










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de fitopatología
versión On-line ISSN 2007-8080versión impresa ISSN 0185-3309
Rev. mex. fitopatol vol.25 no.2 Texcoco 2007
Artículos científicos
Detección del Virus Mosaico Amarillo de la Calabaza Zucchini (ZYMV) y su Coinfección con otros Virus en Cucurbitáceas Cultivadas y Plantas Silvestres en el Valle del Fuerte, Sinaloa, México
Detection of Zucchini yellows mosaic virus (ZYMV) and its coinfection with other virus in cultivated Cucurbitaceae and wild plants in Fuerte Valley, Sinaloa, Mexico
Rubén Félix-Gastélum1, Marco Antonio Magallanes-Tapia2, Jesús Méndez-Lozano2, Hervé Huet3, José Ángel Trigueros-Salmerón1 y Rosa María Longoria-Espinoza4
1 Universidad de Occidente (U de O), Unidad Los Mochis, Depto. de Ciencias Biológicas (DCB), Blvd. Macario Gaxiola y Carr. Internacional s/n, Los Mochis, Sinaloa, México CP 81223. Correspondencia: rfelix@mochis.udo.mx.
2 CIIDIR-IPN, Unidad Sinaloa, Blvd. Juan de Dios Batiz Paredes 250, Guasave, Sinaloa, México CP 81101.
3 Bio-Oz. Ltd., Yad-Mordechai, Hof-Ashkelon 79145, Israel.
4 U de O, Unidad Guasave, DCB, Av. Universidad s/n, Guasave, Sinaloa, México CP 81120.
Recibido: Marzo 26, 2007
Aceptado: Junio 26, 2007
Resumen
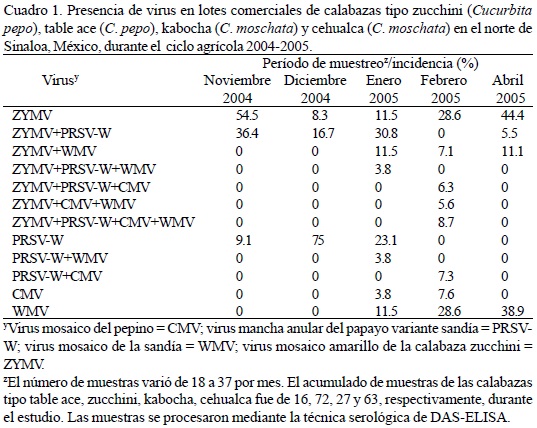
Las enfermedades de origen viral son el principal factor limitante en la producción de cucurbitáceas en el Valle del Fuerte, Sinaloa, México. El objetivo del presente estudio fue detectar al virus mosaico amarillo de la calabaza zucchini (ZYMV), virus mosaico de la sandía (WMV), virus mancha anular del papayo variante sandía (PRSV-W) y virus mosaico del pepino (CMV) mediante ELISA y RT-PCR. En el mes de noviembre predominaron ZYMV con 54.5% de incidencia y la mezcla ZYMV + PRSV-W con 36.4%; en diciembre ocurrieron el ZYMV y PRSV-W en 8.3 y 75%, respectivamente, y la mezcla ZYMV + PRSV-W ocurrió en 16.7%. En enero las infecciones por ZYMV, PRSV-W, CMV y WMV variaron de 3.8 a 23.1% y las mezclas de éstos oscilaron de 3.8 a 30.8%; en febrero predominaron el ZYMV y WMV con incidencias de 28.6% y CMV con 7.6%, las mezclas de éstos variaron de 5.6 a 8.7%; en abril, el WMV y ZYMV ocurrieron en 38.9 y 44.4%, mientras que las mezclas ZYMV+PRSV-W y ZYMV+ WMV ocurrieron en 5.5 y 11.1%. Plantas silvestres de tabaco (Nicotiana glauca), melón silvestre (Cucúrbita foetedissima), pepino amargo (Momordica charantia) y pepino espinoso (Cucumis dipsaseus) presentaron los mismos virus excepto ZYMV, el cual no se detectó en tabaco. Este trabajo abre nuevas líneas de investigación orientadas a la determinación del papel de las malezas en la epidemiología de los virus en la región.
Palabras clave: Enfermedades virales, tabaco, Nicotiana glauca, Cucurbita foetedissima, Momordica charantia, Cucumis dipsaseus.
Abstract
Viral diseases are the main limiting factor for cucurbit production in El Fuerte Valley, Sinaloa, Mexico. The objective of this study was to detect Zucchini yellows mosaic virus (ZYWV), Watermelon mosaic virus (WMV), Papaya ring spot virus (PRSV-W), and Cucumber mosaic virus (CMV) using ELISA and RT-PCR procedures. ZYMV and the mixture ZYMV + PRSV-W predominated during the month of November with 54.5 and 36.4% incidence, respectively. In December, infections by ZYMV and PRSV-W occurred at 8.3 and 75%, while ZYMV + PRSV-W was detected at 16.7%. In January, infections by ZYMV, PRSV-W, CMV, and WMV ranged from 3.8 to 23.1%; and the combinations of these viruses varied from 3.8 to 30.8%. In February, ZYMV and WMV occurred at 28.6%, while CMV occurred at 7.6%; combinations of these viruses ranged from 5.6 to 8.7%. In April, WMV and ZYMV reached 38.9 and 44.4%, respectively; the mixtures ZYMV + PRSV-W and ZYMV + WMV ocurred at 5.5 and 11.1%. All these viruses were detected in wild plants of tobacco (Nicotiana glauca), wild melon (Cucurbita foetedissima), bitter cucumber (Momordica charantia), and spiny cucumber (Cucumis dipsaceus), except ZYMV, which was not detected in wild tobacco. This study indicates the role of the wild plant species in the epidemiology these viruses merits further research in the region.
Keywords: Viral diseases, tabacco, Nicotiana glauca, Cucurbita foetedissima, Momordica charantia, Cucumis dipsaseus.
En el norte de Sinaloa, México, se siembra un promedio de 7,055 ha de cucurbitáceas anualmente (CAADES, 2004) e incluyen dos especies de calabaza (Cucurbita pepo L. y Cucurbita moschata Duch. ex Poir.), pepino (Cucumis sativus L.), melón (Cucumis melo L.) y sandía [Citrullus lanatus (Thumb.) Matsum. y Nakai]. Estos cultivos son afectados por enfermedades de origen fungoso y bacterial (León-Gallegos, 1988; Ramírez, 1991). Dentro de las primeras se encuentran el mildiú (Pseudoperonospora cubensis Berk. y Curt.) Rost., el tizón foliar por Alternaria cucumerina (Ellis y Everhart) Elliot, la mancha foliar inducida por Corynespora cassicola (Berk. y Curt.) Wei (Félix-Gastélum y Sánchez-Castro, 1980) y la cenicilla causada por Podosphaera (Sect. Sphaerotheca) xanthii (Castagne) U. Braun y N. Schishkoff (Félix-Gastélum et al., 2005) En relación a los virus, 32 de ellos han sido consignados en cucurbitáceas en el mundo (Lovisolo, 1980; Zitter et al., 1996.). Dentro de estos virus sobresalen por su importancia los que son trasmitidos por áfidos, particularmente el virus mosaico del pepino (CMV), virus mancha anular del papayo variante sandía (PRSV-W), virus mosaico de la sandía (WMV) y virus mosaico amarillo de la calabaza zucchini (ZYMV), los cuales son trasmitidos por 60, 25, 42, y 12 especies de áfidos (Homoptera: Aphididae), respectivamente (Kucharek y Purcifull, 2001). Estudios realizados en otros países indican que estos cuatro virus causan efectos severos en la producción y calidad de diversas cucurbitáceas (Blua y Perring, 1989; Castle et al., 1992; Desbiez y Lecoq, 1997; Lisa y Lecoq, 1984; Wang et al., 1992); por ejemplo, en los estados de Maranhao (Moura, et al., 2001) y Sao Paulo, Brasil (Yuki, et al., 2000), la incidencia del PRSV-W ha alcanzado el 64.4 y 49%, respectivamente. De igual forma, en África del Sur se ha consignado la incidencia del ZYMV, WMV y CMV en niveles de 50.6, 22.6 y 6.67%, respectivamente, encontrándose que el 30% de las plantas resultaron positivas para la presencia de más de uno de estos virus (Cradock et al., 2001). En Turquía, WMV, ZYMV y CMV han presentado incidencias de 53.9, 38.8, y 26.6%, respectivamente, en cucurbitáceas cultivadas (Sevik y Arli-Sokmen, 2003). Se ha observado que los síntomas inducidos por el ZYMV son más severos cuando ocurre en infecciones mixtas con otros virus, particularmente con CMV (Lecoq et al., 1981). Adicionalmente, en Hungría se ha detectado CMV, ZYMV y WMV en pepino, y su incidencia en el campo varía de un año al siguiente (Kiss et al., 2002). El PRSV-W y WMV se consignaron en Australia, donde el segundo ha prevalecido en C. pepo, C. maxima Duchesne ex Lam. y C. moschata (Greber, 1978), en tanto que ZYMV se reportó algunos años más tarde en el mismo país en un amplio grupo de cucurbitáceas cultivadas (Greber et al., 1987). Los virus en cuestión también se han detectado en malezas; así, el CMV, PRSV-W y ZYMV se reportaron en bule [Lagenaria siceraria (Molina) St.] pepino amargo (Momordica charantia L.) y pepino espinoso (Cucumis dipsaseus Eh.) (Ullman et al., 1991). Estudios subsiguientes indicaron la presencia de WMV y ZYMV en M. charantia en Costa Rica (Sánchez et al., 1998) y el CMV en Nicotiana glauca Graham. en Grecia (Volvas y Di Franco, 2004). Aún cuando las enfermedades virales en cucurbitáceas son limitantes económicas de gran importancia en el norte de Sinaloa, México, no se han realizado estudios sobre detección de virus en estos cultivos, y si bien existen, se realizaron hace varios años en el valle de Culiacán, Sinaloa, México, donde se determinó la presencia del CMV, virus mosaico de la calabaza (SqMV), WMV y PRSV-W (Nelson et al., 1966). Con base en lo anterior, se llevó a cabo el presente trabajo con el objetivo de detectar a CMV, WMV, PRSV-W y ZYMV asociados a mosaicos, amarillamientos, así como enanismo de plantas y deformación de hojas y frutos en cucurbitáceas cultivadas, tales como calabaza tipo table ace (Cucurbita pepo L.), kabocha (C. moschata), cehualca (C. moschata) y zucchini (C. pepo) y algunas malezas como M. charantia, C. dipsaseus, Cucurbita foetedissima Kunth. y N. glauca en el norte de Sinaloa, México.
MATERIALES Y MÉTODOS
Presencia de los virus ZYMV, CMV, WMV y el PRSV-W en lotes comerciales de cucurbitáceas cultivadas y plantas silvestres en el Valle del Fuerte. Para determinar la presencia de los virus se colectaron 178 muestras en 31 lotes comerciales de calabaza tipo table ace, kabocha, cehualca y zucchini (16, 27, 63 y 72 muestras de cada tipo, respectivamente) con síntomas presumiblemente de origen viral, en los Municipios de Ahome, Guasave y El Fuerte, Sinaloa, México. El material se colectó de noviembre de 2004 a abril de 2005 y el número muestras por mes varió de 18 a 37. Durante dicho período también se colectaron 5, 5, 3, y 39 muestras con síntomas de posible origen viral de pepino espinoso (C. dipsaseus), melón silvestre (C. foetedissima), pepino amargo (M. charantia) y tabaco silvestre (N. glauca), respectivamente.
Detección de ZYMV, PRSV-W, CMV y WMV por DAS-ELISA. Para el análisis de las muestras se utilizó la técnica serológica de inmunoadsorción ligada a enzimas en sándwich de doble anticuerpo (DAS-ELISA) (Clark y Adams, 1977). Las celdas de las placas (Dynatech, Alexandria, VA) se tapizaron en forma separada a partir de antisueros específicos para el ZYMV, PRSV-W, CMV y WMV proporcionados por el Dr. Hervé Huet (Bio-Oz. Ltd., Yad-Mordechai, Hof-Ashkelon 79145, Israel). A cada celda se le agregaron 100 µL savia obtenida de tejido sintomático; en el proceso se agregó un volumen igual de conjugado IgG-fosfatasa alcalina. Finalmente, se agregaron 100 µL de sustrato PNP diluido en buffer de sustrato a cada celda de la placa y se incubó en cámara húmeda a temperatura ambiente y en oscuridad. La absorbancia de la reacción antígeno-anticuerpo se registró en un lector Minireader II (Dyanatech laboratories, Inc., 900 Slaters Lane, Alexandria, Virginia, 22314) con una longitud de onda de 405 nm. Se consideró positiva la reacción, cuando el valor de la absorbancia fue tres veces más alto que la absorbancia del testigo negativo, el cual consistió en 2 celdas conteniendo solución de extracción y dos celdas adicionales con savia de calabaza o pepino sanos.
Extracción de RNA de plantas. Cuarenta y cinco muestras que resultaron positivas para la presencia de ZYMV por DAS-ELISA, también se analizaron mediante la técnica de RT-PCR, para la detección del mismo virus, recurriendo al siguiente procedimiento: 5 discos de tejido sintomático de 9 mm de diámetro de cada muestra se colocaron en tubos de microcentrífuga de 1.5 mL, luego se agregaron 400 µL de buffer de borato 0.5M pH 8.0, el tejido se maceró y se centrifugó por 6 segundos; 150 µL del sobrenadante se transfirieron a tubos de microcentrífuga de 1.5 mL y se centrifugaron a 13,000 rpm por 5 min; posteriormente, 40 µL del sobrenadante se transfirieron a otro tubo de 0.6 mL, al que se le agregaron 40 µL de PEG 6000 (12%) + NaCl (1%) y se reposó en hielo por 5 min, para centrifugarse a 10,000 rpm por 10 min; finalmente, la pastilla se resuspendió en 400 µL de buffer de borato 0.5M pH 8.0.
Detección de ZYMV por RT-PCR. El volumen total de la de reacción de RT fue de 10 µL, conteniendo 2.0 µL de solución amortiguadora AMV (5X), 0.5 del iniciador PR75 (AGG ATC CTG GGT AAT TC), 0.37 mM de dNTP's, 2.0 U/µL de inhibidor de RNAsa, 0.07 U/µL de AMV-RT, 3.43 µL de agua triple destilada estéril y 2.0 µL de RNA total del extracto de plantas sintomáticas. La mezcla de reacción se incubó en un termociclador DNA de Perkin Elmer CETUS serie P-7376 por 90 min a 46°C. Enseguida, las muestras se sometieron a amplificación por PCR. La mezcla final de reacción contenía un volumen total 20 µL, la cual consistió en 2.0 µL de buffer MMLV (5X), 1.5 mM de MgCl2 , 0.2 de los iniciadores PR151 (GTA GTG AAA CTA TCG TGC CAA ACA TGC) y PR75 (AGG ATC CTG GGT AAT TC), 0.2 µM de dNTPs , 0.03 U/µL de taq DNA polimerasa, 12.68 µL de agua triple destilada estéril y 2.0 µL del producto del RT (cDNA). La mezcla de reacción se incubó en el termociclador antes mencionado por 30 ciclos con una temperatura inicial de desnaturalización de 95°C por 2 min. Cada ciclo se programó para: 30 seg a 94°C, 30 seg a 56°C y 50 seg a 72°C; al finalizar los ciclos, se dio una extensión de 72°C por 3 min. Seis µL del producto de la PCR se analizaron por electroforesis con un voltaje constante de 77 V por 1.5 h en gel de agarosa al 1%. El producto se visualizó en un transiluminador.
RESULTADOS
Determinación de la presencia de virus en forma individual y en mezcla en lotes comerciales de cucurbitáceas en el valle del Fuerte, Sinaloa. Los síntomas consistieron en moteado, mosaico amarillo, malformación de hojas y enanismo de las plantas en general. En calabaza zucchini, fue frecuente la presencia de ampollamiento y agrietamiento de frutos. La incidencia de infecciones de virus en mezcla no permitieron discernir los síntomas inducidos por cada uno de ellos en particular. Durante el mes de noviembre se detectó al ZYMV, la mezcla ZYMV+PRSV-W y al PRSV-W en un 54.5, 36.4 y 9.1% de las muestras, respectivamente (Cuadro 1). Durante diciembre se encontró al ZYMV en un 8.3%, la mezcla ZYMV + PRSV-W en un 16.7% y al PRSV-W en un 75%. En enero se detectó el ZYMV en un 11.5%, mientras que la incidencia de las mezclas ZYMV+PRSV-W, ZYMV+WMV, ZYMV+PRSV-W+WMV y PRSV-W + WMV varió de 3.8 a 30.8%, al mismo tiempo la incidencia del PRSV-W en forma individual fue de 23.1%; de igual forma, por primera vez se detectaron los virus CMV y WMV infectando en forma individual, con incidencia de 3.8 y 11.5%, respectivamente. En febrero, la incidencia del ZYMV fue de 28.6%, no se encontró la mezcla ZYMV+PRSV-W, pero se encontró ZYMV + WMV con una incidencia del 7.1%; además se detectaron por única ocasión a las mezclas ZYMV+PRSV-W+CMV, ZYMV + CMV+WMV y ZYMV + PRSV-W+CMV+WMV con una incidencia de 6.3, 5.6 y 8.7%, respectivamente; a su vez se encontró por única ocasión a la mezcla PRSV + CMV con una incidencia de 7.3%; al mismo tiempo se encontró a CMV y WMV con incidencia de 7.6 y 28.6%, respectivamente. Los muestreos durante el mes de abril indicaron una incidencia de 44.4% para el ZYMV; nuevamente se detectó la mezcla ZYMV + PRSV con una incidencia del 5.5%, en tanto que la incidencia de la mezcla ZYMV + WMV fue de 11.1%; no se detectó al CMV sólo ni en mezcla con otros virus en esta fecha de muestreo; en tanto que la incidencia del WMV fue de 38.9%.
Detección de virus en plantas silvestres mediante el procedimiento de ELISA en el Valle del Fuerte, Sinaloa. De un total de 39 muestras de tabaco silvestre colectadas durante noviembre-abril, 13, 3 y 5 resultaron positivas para el CMV, PRSV-W y WMV, respectivamente; mientras que las mezclas CMV+WMV, PRSV-W+CMV y WMV + CMV+PRSV-W se detectaron en 2, 9 y 2 de las muestras, destacando por su mayor incidencia el CMV y la mezcla CMV+PRSV-W (Cuadro 2). De igual forma, de los ejidos La Arrocera, San Miguel y Flores Magón, se obtuvo un total de 13 muestras de cucurbitáceas silvestres; de ellas, 5 correspondieron a pepino espinoso y una fue positiva para ZYMV; un número igual de muestras de melón silvestre presentaron a PRSV y la mezcla ZYMV + PRSV-W, mientras que en tres de las muestras de pepino amargo se detectó PRSV-W, WMV y la mezcla de ambos (datos no mostrados).
Detección del ZYMV mediante RT-PCR en muestras de calabaza zucchini. Este procedimiento permitió detectar el ZYMV en el 100% de las muestras que resultaron positivas mediante el procedimiento de DAS-ELISA. La amplificación del fragmento esperado de 1069 pb, indicó la presencia de ZYMV en las muestras (Fig. 1).
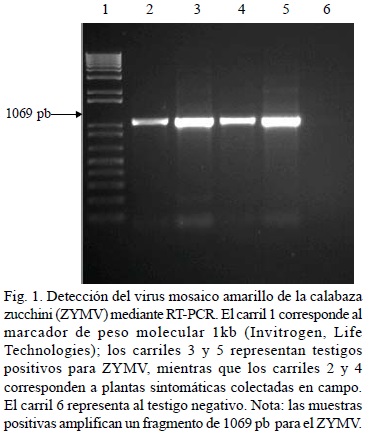
DISCUSIÓN
Los resultados indican que el ZYMV, CMV, PRSV-W y el WMV están presentes en cucurbitáceas cultivadas el norte de Sinaloa, México, como ocurre en otras zonas productoras de cucurbitáceas en el mundo (Blua y Perring, 1989; Castle et al., 1992; Cradock et al., 2001; Desbiez y Lecoq, 1997; Lisa y Lecoq, 1984; Wang et al., 1992), de estos virus, el ZYMV fue el más frecuente en infecciones simples o en mezcla con otros virus. La técnica de RT-PCR permitió la detección del ZYMV en muestras que previamente resultaron positivas mediante DAS-ELISA; dicha prueba permitió amplificar un producto de 1069 pb que corresponde a un fragmento del virus ZYMV reportado en Israel (Comunicación personal, Dr. Hervé Huet; Bio-Oz Ltd., Yad-Mordechai, Hof-Ashkelon). La alta sensibilidad y especificidad de la técnica de RT-PCR representan una ventaja con respecto a DAS-ELISA, pues permitirá la detección del ZYMV en vectores potenciales y en semilla de cucurbitáceas, donde el título de los virus en general es bajo. Esto a su vez contribuirá en la implementación de estrategias para el manejo de este virus. Resalta la detección de infecciones mixtas de hasta cuatro virus en una misma planta, como se ha consignado en Hawaii (Ullman et al., 1991) y Africa del Sur (Cradock et al., 2001). En esta ocasión, mezclas de virus fueron más frecuentes hacia el final del ciclo agrícola, lo que refleja una mayor disponibilidad de inóculo de los diferentes virus y altas poblaciones de insectos vectores; al mismo tiempo, se observó que la severidad de los síntomas consistentes en moteado, mosaico amarillo, malformaciones de hojas, ampollamiento y agrietamiento de frutos, así como enanismo de las plantas en general, particularmente en calabaza zucchini, fueron más evidentes al final del ciclo del cultivo lo que se puede deber a la presencia de más de un virus en las plantas, como ocurre en coinfecciones del ZYMV con el CMV (Lecoq et al., 1981). Aunque todas las muestras colectadas y procesadas por DAS-ELISA, mostraban síntomas presumiblemente de origen viral, no todas resultaron positivas para alguno de los virus analizados, pero podrían ser hospedantes naturales de otros virus no incluidos en este trabajo, pues se sabe que las cucurbitáceas pueden ser infectadas por más de 32 virus diferentes (Lovisolo, 1980; Zitter et al., 1996). Aún cuando se ha detectado a ZYMV, CMV, WMV y a PRSV-W en cucurbitáceas silvestres, tales como M. charantia, C. foetedissima y C. dipsaceus en Costa Rica (Sánchez et al., 1998), Florida (Adlerz, 1972) y Hawaii (Ullman et al., 1991), no se ha demostrado su importancia en la epidemiología de dichos virus. Estas malezas mostraron síntomas consistentes en mosaico y deformación de hojas (Figs. 2 y 3), excepto enM. charantia, en la que los síntomas no fueron visibles. De igual forma, la detección del CMV en tabaco silvestre, concuerda con estudios en Grecia (Vovlas y Di Franco, 2004), sin embargo, no se encontraron trabajos indicando la presencia del PRSV-W y el WMV en dicho hospedante. La detección de los virus, así como las mezclas de ellos en tabaco silvestre es relevante, pues esta especie crece en altas densidades principalmente en terrenos de aluvión abandonados, áreas limítrofes entre lotes agrícolas y a los costados de carreteras, caminos vecinales, vías de ferrocarril, así como en los bordos y taludes de canales de riego y drenes agrícolas; de igual forma, la presencia de estos virus además del ZYMV, en calabaza cehualca en huertos familiares de las comunidades ejidales de la región (datos no mostrados) reviste importancia, pues esta cucurbitácea está presente durante las cuatro estaciones del año, particularmente durante el verano y otoño, al igual que el tabaco silvestre, que representa una fuente potencial de inóculo. Con frecuencia esta cucurbitácea muestra síntomas de origen viral y su producción y calidad de la cosecha no son afectados en forma significativa; sin embargo, los virus detectados en esta especie causan pérdidas cuantiosas en otras especies de importancia económica. En el norte de Sinaloa, México, durante el verano y propiciado por las lluvias, abundan las cucurbitáceas y tabaco silvestres, así como la calabaza cehualca, las cuales se originan por germinación de semilla del año anterior o bien por el rebrotamiento en el caso de plantas perennes como ocurre en tabaco silvestre. Es importante resaltar que con frecuencia las cucurbitáceas silvestres, particularmente la calabaza cehualca, son colonizadas por especies no identificadas de pulgones, y considerando que estos virus pertenecen a los grupos de los potyvirus y cucumovirus, podrían ser diseminados por dichos insectos a lotes comerciales como ocurre en otras áreas productoras de cucurbitáceas del mundo (Kucharek y Purcifull, 2001). De los resultados del presente estudio se abren nuevas líneas de investigación, tales como el determinar si las especies predominantes de áfidos en la región actúan como vectores de virus presentes en las malezas antes señaladas. De igual forma, es importante determinar la presencia de otros virus que infectan a las cucurbitáceas cultivadas y la fluctuación poblacional de los vectores, así como la distribución espacio-temporal de los virus en la región. Esta información es fundamental para la implementación de estrategias de manejo de enfermedades de origen viral, donde se integren medidas de saneamiento (identificación y eliminación de hospedantes alternos), variedades resistentes, fechas de siembra y determinación de niveles de riesgo de incidencia de estas enfermedades en la diferentes zonas productoras de cucurbitáceas en el norte de Sinaloa, México.


CONCLUSIONES
El ZYMV fue el virus predominante en cucurbitáceas cultivadas en el norte de Sinaloa, México, durante el ciclo agrícola 2004-2005. La incidencia del ZYMV, CMV, WMV y el PRSV-W en forma individual e infecciones mixtas en cucurbitáceas cultivadas y malezas en el valle del Fuerte, Sinaloa, justifica la conducción de investigación sobre la ecología y epidemiología de estos virus; estos estudios permitirán el diseño de medidas apropiadas para el manejo de las enfermedades inducidas por estos virus en cucurbitáceas y otros cultivos de importancia económica a nivel regional.
LITERATURA CITADA
Adlerz, W.C. 1972. Momordica charantia as a source of watermelon mosaic virus I for cucurbit crops in palm beach County, Florida. Plant Disease Reporter 56:563-564. [ Links ]
Blua, M.J., and Perring, T.M. 1989. Effect of zucchini yellow mosaic virus on development and yield of cantaloupe (Cucumis melo). Plant Disease 73:317-320. [ Links ]
CAADES. 2004. Comparativo del Avance de Siembras de Hortalizas en las Temporadas 2002-2003 y 2003-2004. Documento interno del Departamento de Estudios Económicos, de la Confederación de Asociaciones Agrícolas del Estado de Sinaloa. Culiacán, México. s/p. [ Links ]
Castle, S.J., Perring, T.M., Farrar, C.A., and Kishaba, A.N. 1992. Field and laboratory transmission of watermelon mosaic virus 2 and zucchini yellow mosaic virus by various aphid species. Phytopathology 82:235-240. [ Links ]
Clark, M.F., and Adams, A.N. 1977. Characteristcs of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. Journal of General Virology 34:475-483. [ Links ]
Cradock, K.R., da Graca V.J., and Laing, M.D. 2001. Viruses infecting cucurbits in KwaZulu-Natal, South Africa. Revista Mexicana de Fitopatología 19:251-252. [ Links ]
Desbiez, C., and Lecoq, H. 1997. Zucchini yellow mosaic virus. Plant Pathology 46:809-829. [ Links ]
Félix-Gastélum, R., Apodaca-Sánchez, M.A., Martínez-Valenzuela, M.C. y Espinosa-Matías, S. 2005. Podosphaera (sect. Sphaerotheca) xanthii (Castagne) U. Brawn N. Shishkoff en Cucurbitáceas en el Norte de Sinaloa, México. Revista Mexicana de Fitopatología 23:162-168. [ Links ]
Félix-Gastélum, R. y Sánchez-Castro, M.A. 1980. Estudio epifitiológico y combate químico de la mancha foliar del pepino causada por Corynespora cassicola (Berk. & Curt.) Wei, en el Valle de Culiacán, Sinaloa, México. Memorias del IX Congreso Nacional de la Sociedad Mexicana de Fitopatología. Uruapan, Michoacán. México. 4 p. [ Links ]
Greber, R.S. 1978. Watermelon mosaic virus 1 and 2 in Queensland cucurbit crops. Australian Journal of Agricultural Research 29:1235-1245. [ Links ]
Greber, R.S, Mclean, G.D., and Grice, M.S. 1987. Zucchini yellow mosaic virus in three states of Australia. Australasian Plant Pathology 16:19-21. [ Links ]
Kiss, E., Kazinczi, G., Horváth, J., Kobza, S., Baranyi, T., Varga, M., Havasréti, B., and Fehér, A. 2002. Virus disease problems on field cucumber in Hungary with some international aspects. Acta Phytopathologica et Entomologica Hungarica 37:317-327. [ Links ]
Kucharek,T., and Purcifull, D. 2001. Aphid-transmitted viruses of cucurbits in Florida. Florida Cooperative Extension Service. Institute of Food and Agricultural Sciences. Circular 1184. University of Florida. Gainesville, Florida, USA. 14 p. [ Links ]
Lecoq, H., Pitrat, M., and Clement, M. 1981. Identification, et caractérisation d'un potyvirus provoquant la maladie du rabougrissementjaune du melon. Agronomie 1:827-834. [ Links ]
Lisa, V., and Lecoq, H. 1984. Zucchini yellow mosaic virus. In: B.D. Harrison, and A.F. Murant (eds.). Description of Plant Viruses. Commonwealth Mycological Institute and Association of Applied Biologist. Slough, Inglaterra. No. 282. [ Links ]
León-Gallegos, H.M. 1988. Enfermedades de Cultivos en el Estado de Sinaloa. Tercera edición. CAEVACU-CIAPAN. INIFAP. Culiacán, Sinaloa, México. 262 p. [ Links ]
Lovisolo, O. 1980. Virus and viroid deseases of cucurbits. Acta Horticulturae 88:33-71. [ Links ]
Moura, C.C.L., Lima, J.A.A., Oliveira, V.B. y Goncalves, F.B. 2001. Identificacao serológica de espécies de virus que infectam cucurbitáceas em áreas produtoras do Maranhao. Fitopatologia Brasileira 26:90-92. [ Links ]
Nelson, M.R., Laborde, J.A., and McDonald, H.H. 1966. Cucurbit viruses on the west coast of Mexico. Plant Disease Reporter 50:947-950. [ Links ]
Ramírez, V.J. 1991. Enfermedades de las Hortalizas. Primera Edición. Universidad Autónoma de Sinaloa. Culiacán, Sinaloa, México. 105 p. [ Links ]
Sánchez, M.V., Aguero, R. y Rivera, C. 1998. Plantas hospederas de los virus mas importantes que infectan el melón (Cucumis melo) en Costa Rica. Revista de Biología Tropical 46:13-25. [ Links ]
Sivik, M.A., and Arli-Sokmen, M.2003. Virus infecting cucurbits in Samsun,Turkey. Plant Disease 87:341-344. [ Links ]
Ullman, D.E., Cho, J.J., and German, T.L. 1991.Occurrence and distribution of cucurbit viruses in the Hawaiian Islands. Plant Disease 75:367-370. [ Links ]
Volvas, C., and Di Franco, A. 2004. Cucumber mosaic virus in Nicotiana glauca in Greece. Journal of Plant Pathology 86:91-92. [ Links ]
Wang, H.L., Gonsalves, D., Providenti, R., and Zitter, T.A. 1992. Comparative biological and serological properties of four strains of zucchini yellow mosaic virus. Plant Disease 76:530-535. [ Links ]
Yuki, V.A., Rezende, J.A.M., Kitajima, E.W., Barroso, P.A.V., Kuniyuki, H., Groppo, G.A., and Pavan, M.A. 2000. Occurrence, distribution and relative incidence of five viruses infecting cucurbits in the state of Sao Paulo, Brazil. Plant Disease 84:516-520. [ Links ]
Zitter, T.A., Hopkins, D.L., and Thomas, C.E. (eds.). 1996. Compendium of Cucurbit Diseases. APS Press. St. Paul, Minnesota, USA. 87 p. [ Links ]

