











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkLos hábitos alimentarios en lagartijas son considerados una característica de historia de vida, ya que del alimento ingerido se obtiene la energía utilizada para el crecimiento, reproducción y regeneración de tejidos (Ramírez-Bautista, 1995; Gadsden & Palacios-Orona, 2000). Las especies del género Aspidoscelis son consideradas insectívoras y generalistas, pues consumen gran cantidad de artrópodos de las clases Arachnida e Insecta; sin embargo hay cierta especialización en el consumo de un tipo de presa en particular durante la temporada de lluvias. En esta temporada del año, se ha reportado un consumo del 80 al 90% de isópteros (termitas) en A. marmorata, A. tesselata y A. tigris, para la región centro del Desierto Chihuahuense (Gadsden & Palacios-Orona, 2000; Mata-Silva et al., 2013); mientras que en A. tesselata, (Taylor et al., 2011), determinaron un alto consumo de escarabajos (73%) y larvas de lepidópteros (84%) en la región sureste de Colorado en Estados Unidos. En el centro de México existe una gran diversidad de especies de Aspidoscelis (Wilson et al., 2013), algunas presentan distribución restringida y otras amplia, y se ha considerado que la distribución de algunas especies depende de la abundancia de un determinado tipo de presas como las termitas (Güizado-Rodríguez, 2012). A. gularis, es una especie con amplia distribución, se encuentra desde el oeste de Oklahoma hasta el sur de Texas en Estados Unidos, entra a México por Tamaulipas y se extiende a los estados de Chihuahua, Durango, Coahuila, Zacatecas, San Luis Potosí, Nuevo León, Querétaro, Guanajuato, Aguascalientes, Jalisco, Hidalgo y Veracruz (Smith & Taylor, 1950; Pérez-Almazán et al., 2014); es común encontrarla en diversos tipos de vegetación como bosque de coníferas, bosque de encino, bosque tropical caducifolio, tropical subcaducifolio, pastizales y matorral xerófilo (Wilson et al., 2013).
El presente trabajo describe la dieta de A. gularis en una parte de la temporada de lluvias (junio-julio) de 1982. Se revisaron 19 machos (LHC ¯X = 68.2, 52-80.3 mm) y 12 hembras adultos ( ¯X = 60.0, 51.5-74.1 mm) de la localidad ubicada a 2.63 km al este y 0.67 km al norte (20° 57´ 44.17" N, 99° 30´ 45.45" W, Datum WGS84; 1796 msnm) de El Apartadero, municipio de San Joaquín, en la región sureste del estado de Querétaro, México. El clima de esta localidad es subhúmedo con lluvias en verano, con un intervalo de temperatura media anual que va de los 16 a los 20 °C, y una precipitación mínima de 600 y una máxima de 1000 mm anuales (INEGI, 1986). Los ejemplares revisados (Machos: CRD-929, 931, 932, 934, 935, 941, 943, 944, 945, 946, 948, 950, 951, 954, 955, 956, 957, 959, 960. Hembras: CRD-930, 936, 939, 940, 942, 947, 949, 952, 953, 958, 961, 962) se encuentran depositados en la Colección de anfibios y reptiles del CIIDIR Unidad Durango perteneciente al Instituto Politécnico Nacional.
A cada ejemplar se le realizó una incisión en la región ventral para remover el tubo digestivo, el cual fue pesado en una balanza granataria a una precisión de 0.1 g. Se extrajo todo el contenido estomacal, el cual fue colocado y expandido en una caja Petri forrada en su base con una cuadrícula de 1 mm; dicha cuadrícula ayudó a conocer el largo y ancho del total del tubo digestivo, así como de cada tipo de presa (Luria-Manzano 2013). Todos los insectos (enteros y semi-digeridos) fueron identificados a nivel de orden con ayuda de las claves para insectos de (Triplehorn & Johnson, 2005), únicamente el orden Hymenoptera fue dividido en dos clases, formícidos y no formícidos.
Con ayuda de la fórmula de una elipsoide (V = 4/3 π (largo/2) (ancho/2)2 se obtuvo el volumen de cada tipo de presa (Duré et al., 2009), y con la fórmula de (Biavati et al., 2004) (I = (F% + N% + V%)/3) se obtuvo el índice de importancia alimentaria (IIA) para cada tipo de presa, donde F es el porcentaje de ocurrencia, N es el porcentaje numérico y V el porcentaje volumétrico. Por otro lado, para conocer la diversidad de presas consumidas por machos y hembras, se utilizó el índice de Shannon-Weaver, calculado con la fórmula: H'= -∑ pi ln (pi ), donde pi es la proporción del recurso i (número de tipos de presas) en la dieta, y ln es el logaritmo natural (Shannon & Weaver, 1949). Al valor obtenido de H´se le aplicó la función D = exp (H´) para conocer la diversidad verdadera, que establece como unidad de medida el número de presas efectivas consumidas por cada sexo (Shannon & Weaver, 1949). Finalmente, para reconocer el solapamiento de los tipos de presas consumidos por ambos sexos se utilizó el índice de solapamiento alimentario de Pianka (Pianka, 1973).
Los machos se alimentaron de 12 tipos de presas (Cuadro 1), de los cuales las termitas fueron el grupo más importante en su dieta (IIA = 48.05%), en segundo lugar estuvieron los coleópteros adultos (IIA = 19.7%) y en tercero las larvas de coleóptero (IIA = 8.84%), el valor de diversidad de los tipos de presas consumidas por los machos fue H´= 2.6. Por su parte, las hembras se alimentaron de un menor número de presas (10 tipos de presas; Cuadro 1), entre los grupos más consumidos se encuentran las termitas (isópteros) (IIA = 44.09%), en segundo lugar los coleópteros adultos (IIA = 15.6%) y en tercero las cucarachas (Blattodea) (IIA = 11.52%), junto con las hormigas (Hymenoptera) (IIA = 11.26), el valor de diversidad de las presas consumidas por las hembras fue de H´= 4.3. Finalmente, el solapamiento alimentario para machos y hembras respecto al consumo de presas fue alto (97%), indicando amplias semejanzas en las presas consumidas por ambos sexos.
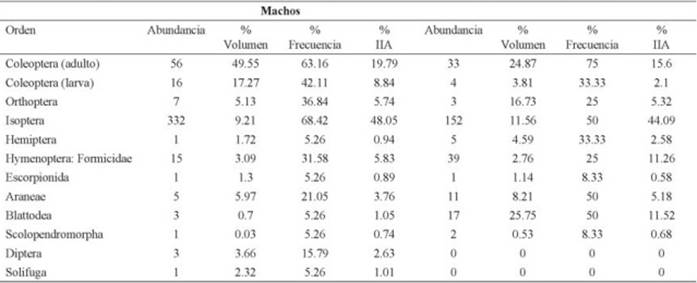
Cuadro 1: Ordenes de insectos que componen la dieta de machos y hembras de Aspidoscelis gularis durante la temporada de lluvias de la localidad El Apartadero, municipio de San Joaquín, Querétaro, México. Índice de importancia alimentaria (IIA).
Nuestros resultados coinciden con lo reportado por (Gadsden & Palacios-Orona, 2000) para A. tigris, y por (Güizado-Rodríguez & Casas-Andreu, 2011) para A. lineatissima, quienes observaron un importante consumo de termitas en la dieta de estas especies, alcanzando promedios de consumo mayores al 50%. Se ha mencionado que algunas especies de Aspidoscelis distribuidas en bajas altitudes son especialistas en el consumo de termitas (Güizado-Rodríguez, 2012), tal como fue mencionado en A. hyperythrus sobre un rango de elevación de 200 a 900 m, consumiendo entre 60 y 90% de termitas (Vázquez-Reyes 2006), en A. sacki a una elevación de 1500 m, consumiendo cerca del 30% de este mismo grupo de invertebrados (Aldape-López et al., 2009), y A. calidipes (Raya-García et al., 2015) a una elevación de 200 a 600 m, reportándose un consumo del 80% de termitas. Sin embargo, este patrón no es consistente con A. gularis ya que los individuos revisados en este trabajo se encuentran cerca de los 2000 m de elevación, y hace suponer que el consumo de termitas no siempre está correlacionado con la distribución de algunas especies de este género.
El segundo grupo más consumido por machos y hembras fue el de los coleópteros adultos, el tercer grupo más consumido por machos fueron las larvas de coleóptero y para las hembras fueron las cucarachas y hormigas, lo que revela que en lluvias pueden ser los grupos más consumidos, y por lo tanto el recurso más aprovechado por ambos sexos, lo que resulta en un alto grado de solapamiento alimentario. Por último, el índice de diversidad de Shannon fue más alto en hembras que en machos, debido a que el número de presas consumidas por cada hembra fueron más heterogéneas que en los machos; el índice de solapamiento alimentario de Pianka resultó elevado, debido al consumo casi idéntico de los tipos de presas entre sexos. Es necesario desarrollar trabajos que revelen el aporte energético del grupo más consumido (termitas) por machos y hembras en lluvias como en secas, de esta forma se podrá determinar si el alto consumo de una presa les reditúa mayor energía o solo es consumido por ser la más abundante en una determinada estación del año.

