to induce Rhizoctonia solani Kühn hyphal cell death. Biochimica Biophysica Acta (BBA), Proteins and Proteomics. 1864:1152-1159. http://dx.doi.org/10.1016/j.bbapap.2016.06.003 [ Links ]
Zhang X, Huang Y, Harvey PR, Ren Y, Zhang G, Zhou H and Yang H. 2012. Enhancing plant disease suppression by Burkholderia vietnamiensis through chromosomal integration of Bacillus subtilis chitinase gene chi113. Biotechnology Letters. 34:287-293. http://dx.doi.org/10.1007/s10529-011-0760-z [ Links ]
Received: June 30, 2017; Accepted: December 15, 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
