










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex no.85 Xalapa abr. 2002
Article
Comparison of rodent communities in sites with different degrees of disturbance in deciduous forest of southeartern Morelos, Mexico
Carlos García-Estrada1, Ma. de Lourdes Romero-Almaraz1, and Cornelio Sánchez-Hernández2
1 Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de Morelos, Av. Universidad 1001, C. P. 62210, Col. Chamilpa, Cuernavaca, Morelos, MÉXICO.
2 Instituto de Biología, Universidad Nacional Autónoma de México. Dept. Zoología, Ap. Postal 70-153. C. P. 04510, México, D. F. MÉXICO.
Recibido: 26 de marzo 2001
Aceptado: 20 de septiembre 2001
Resumen
Este estudio es el primer trabajo que compara comunidades de roedores en una selva baja caducifolia en México. Señala las diferencias entre dos sitios con diferentes grados de alteración ocasionado por la extracción de madera y leña, y el pastoreo del ganado; en un sitio relativamente poco alterado (Sitio 1) y otro sitio más alterado (Sitio 2) en el sureste del estado de Morelos. En cada sitio se capturaron seis especies de roedores. Aunque la alteración del hábitat no modificó la diversidad o la equitatividad de las especies de roedores, el número total de inviduos capturados en el Sitio 1 (n = 319) fue mayor que en el Sitio 2 (n = 90). Los efectos de la fragmentación de hábitat se expresaron en diferencias significativas en la densidad poblacional, estructura de edades y preferencia en el microhábitat entre los dos de sitios para Baiomys musculus, Peromyscus levipes y P. melanophrys. La alteración de la selva baja caducifolia parece ser el principal factor que explica las diferencias en los parámetros demográficos de las especies entre los dos sitios, debido a que los dos sitios tienen el mismo clima, el suelo es similar, y la topografía difiere ligeramente.
Palabras clave: comunidades de roedores, alteración, diversidad, ecología poblacional, selva baja caducifolia, México.
Abstract
This study is the first work that compares rodent communities in a deciduous forest in Mexico. It documents differences between sites experiencing different degrees of disturbance caused by firewood and lumber extraction, and cattle grazing; a relatively undisturbed site (Site 1) and another more disturbed site (Site 2) in southeastern Morelos State. In each site we captured six species of rodents. Though habitat disturbance did not modify diversity or evenness of rodent species, the total number of individuals captured in Site 1 (n = 319) was greater than in Site 2 (n = 90). Effects of habitat fragmentation were expressed in significant differences in population density, age structure and microhabitat preference between two sites for Baiomys musculus, Peromyscus levipes and P. melanophrys. The disturbance of deciduous forest appears to be the principal factor explaining differences in demographic parameters of rodent species between the two study sites, as these two sites have the same climate, similar soils, and topography which differ only slightly.
Key Words: rodent communities, disturbance, diversity, population ecology, deciduous forest, Mexico.
Introduction
Stability of ecological communities is dependent upon the inherent balance of their component species populations (Chapman & Reiss 1992, Putman 1994; Stiling 1999). Ecological disturbances caused human activities as agriculture, forestry and urbanitation commonly affect community composition, structures and processes (Matson et al. 1997, Noble & Dirzo 1997, Pickett & White 1985). Disturbance also reduces the average size of patches of a habitat and increases the distance between them. Such changes can reduce animal population densities, as well as dispersion to new areas, thus increasing population instability and the probability of local extinction (Terborgh 1992, Herkert 1994, Waiters et al. 1994, Johannesen & Ims 1996, Bowers & Matter 1997). With increasing habitat fragmentation, one of the outcomes is the reduction in the number and size of suitable patches in a region, this increases the probability of local extinction (Moilanen & Hanski 1995), by lowering animal densities and not permiting the recolonization of patches. Due to these and other factors associated with disturbances, populations and communities in fragmented landscapes are not likely to be in overall equilibrium (Wiens 1985).
Several studies have concluded that intermediate levels of disturbances may enhance species diversity by lowering the dominance of one or a few species (Connell 1980, Terborgh & Winter 1980, Kirkland 1989) or by increasing environmental heterogeneity (Tilman 1982, Yahner 1991). However, if disturbance occurs frequently, richness can decreases because species intolerant to disturbance become locally extinct (Denslow 1985, Adler & Levins 1994, Collins et al. 1995, Williams & Marsh 1998) or can be maintained since species are tolerant to disturbance (Jonson et al. 1979, Anthony et al. 1981, Huntly & Inouye 1987). While the longer-term impacts of disturbance on population, community and ecosystem-level processes are still under debate, the immediate effects of habitat fragmentation are more clear, following habitat fragmentation species richness of mammals frequently declines dramatically producing large shift in species composition of the entire faunal assemblage. While many resident species disappear with habitat fragmentation, other tolerant or exotic species arrive in these areas to exploit the disturbed areas left vacant (Laurance 1991, Laurance & Yensen 1991, Nupp & Swihart 1998). Even among native species changes in dominance, diversity and evenness may occur as a result of habitat fragmentation (Kirkland 1990, Holt 1993).
There are not many information exists on rodent community dynamics in deciduous forest in Mexico (Collett et al. 1975, Ceballos 1990), and studies that compare demographic characteristics of species occupying fragmented and unfragmented habitats are exceedingly rare (Medellín 1992, Fa & Sánchez-Cordero 1993, Martínez Gallardo 1995, Vázquez et al. 2000). Our objective was to address this problem by examining the effects of disturbance caused by firewood and lumber extraction, as well as the cattle grazing, on rodent diversity, evenness, community and population dynamics.
We hypothesized that species diversity and evenness of rodents should be greater in a relatively undisturbed site (Site 1) versus a disturbed site (Site 2). Similarly, as forest fragmentation produces a biotic isolation for rodents within deciduous forests, and has been documented that habitat fragmentation can be species-specific, we expect that Baiomys musculus, Peromyscus levipes and P. melanophrys which habitat requirements are specialized (Baker and Greer 1962, Packard and Montgomery 1978) will be intolerant species to disturbance and their population sizes will reduced in Site 2. Furthermore, we expected that demographic parameters (population density, sex ratio, age structure, residence) of those rodent species in Site 1 to be greater than in Site 2. Meanwhile Liomys irroratus will be a tolerant species to disturbance and its demographic parameters should be similar in both sites because it is adapted to arid regions, has affinities for edge habitats and disturbed sites and well-developed vagility (Dowler & Genoways 1978). Finally, we predict that preferences for microhabitats in four species will be modified due to differences in the availability of resources as food and refuges between the two sites.
Materials and methods
Study sites
Our study was conducted at 3 km north of the Ejido El Limón (18°31'10"-18°31'50"N and 98°56'30"-98°56'55"W). El Limón is located in northeastern Sierra de Huautla Biosphere Reserve in Morelos State, Mexico (Fig. 1). Topography is variable, with numerous high hills. Elevation at the study site ranges from 1,300 to 1,700 m. The climate is subhumid and warm. Rains occur regularly in June-July and then again in September; October through May tipically is very dry. Annual precipitation is about 800 mm per year (data from the El Limón meteorological station of Comisión Nacional del Agua).
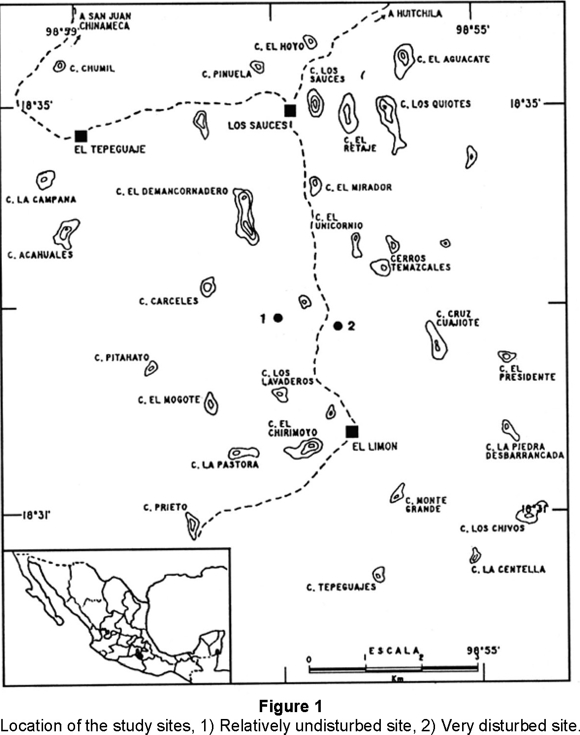
The two study sites were chosen as representative of vegetation disturbance by firewood and lumber extraction, and cattle grazing. They are separated by a distance of 1 km as well as by a land-highway with irregular traffic of vehicles. In El Limón there are 300 inhabitants established approximately more than 100 years ago. It is isolated of the city by 20 km of land-highway. The people use and sale firewood and lumber, and cattle grazing as means of subsistence. Disturbance is concentrated mainly around the town and most of the hills are relatively undisturbed.
Deciduous forest (Rzedowski 1978) covers the site of Ejido El Limón. The relatively undisturbed site (Site 1) is rocky, it includes a narrow canyon, with steep slope, distances between the highest and lowest points are approximately 30 m. In this site, there are species of original vegetation as Licania arborea (cacahuananche; Chrysobalanaceae), Sapindus saponaria (amole; Sapindaceae), Guazuma ulmifolia (cuahulote; Sterculiaceae), Ficus petiolaris (amate; Moraceae), Daphnopsis americana (ahuejote; Thymelaeaceae), Enterolobium cyclocarpum (parota; Leguminosae), Pithecellobium dulce (guamúchil; Leguminosae), Lysiloma divaricata (tepeguaje; Leguminosae), Bursera grandifolia (cuajiote colorado; Burseraceae) and Euphorbia fulva (pegahueso; Euphorbiaceae). Also, there are columnar cacti of Cactaceae family such as Neobuxbaumia mezcalaensis, Pachycereus weberi, Stenocereus beneckei and S. stellatus.
The very disturbed site (Site 2) is located 1 km E of Site 1. Site 2 is a hillside with slight slope; difference between the highest and lowest points is approximately 15 m, and ground also is rocky. In this site, approximately 90% of the vegetation is disturbed and secondary associations include mainly thorny shrubs of Leguminosae family as Acacia bilimeckii (tehuistle), A. cochliacantha (cubata), A. farnesiana (huizache blanco), A. pennatula (quisache), Eysenhardtia polystachya (palo dulce), Mimosa benthamii, M. chaetocarpa (tepescolhuistle), M. polyantha (palo prieto) and Pithecellobium acatlense (huizache de chintete). In both sites, during the dry season herbaceous vegetation is reduced and during wet season a dense stratum of grass grows to 40 or 50 cm in height. Site 2 is subjected to intense cattle grazing during the wet season, while cattle activity is reduced in Site 1. The Sites have the same climate, similar soils and topography that differ only slightly. The main difference between the two sites is the disturbance caused by firewood and lumber extraction, cattle grazing and the type of vegetation associated with each site.
Rodent trapping
Rodents were trapped with 2-dimensional grids in both habitats. Two-night trapping sessions were performed at 40 days intervals between February 1991 and March 1992. Trapping grids in each site were 100 by 100 m with trap stations spaced at 10-m intervals (100 stations/grid). A total of four hundred Sherman traps were used, two at each capture station, providing 8,800 trap nights for analysis. Traps were placed on the ground and were baited with rolled oats in the afternoon prior to each day of checking traps. Each animal captured was weighed, sexed and aged (by size, pelage and sexual conditions) as juvenile, subadult, or adult. Animals were toe-clipped following a numbered sequence (Martoff 1963). After processing, each animal was released at the point of capture.
Statistical analysis
Using this trapping protocol we determined overall community structure of rodents in each site, as well as how this structure change over the course of the study. Rodent species diversity was calculated using the Shannon-Wiener index (Krebs 1989) and compared with a t-test. As this index underestimates rodent diversity (Zar 1999), we calculated the maximum diversity (H'max) and evenness (J). Population size on each site at each sampling period was estimated by direct enumeration of the Minimum Number Known to be Alive method (MNKA; Krebs 1966). Differences between population densities, residence and movements of the two sites were analyzed by two-factor (sites and time) analysis without replication. Differences in sex ratio were examinated by X2 test (Zar 1999). Movements were measured according the method of activity center by Hayne (1950) modified by Vázquez Bárcena (1980), which considers average deviations from the points of capture, as the radius of a circle, and surface of a circle as the area of activity. Annual wet-season vegetation was compared between sites. Trees, shrubs, cactus and herbs were evaluated qualitatively. Vegetative cover was determined visually. We categorized microhabitat or each trap site and used capture locations of each species to infer microhabitat use. We compared microhabitat use of different rodent species by two-factor (microhabitat and seasons) analysis without replication. Data analyses were processed with JMP statistical software (SAS Institute 1998) and Statistical Ecology (Ludwig & Reynolds 1988). All analyses were considered statistically significant when P<0.05.
Results
Population size
In Site 1 we captured 319 individuals of six species of rodents. The most abundant species were P. levipes (113 individuals, 35.4%), B. musculus (86 individuals, 27.0%), P. melanophrys (59 individuals, 18.5%) and L. irroratus (55 individuals, 17.1%). Rare species were Reithrodontomys megalotis (5 individuals, 1.6%) and Hodomys alleni (1 individual, 0.3%). In Site 2 we captured 90 individuals of six species. The most abundant species were L. irroratus (38 individuals, 42.2%), P. melanophrys (26 individuals, 28.9%) and B. musculus (15 individuals, 16.7%). Rare species were P. levipes (6 individuals, 6.7%), R. megalotis (4 individuals, 4.4%) and Sigmodon hispidus (1 individual, 1.1%).
Cumulative number of species
In Site 1, six species were captured after nine months, while in Site 2 this same number of rodent species was obtained by the second month. Differences in cumulative rate of species were due to capture of H. alleni in Site 1, because other species were recorded at second month in both sites.
Diversity
In Site 1 the number of captured individuals was greater than Site 2 (319 versus 90). However, species diversity in Site 1 (H'= 0.616, H'max= 0.778) and in Site 2 (H'= 0.603, H'max= 0.778) was similar (t= 0.372, df= 8, P= 0.64). Also, evenness in both sites was similar (J= 0.79, J= 0.77).
Population density
In Site 1, from February to June 1991 population of L. irroratus fluctuated from six to seven individuals/ha (ind/ha), but since July population increased to the maximum in November (27 ind/ha). After November the population decreased to three ind/ha in March 1992. Density of B. musculus was 15 ind/ha in February 1991. This number declined to 11 ind/ha in April. A subsequent population maximum was reached in July 1991 with 30 ind/ha; the population was decreased to March 1992 when we did not record individuals. Population density of P. levipes fluctuated from 41 ind/ha in March 1991 to a decrease of 16 ind/ha in March 1992, with increments in June (31 ind/ha) and December 1991 (27 ind/ha). Density of P. melanophrys was 18 ind/ha in February; population maximum (23 ind/ha) was reached in June 1991; following that month, density declined to two ind/ha in March 1992 (Fig. 2).
In Site 2, population density of L. irroratus increased since 12 ind/ha in March to a maximum of 24 ind/ha in July. Following this month, population density declined to March 1992 when no further individuals were captured. In general, population dynamics of B. musculus was low (zero to six ind/ha); population maximum was recorded in March and July of 1991 (six ind/ha). Maximum density of P. melanophrys was recorded in March 1991 (12 ind/ha), this number decreased to September when we found no further individuals, but increased to five ind/ha in January 1992, in March no specimen was captured (Fig. 3). Population densities of B. musculus, P. levipes and P. melanophrys were greater in Site 1 than in Site 2 (F= 9.34, df= 1,10, P= 0.001; F= 154.98, df= 1,10, P= 0.0001; F= 40.42, df= 1,10, P= 0.001; respectively), while differences in population density of L. irroratus between both sites were not significant (F= 0.02, df= 1,10, P= 0.89).
Sex ratio
In Site 1, differences in the ratio of males to females were not significant for L. irroratus (1:1.3; X2= 1.2, df= 1, P= 0.72), B. musculus (1:1.1; X2= 0.1, df= 1, P= 0.24), P. levipes (1:0.8; X2= 1.5, df= 1, P= 0.77) and P. melanophrys (1:1.2; X2= 0.7, df= 1, P= 0.59) over the duration of the study. Also, in Site 2, differences in the ratio of males to females were not significant for L. irroratus (1:1.6; X2= 1.8, df= 1, P= 0.82), B. musculus (1:1.3; X2= 0.3, df= 1, P= 0.41) and P. melanophrys (1:0.7; X2= 0.6, df= 1, P= 0.56).
Age structure
In Site 1, subadults and juveniles were captured almost all year, with a maximum for L. irroratus in November 1991, for B. musculus in February and March 1991, for P. levipes in February, March and December 1991, and for P. melanophrys in February, June and December 1991 (Fig. 2). In Site 2, populations of subadults were recorded in March 1991 for L. irroratus; March, July and August 1991 for B. musculus; and in March and June 1991 for P. melanophrys. Juveniles only were captured from November 1991 to January 1992 for P. melanophrys (Fig. 3).
Residence
In Site 1, average of residence, minimum, maximum and number of individuals in parentheses was for L. irroratus 129 days (35-395, n= 19), for B. musculus 100 days (34-236, n= 20), for P. levipes 175 days (41-395, n= 60) and for P. melanophrys 136 days (40-323, n= 25). In Site 2, average of residence was 143 days for L. irroratus (35-310, n= 19), 92 days for B. musculus (32-138, n= 4) and 76 days for P. melanophrys (32-106, n= 9). Residence of individuals was significantly larger in Site 1 than Site 2 for P. melanophrys (F= 327.18, df= 1,15, P= 0.001) and B. musculus (F= 269.17, df= 1,11, P= 0.001), and were not significant for L. irroratus (F= 2.25, df= 1,18, P= 0.85).
Movements
In Site 1, average movements were greater in the wet than in the dry season for L. irroratus (311.5 versus 130.8 m2), B. musculus (1107.7 versus 550.6 m2) and P. levipes (677.3 versus 448.2 m2), only for P. melanophrys average movements were larger in dry than wet season (762.1 versus 567.9 m2). In Site 2, movements were larger in wet than dry season for L. irroratus (909.4 versus 603.0 m2) and P. melanophrys (1,394.0 versus 990.4 m2). For B. musculus we only recorded the movement of one male (1,570.7 m2) and one female (463.2 m2). The movements of individuals of L. irroratus and P. melanophrys in Site 1 were significantly lower than individuals in Site 2 in wet season (F= 10.01, df= 1,5, P= 0.02; F= 24.04, df= 1,6, P= 0.01; respectively) but differences were not significant in dry season (F= 0.98, df= 1,5, P= 0.65; F= 3.29, df= 1,8, P= 0.10; respectively)
Analysis of vegetation
In Site 1, we recorded 66 plant species of 53 genera and 27 families. The most represented families were Compositae (nine species), Leguminosae (eight species) and Burseraceae (eight species). Out of 66 species recorded, cover in parenthesis, 17 are trees (5,800 m2), 26 are shrubs (1,200 m2), one is cactus (550 m2) and 22 are herbs (2,450 m2). In Site 2, we recorded 50 plant species of 42 genera and 20 families. The most represented families were Leguminosae (17 species) and Cactaceae (nine species). Out of 50 species recorded, cover in parenthesis, eight are trees (1,050 m2), 16 are shrubs (3,800 m2), nine are cactus (800 m2) and 17 are herbs (3,500 m2).
Microhabitat
In Site 1, all rodents were captured more often at trapsites with arboreal coverage (60%, n= 43, for L. irroratus; 68%, n= 40, for B. musculus; 67% n= 184, for P. levipes; 59%, n= 72, for P. melanophrys). However, only P. levipes and P. melanophrys showed a statistical difference (F= 134.58, df= 1,4, P= 0.001, n= 305; F= 15.77, df= 1,4, P= 0.04, n= 134; respectively). In Site 2, species had preference by shrub coverage (47%, n= 38, for L. irroratus; 56%, n= 10, for B. musculus; 55%, n= 16, for P. melanophrys) and in less proportion by the arboreal, annual plants and cactus. Only for L. irroratus preferences for shrub coverage were significant (F= 16.35, df= 1,4, P= 0.04, n= 81).
Discussion and conclussions
Our study is the first work that compares rodent communities in a deciduous forest in Mexico. It is important to recognize the limitations of the design and results of this study, since the treatments were unreplicated. Despite these limitations, our results suggests interesting and important trends, such as indicates that habitat fragmentation can be species-specific. Similar studies have been reported in United States and Australia (Laurance 1991, Laurance & Yensen 1991, Diffendorfer et al. 1995a, Nupp & Swihart 1998, Waters & Zabel 1998).
Contrary to our prediction, there were no significant differences in diversity and evenness of rodent species between the two studies sites. However, the number of individuals captured in Site 1 (319) was larger than Site 2 (90) and there was a shift in species composition in abundant and dominant species between sites. Liomys irroratus, B. musculus and P. melanophrys were abundant in both sites, while P. levipes was the most abundant species in Site 1 and was occasionally captured in Site 2. Hodomys alleni was only captured in Site 1, S. hispidus was only recorded in Site 2 and R. megalotis were occasional in both sites. Similar studies had documented disappearance of resident species and changes in dominance with habitat fragmentation (Diffendorfer et al. 1995a, Nupp & Swihart 1998, Kirkland 1990, Holt 1993, Vázquez et al. 2000). Our results support previous findings, which suggested that habitat disturbance does not modify the diversity of rodents (Johnson et al. 1979, Anthony et al. 1981, Huntly & Inouye 1987, Medellín 1992). However in Site 2 there were 33 species of perennial plants, of which 23 were associated with primary vegetation and they are distributed in patches. Its disturbance caused by firewood and lumber extraction, as well as cattle grazing favored reduction of available refuges for rodents. This pattern was reflected in a smaller number of abundant rodent species (three) in Site 2, in comparison with the four dominant species in Site 1 with lesser disturbances. Occasional captures of H. alleni and R. megalotis in Site 1, and of P. levipes, R. megalotis and S. hispidus in Site 2, suggest that they are transient individuals captured in their way toward other site or searching resources.
Livestock could negatively impact rodent populations directly by trampling burrows and compacting the soil or harvesting seed heads and other plants parts while grazing, thus removing food resources that would otherwise be available to rodents. Although livestock might affect rodents indirectly by altering the species composition of the vegetation and the food available for insects (Heske & Campbell 1991), we estimated only percent cover of annual annual plants and grasses in our vegetation anaylisis and, therefore, cannot measure potential impact of livestock grazing on either seed production or plant species composition.
Liomys irroratus
In Site 1 maximum density was recorded at the beginning of dry season, while in Site 2 it was in wet season. Differences in the timing of the population maximum between the sites could be the result of larger coverage (7,500 m2 versus 5,650 m2 of perennial plants) and richness of plant species (66 versus 50 species) in Site 1, resulting in differences in timing on abundance of seed resources. Densities were similar in both sites, perhaps because this species is adapted to arid regions and to open places (Jones & Webster 1976).
Baiomys musculus
In Site 1 maximum density was recorded in the wet season, which was similar to the pattern observed in Site 2. As we expected, density in Site 2 was lower than in Site 1. Although Packard & Montgomery (1978) have recorded B. musculus in places with secondary vegetation, our study is the first to document changes in habitat use depending on the site-specific degree of disturbance. In Site 1 individuals prefer arboreal cover, while in Site 2 they chose shrubs.
Peromyscus levipes and P. melanophrys
In Site 1 maximum density of P. levipes was recorded at the middle of the dry season, declining at the end of this season and increasing in the wet season. As we hypothesized, population density in Site 1 was significantly larger than Site 2. Our data suggests that P. levipes selects undisturbed places, rocky areas (Jones & Webster 1976), with arboreal cover. Their refuges are in rocks, hollows of trees and cactus. For P. melanophrys in Site 1, the maximum population density was recorded in the wet season, while in Site 2 density was low in the dry season and increased in wet season. Density in Site 1 was significantly larger than in Site 2. This may be due to preference for arboreal cover, which was more abundant in Site 1. Density of P. levipes and P. melanophrys is in the range reported in Mexico for P. boylii (Drake 1958, Collett et al. 1975, Álvarez & Arroyo-Cabrales 1990) and P. levipes (Salgado Pérez 1988). Even where densities are similar across habitat types, disturbance may affect aspects of demography and habitat selection (Geier & Best 1980, Diffendorfer et al. 1995b).
Our data therefore are consistent with the hypotesis that fragmentation can be species-specific, as we expect population sizes and demographic parameters (population density, sex ratio, residence) of Baiomys musculus, Peromyscus levipes and P. melanophrys were reduced in Site 2, therefore we think these species were intolerant to disturbance. Peromyscus levipes was the most affected species with respect to number of individuals captured between two sites (113 versus 6). This species exhibited preferences for arboreal and shrub coverage, which suggests that reduction in plant cover decreased the availability of refuges and resources, and affected significantly population number of this species. Continual disturbance of deciduous forest in Morelos as is now occurring, could cause resident species like B. musculus, P. levipes and P. melanophrys to become locally extinct. As we predicted, population densities, sex ratios and residence for L. irroratus were not different between sites. Although we think this species in tolerant to disturbance because its adapted to arid regions, has affinities for edge habitats and disturbed sites and well-developed vagility, more habitat fragmentation could produce population instability and changes in demographic parameters.
Data on population dynamics are critical to development of management strategies for animal and plant species. We must conserve communities and thus whole assemblages of species. Conservation of a species with its associated community must occur in conjuction with the understanding of how the community and the inclusive species function. One without the other will probably not be successful.
Acknowledegments
We thank Julián Morán-Altamirano, Julio César Lara-Manrique, José Antonio Guerrero-Enríquez, Juan Carlos Arroyo-Cruz, Lucas Benítez-Arellano, Leticia Osorio-Navarro and Alma Fernández-Román for their collaboration and enthusiasm during fieldwork. An earlier draft of the manuscript was reviewed by Robert Manson, Celia López González, Michael and Phyllis Kennedy. Two anonymous reviewers provided many constructed comments on the manuscript. César Barona Ríos provide us logistic support. This study was supported by the Secretaría de Educación Publica (FOMES/91 and FOMES/92) awarded to Universidad Autónoma del Estado de Morelos.
Literature cited
Adler, G. H. & R. Levins. 1994. The island syndrome in rodent populations. Quart. Rev. Biol. 69:473-496. [ Links ]
Álvarez, T. & J. Arroyo-Cabrales. 1990. Cálculos de captura, densidad y ámbito hogareño de tres especies de roedores en un área de influencia humana en el sureste de Durango, México. An. Esc. nac. Cien. biol., Méx. 33:185-210. [ Links ]
Anthony, R. G., L. J. Niles & J. D. Spring. 1981. Small-mammal associations in forested and old-field habitats -A quantitative comparisons. Ecology 62:955-963. [ Links ]
Baker, R. H. & J. K. Greer. 1962. Mammals of the Mexican State of Durango. Publ. Misc. Michigan State Univ., Biol. Ser. 2:25:104. [ Links ]
Bowers, M. A. & S. F. Matter. 1997. Landscape ecology of mammals: relationships between density and patch size. J. Mamm. 78:999-1013. [ Links ]
Ceballos, G. 1990. Comparative natural history of small mammals from tropical forest in western Mexico. J. Mamm. 71:263-266. [ Links ]
Chapman, J. L., and M. J. Reiss. 1992. Ecology. Principles and Applications. Cambridge University Press, Great Britain. [ Links ]
Collett, S. F., C. Sánchez Hernández, K. A. Shump, Jr., W. R. Teska & R. H. Baker. 1975. Algunas características poblacionales de pequeños mamíferos en dos hábitats mexicanos. Anales Inst. Biol. Univ. Nac. Autón. México, Ser. Zool. 46:101-124. [ Links ]
Collins, S. L., D. M. Glenn & D. J. Gibson. 1995. Experimental analysis of intermediate disturbance and initial floristic composition: decoupling cause and effect. Ecology 76:486-492. [ Links ]
Connell, J. H. 1980. Diversity and the coevolution of competitors, or the ghost of competition past. Oikos 35:131-138. [ Links ]
Denslow, J. S. 1985. Disturbance-mediated coexistence of species. Pp. 307-323. In: S. T. A. Pickett & P. S. White (eds). The ecology of natural disturbance and patch dynamics. Academic Press, Inc. San Diego, California. [ Links ]
Diffendorfer, J. E., N. A. Slade, M. S. Gaines & R. D. Holt. 1995a. Population dynamics of small mammals in fragmented and continuous old-field habitat. Pp. 175-199. In: W. Z. Lidicker, Jr. (ed). Landscape approaches in mammalian ecology and conservation. University of Minnesota Press, Minneapolis. [ Links ]
Diffendorfer, J. E., M. S. Gaines & R. D. Holt. 1995b. Habitat fragmentation and movements of three small mammals (Sigmodon, Microtus, and Peromyscus). Ecology 76:827-839. [ Links ]
Dowler, R. C. & H. H. Genoways 1978. Liomys irroratus. Mamm. Species 82:1-6. [ Links ]
Drake, D. D. 1958. The brush mouse Peromyscus boylii in southern, Durango. Publ. Mus., Michigan State Univ., Biol. Ser. 1:97-132. [ Links ]
Fa, J. E. & V. Sánchez-Cordero. 1993. Small mammal population responses to fire in a Mexican high-altitude grassland. J. Zoology, London 230:343-347. [ Links ]
Geier, A. R. & L. B. Best. 1980. Habitat selection by small mammals of riparian communities: evaluating effects of habitat alterations. J. Wildl. Manage 44:16-24. [ Links ]
Gilpin, M. 1996. Metapopulation and Wildlife Conservation: Aproaches to modeling spatial structure. Pp. 11-27. In D. R. McCullough (ed). Metapopulation and Wildlife Conservation. Washington, D.C. [ Links ]
Hayne, O. W. 1950. Apparent home range of Microtus in relation to distance between Traps. J. Mamm. 31:26-39. [ Links ]
Herkert, J. R. 1994. The effects of habitat fragmentation on midwestern grassland bird communities. Ecol. Appl. 4:461-471. [ Links ]
Heske, E. J. & M. Campbell. 1991. Effects of on 11-year livestock exclusure on rodent and ant numbers in the Chihuahuan desert, southeastern Arizona. Southwest. Nat. 36:89-93. [ Links ]
Holt, R. D. 1993. Ecology at the mesoscale: The influence of regional processes on local communities. Pp. 77-88. In: R. Ricklefs & D. Schuleter. (eds). Species Diversity in Ecological Communities: Historical and Geographical Perspectives. University of Chicago Press, Chicago. [ Links ]
Huntly, N. & R. S. Inouye. 1987. Small mammal populations and old-field chronosequence: successional patterns and associations with vegetation. J. Mamm. 68:739-745. [ Links ]
Johannesen, E. & R. A. Ims. 1996. Modeling survival rates: habitat fragmentation and destruction in root vole experimental populations. Ecology 77:1196-1209. [ Links ]
Johnson, W. C., R. K. Schreiber & R. L. Burgess. 1979. Diversity of small mammals in a powerline right-of-way and adjacent forest in east Tennessee. Am. Midl. Nat. 101:231-235. [ Links ]
Jones, G. S. & J. D. Webster. 1976. Notes on distribution, habitat and abundance of some mammals of Zacatecas, Mexico. Anales Inst. Biol. Univ. Nac. Autón. México, Ser. Zool. 47:75-84. [ Links ]
Kirkland, G. L., Jr. 1989. Meadow voles (Microtus pennsylvanicus) on forest clearcuts: the role of long-distance dispersal. J. Pennsylvania Acad. Sci. 62:83-85. [ Links ]
––––––––––. 1990. Patterns of initial small mammal community change after clearcutting of temperate North American forests. Oikos 59:313-320. [ Links ]
Krebs, C. J. 1966. Demographic changes in fluctuating populations of Microtus californicus. Ecol. Monog. 36:239-273. [ Links ]
––––––––––. 1989. Ecological methodology. Harper Collins Publishers, New York. [ Links ]
Laurance, W. F. 1991. Ecological correlates of extinction proneness in Australian tropical rain forest mammals. Conserv. Biol. 5:79-89. [ Links ]
Laurance, W. F., and E. Yensen. 1991. Predicting the impacts of edge effects in fragmented habitats. Biol. Conserv. 55:77-92. [ Links ]
Ludwig, J. A., and J. F. Reynolds. 1988. Statistical ecology. A primer on methods and computing. John Wiley & Sons, New York. [ Links ]
Martoff, B. S. 1963. Territoriality in the green frog, Rana clamitans. Ecology 34:165-174. [ Links ]
Martínez Gallardo, R. 1995. Remoción postdispersión de semillas y frutos por mamíferos en diferentes grados de perturbación antropogénica de la selva alta perennifolia en la región de los Tuxtlas, Veracruz. Ph.D. dissertation. Facultad de Ciencias, Universidad Nacional Autónoma de México. [ Links ]
Matson, P. A., W. J. Parton, A. G. Power & M. J. Swift. 1997. Agricultural intensification and ecosystem properties. Science 277:509-515. [ Links ]
Medellín, R. A. 1992. Community ecology and conservation of mammals in a mayan tropical rainforest and abandoned agricultural fields. Ph.D. dissertation, University of Florida. [ Links ]
Moilanen, A. & I. Hanski. 1995. Habitat destruction and coexistence of competitors in a spatially realistic metapopulation model. J. Anim. Ecol. 64:141-144. [ Links ]
Noble, I. R. & R. Dirzo. 1997. Forests as human-dominated ecosystems. Science 277:522-525. [ Links ]
Nupp, T. E. & R. K. Swihart. 1998. Effects of forest fragmentation on population attributes of white-footed mice and eastern chipmunks. J. Mamm. 79:1234-1243. [ Links ]
Packard, R. L. & J. B. Montgomery, Jr. 1978. Baiomys musculus. Mamm. Species 102:1-3. [ Links ]
Pickett, S. T. A. & P. S. White. 1985. Patch dynamics: A synthesis. In: S. T. A. Pickett & P. S. White (eds). The ecology on natural disturbance and patch dynamics. Academic Press, Inc. San Diego, California. pp. 371-384. [ Links ]
Putman, R. J. 1994. Community Ecology. Chapman & Hall, Great Britain. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa, México, D.F. [ Links ]
Salgado Pérez, A. 1988. Contribución al conocimiento de la densidad poblacional y actividad reproductiva de Peromyscus boylii levipes (Rodentia: Cricetidae) en Conejos, Estado de Hidalgo. B.S. Thesis. Escuela Nacional de Estudios Profesionales Iztacala, Universidad Nacional Autónoma de México. [ Links ]
SAS Institute, Inc. 1998. JMP. Statistics and Graphics Guide. Cary, NC. [ Links ]
Stiling, P. 1999. Ecology. Theories and applications. Prentice-Hall, Inc. New Jersey. [ Links ]
Terborgh, J. 1992. Maintenance of diversity in tropical forest. Biotropica 24:283-292. [ Links ]
Terborgh, J. & B. Winter. 1980. Some causes of extinction. Pp. 119-133. In: M. E. Soulé & S. A. Wilcox (eds). Conservation biology: An evolutionary-ecological perspective. Sinauer Press, Sunderland, Massachusetts. [ Links ]
Tilman, D. 1982. Resource Competition and Community Structure. Princeton Univ. Press, Princeton, New Jersey. [ Links ]
Vázquez Bárcena, L. A. 1980. Contribución al conocimiento del área de actividad, densidad de población y actividad reproductiva de Microtus mexicanus mexicanus (Rodentia: Microtinae) en la Sierra del Ajusco, México. B.S. Thesis. Facultad de Ciencias, Universidad Nacional Autónoma de México. [ Links ]
Vázquez, L. B., R. A. Medellín & G. N. Cameron. 2000. Population and community ecology of small rodents in montane forest of western Mexico. J. Mamm. 81:77-85. [ Links ]
Waters, J. R. & C. J. Zabel. 1998. Abundances of small mammals in fir forest in northeastern California. J. Mamm. 79:1244-1253. [ Links ]
Waiters, L., P. Casale & A. A. Dhondt. 1994. Space use and dispersal of red squirrels in fragmented habitats. Oikos 69:140-146. [ Links ]
Wiens, J. A. 1985. Vertebrate responses to environmental patchiness in arid and semiarid ecosystems. Pp. 169-193. In: S. T. A. Pickett & P. S. White (eds). The ecology on natural disturbance and patch dynamics. Academic Press, Inc. San Diego, California. [ Links ]
Williams, S. E. & H. Marsh. 1998. Changes in small mammal assemblage structure across a rain forest/open forest ecotone. J. Trop. Ecol. 14:187-198. [ Links ]
Yahner, R. H. 1991. Dynamics of a small mammal community in a fragmented forest. Am. Midl. Nat. 127:381-391. [ Links ]
Zar, J. H. 1996. Biostatistical analysis. Prentice-Hall. Inc. Englewood Cilffs, New Jersey. [ Links ]

