










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAbanico veterinario
versión On-line ISSN 2448-6132versión impresa ISSN 2007-428X
Abanico vet vol.16 Tepic ene./dic. 2025 Epub 04-Jul-2025
https://doi.org/10.21929/abavet2025.1
Case study
Factors affecting the density of Vibrio parahaemolyticus in the hepatopancreas of shrimp Penaeus vannamei in production units in Northwest Mexico
1
 http://orcid.org/0009-0002-6044-2890
http://orcid.org/0009-0002-6044-2890
1
2
http://orcid.org/0000-0002-9460-4192
3
http://orcid.org/0000-0003-3216-2007
1Universidad Autónoma de Nayarit. Unidad académica de Agricultura. Programa de Doctorado en Ciencias del Posgrado en Ciencias Biológico Agropecuarias. Xalisco, Nayarit, México.
2 Universidad Autónoma de Nayarit. Escuela Nacional de Ingeniería Pesquera. San Blas, Nayarit, México.
3Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, Unidad Mérida. Mérida, Yucatán, México.
Vibrio parahaemolyticus acts as an opportunistic and primary pathogen in shrimp production units, causing Acute Hepatopancreatic Necrosis Disease (AHPND) and the much more virulent postlarvae glass disease (HLVD) than the AHPND strain. However, the variables that determine the density of this pathogen in production units are not yet clear. For this reason, the aim of this study was to determine the physical, chemical and biological variables that favour its development in the hepatopancreas of Penaeus vannamei shrimp, in the water and sediment of culture ponds. The research was carried out in commercial farms located on the northwest coast of Mexico. At each farm, water, sediment and shrimp samples were collected for bacteriological analysis (CHROMAgar Vibrio plate seeding), water quality (nitrite NO2 -, nitrate NO3 -, ammonium NH4 +, phosphate PO4 3- and alkalinity CaCO3), organic matter and pH (in sediment), as well as measurements of salinity, temperature, dissolved oxygen, pH (in water) and transparency. To analyze the association of physical, chemical and biological variables with the density of V. parahaemolyticus in the shrimp hepatopancreas, a Generalized Linear Model (GLM) was constructed. The model that best explained the data had an explained variance of 34.85%. Bacteriological examination showed that the northern and southern zones showed the highest average density of V. parahaemolyticus in the shrimp hepatopancreas. The model revealed a significant effect (P < 0.05) on the density of V. parahaemolyticus in culture pond water and sediment, culture cycle, dissolved oxygen, salinity, pH in culture water, NH4 + and alkalinity. The evidence found for the effect of some of the factors on the density of V. parahaemolyticus in the shrimp hepatopancreas suggests constant monitoring to avoid densities detrimental to shrimp health.
Keywords: shrimp farms; environmental variables; generalized linear model; Vibrio parahaemolyticus
El Vibrio parahaemolyticus actúa como patógeno oportunista y primario de las unidades de producción camaronícola, lo que ocasiona la Enfermedad de la Necrosis Hepatopancreática Aguda (AHPND, por sus siglas en inglés) y la Enfermedad de las postlarvas de cristal (HLVD) mucho más virulenta que la cepa AHPND. Sin embargo, las variables que determinan la densidad de este patógeno en las unidades de producción no son todavía claras. Por tal motivo, el objetivo del presente trabajo fue determinar las variables físicas, químicas y biológicas que favorecen su desarrollo en el hepatopáncreas del camarón Penaeus vannamei, en el agua y en el sedimento de los estanques de cultivo. La investigación se realizó en granjas comerciales localizadas en la costa del noroeste de México. En cada granja se recolectaron muestras de agua, sedimento y camarón para análisis bacteriológico (siembra en placa con CHROMAgar Vibrio), calidad de agua (nitritos NO2 -, nitratos NO3 -, amonio NH4 + , fosfatos PO4 3- y alcalinidad CaCO3), materia orgánica y pH (en sedimento), así como mediciones de salinidad, temperatura, oxígeno disuelto, pH (en agua) y transparencia. Para analizar la asociación de las variables físicas, químicas y biológicas con la densidad de V. parahaemolyticus en el hepatopáncreas del camarón, se construyó un Modelo Lineal Generalizado (MLG). El modelo que mejor explicó los datos presentó una devianza explicada del 34.85%. El examen bacteriológico arrojó que la zona norte y la zona sur mostraron la mayor densidad promedio de V. parahaemolyticus en el hepatopáncreas del camarón. El modelo reveló un efecto significativo (P<0.05) en la densidad de V. parahaemolyticus en el agua y en el sedimento de los estanques de cultivo, ciclo de cultivo, oxígeno disuelto, salinidad, pH en el agua de cultivo, NH4 + y alcalinidad. La evidencia encontrada del efecto de algunos de los factores sobre la densidad de V. parahaemolyticus en el hepatopáncreas del camarón sugieren un monitoreo constante para evitar densidades perjudiciales para la salud del camarón.
Palabras clave: granjas de camarón; variables ambientales; modelo lineal generalizado; Vibrio parahaemolyticus
INTRODUCTION
The bacterium Vibrio parahaemolyticus inhabits coastal marine environments and estuarine areas (Cariani et al., 2012); it is found free in suspension or associated with suspended particles, such as: sediment (Heitmann et al., 2005), plankton (Turner et al., 2013), fish (Aliaga et al., 2010), bivalve mollusks (Rodríguez-Camacho et al., 2014), and crustaceans. In the latter, it usually lodges in the exoskeleton of shrimp because chitin is used by the bacteria as a nutrient (Dulanto, 2013).
The main factors influencing the occurrence and distribution of V. parahaemolyticus in seawater are temperature, salinity, nutrient availability, and fluctuations in plankton concentration (Hernández-Ayón et al., 2003; Tantillo et al., 2004). Therefore, an imbalance in physical, chemical, and biological factors can cause the proliferation of V. parahaemolyticus, which is an opportunistic, facultative anaerobic, Gram-negative pathogenic bacterium responsible for high mortality rates (Cañigral-Cárcel, 2011). One of the diseases caused by V. parahaemolyticus is acute hepatopancreatic necrosis disease (AHPND), which has been reported in Asia and in various shrimp species, such as Penaeus monodon. The first outbreaks of this species were reported in 2009 in southern China, causing massive mortalities of up to 100% in crops (Kumar et al., 2021). Another highly lethal disease caused by Vibrio (HLVD), known as “glass postlarvae disease” (GPD) or “translucent postlarvae disease” (TPD), first appeared in 2019 and the causative agent has been identified as virulent strains of V. parahaemolyticus (Vp HLVD) (Zou et al., 2020; Yang et al., 2023). Infection with Vp HLVD severely damages the hepatopancreatic tubule and midgut epithelium of infected shrimp, causing acute and massive mortality in shrimp postlarvae. Although both HLVD and AHPND are caused by vibrios and share similar clinical signs, Vp HLVD is much more virulent than Vp AHPND (Yang et al., 2023).
In Mexico, there is no evidence of HLVD infection, but it is known that the first outbreaks of AHPND occurred in 2013, affecting Penaeus vannamei production in the northeast region, mainly in the states of Sonora, Sinaloa, and Nayarit (Galaviz-Silva et al., 2021), with estimated losses of up to 65 % in approximately 1,500 farms covering an area of 92 962 ha (Nunan et al., 2014). Additionally, it is important to note that the generation time of Vibrio is between 10 and 12 minutes, benefiting from the availability of nutrients in its environment and warm temperatures (> 35 °C), low salinities (< 8 UPS), and alkaline pH (> 7.5) (Zamora-Pantoja et al., 2005). Therefore, some authors mention that the above-mentioned factors would be the main causes of the increase in the occurrence and distribution of this pathogen in seawater (Hernández-Ayón et al., 2003; Tantillo et al., 2004). According to Lai et al. (2015), this disease occurs when shrimp are under stress or when their immune system is depressed, causing significant economic losses to the shrimp industry. It should be noted that the strains of V. parahaemolyticus that cause AHPND in Mexico show genetic differences from Asian strains and have developed resistance to commonly used antibiotics such as oxytetracycline and sulfacloropyridazine-trimethoprim (Galaviz-Silva et al., 2021). These findings highlight the importance of monitoring and managing Vibrio populations in shrimp aquaculture.
Fourteen years after the emergence of AHPND worldwide, this disease continues to affect the shrimp farming industry. Despite this, there are no reports of in situ research confirming the conditions that enhance the development of this microorganism, nor using a Generalized Linear Model (GLM). Nevertheless, predictions have been made and correlations identified for the abundance of Vibrio species using the parametric linear regression model (LRM) and the negative binomial model (NBM), as well as the generalized additive semiparametric model (GAM) (Dequito et al., 2022). The only studies that have analyzed the effect of AHNPD on commercial shrimp farms are those by Estrada-Pérez et al. (2019) and Estrada Pérez et al. (2020) using multiple regression models with a stock model and a bioeconomic model, respectively. In the present study, we evaluated the different physical, chemical, and biological variables that could be contributing to the increase in the densities of this pathogen in the hepatopancreas of Penaeus vannamei shrimp, farmed in production units located in northwestern Mexico. In addition, the association of physical, chemical, and biological variables of coastal water bodies on the coastline of northwestern Mexico and of the water in the cultivation ponds of shrimp farms with the increase in the density of V. parahaemolyticus in the hepatopancreas of Penaeus vannamei shrimp, using Generalized Linear Models (Nelder & Wedderburn, 1972) as an analysis tool. The purpose of the above is to generate scientific bases about the behavior of V. parahaemolyticus in shrimp culture ponds, which will contribute to decision making by the producer for the control and management of the pathogen in his culture facilities.
MATERIAL AND METHODS
Sample collection and measurement of environmental variables
A database of coastal water bodies was obtained, as well as from shrimp farms located in northwestern Mexico, specifically along the coastline of the state of Sinaloa (Figure 1) during 2017, in which ten samples were taken from 26 farms in the first cycle (FebruaryJune) and 18 in the second (July-November). The sampling areas were: northern zone: Ahome and Guasave; north-central zone: Angostura and Navolato; central zone: Eldorado and Cospita; south-central zone: Elota; and southern zone: Mazatlán. During the first 30 days of cultivation, sampling was carried out weekly, as this was considered the critical stage (Hong et al., 2016). Subsequently, sampling was carried out every two weeks.
Samples of organisms, water, and sediment from each aquaculture production unit were obtained from a randomly selected pond at the inlet and outlet gates. For the organism samples, 15 shrimp with feeding disorders and physiological alterations were selected (Soto-Rodríguez et al., 2010). For fresh and bacteriological analysis, the shrimp were kept alive and placed in ice chests with pond water to be transported to the laboratory of the State Committee for Aquaculture Health of Sinaloa (CESASIN) where they were processed. Water samples were taken between 20 cm below the surface and 20 cm above the bottom. For bacteriological analysis, the samples were transferred to the aforementioned laboratory, using CHROMagar Vibrio plates for bacterial culture, and the results were subsequently quantified and expressed in units of volume (CFU/ml) or mass (CFU/g) (Soto-Rodríguez et al., 2015). Sediment samples for organic matter and pH analysis of the pond culture were collected with a sediment probe. Organic matter (OM) was evaluated in accordance with NOM-021-SEMARNAT-2000 using the Walkley & Black (1934) method through the oxidation of organic carbon (OC). In each sampling, data on salinity (S), temperature (T, °C), oxygen (OD, mg L-1), and pH were recorded using a VITAL SINE SR6 handheld refractometer, an YSI-55-12 oximeter, and an YSI pH 10A potentiometer, respectively, and transparency (Tp, cm) using a Secchi disk.
Concentrations of nitrites (NO2 -), nitrates (NO3 -), ammonium (NH4 +), and phosphates (PO4 3-) were determined using the procedures described in the LYSA water analysis manual, based on the principle of Strickland & Parsons (1972). Alkalinity (Alc) was measured by the colorimetric method described by Hanna, using a HI755 manual colorimeter.
Generalized linear model and statistical analysis
To perform an exploratory analysis of the data, measures of central tendency were obtained for the physicochemical variables, nutrients, organic matter, sediment pH, and bacteriology (Figure 2). In order to determine the association between the different variables with respect to the density of V. parahaemolyticus (CFU/g) in the hepatopancreas of shrimp, Generalized Linear Models (GLM) (Nelder & Wedderburn, 1972) were adjusted using the base package of the R Core Team 2021 programming language.
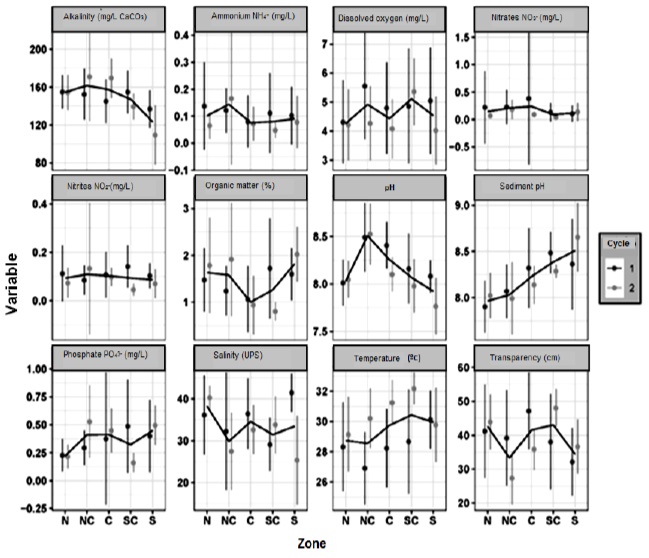
Figure 2 Average ± standard deviation by zone and cultivation cycle of physicochemical and biological variables in shrimp farms in northwestern Mexico. N = North, NC = North Central, C = Central, SC = South Central, and S = South. The black line marks the average between cultivation cycles
To construct the MLGs, T, DO, S, pH, Tp, NO2 -, NO3 -, NH4 +, PO4 3-, Alc, OM, pHs, month of cultivation, and density of V. parahaemolyticus (CFU/ml) in coastal water bodies and cultivation ponds were established as explanatory variables, as shown in Table 1. Since all explanatory variables were quantitative, a quadratic effect (second-degree polynomial) was added to the model in order to improve the fit of the variables, attempting to better explain the trend of the observed data, which was previously evaluated through exploratory analysis (Sokal & Rohlf, 1986). Since the CFU/g values in the hepatopancreas can only be equal to or greater than zero, a Gamma distribution was assumed in the model error, using the value of x + 1 as the response variable to eliminate the zeros. To assume a Gamma distribution, an inverse link function (ƞ = 1/µ) was used. The model selection for the most relevant explanatory variables was performed using the stepwise process with the Akaike criterion as the selection factor (AIC), choosing as the best model the one with the lowest AIC value (Anderson & Burnham, 2002).
Table 1 Explanatory variables used for the Generalized Linear Model
| Cycle I | Cycle II | Cycle I-II | |
|---|---|---|---|
| Variables | February-June | July-December | February-December |
| Mean ± SD | |||
| Physical and chemical properties of water | |||
| Temperature (ºC) | 28.3 ± 2.8 | 30.2 ± 2.2 | 29.0 ± 2.8 |
| Oxigen (mg/L) | 4.8 ± 1.7 | 4.2 ± 1.2 | 4.6 ± 1.6 |
| Salinity (UPS) | 35.6 ± 10.0 | 31.8 ± 9.2 | 34.2 ± 9.9 |
| pH | 8.2 ± 0.3 | 8.1 ± 0.4 | 8.2 ± 0.4 |
| Transparency (cm) | 40.9 ± 13.4 | 36.2 ± 9.9 | 34.2 ± 12.5 |
| Water nutrients (mg/L) | |||
| Nitrates (NO3 -) | 0.242 ± 0.74 | 0.118 ± 0.125 | 0.197 ± 0.592 |
| Ammonium (NH4 +) | 0.112 ± 0.139 | 0.098 ± 0.149 | 0.107 ± 0.143 |
| Nitrites (NO2 -) | 0.107 ± 0.091 | 0.094 ± 0.154 | 0.102 ± 0.118 |
| Phosphates PO4 -3) | 0.324 ± 0.371 | 0.406 ± 0.256 | 0.354 ± 0.335 |
| Alkalinity | 149 ± 22 | 154 ± 38 | 151 ± 29 |
| Sediment | |||
| OM (%) | 1.4 ± 0.7 | 1.6 ± 1.0 | 1.5 ± 0.8 |
| pH | 8.17 ± 0.41 | 8.19 ± 0.38 | 8.19 ± 0.40 |
| Hepatopancreas (UFC/g) | |||
| Densidad de V. parahaemolyticus | 46102.9 ± 265988.73 | 13543.8 ± 54802.76 | 33786.1 ± 212785.82 |
| Bacteriology (UFC/ml) (UFC/g) | |||
| Water- bodies | 104.58 ± 781.87 | 102.46 ± 246.12 | 103.80 ± 639.82 |
| Sediment- bodies | 8256.34 ± 52608.30 | 12383 ± 35684.82 | 9765.47 ± 47131.54 |
| Water -ponds | 47.44 ± 136.50 | 87.86 ± 391.54 | 62.22 ± 260.86 |
| Sediment-ponds | 1931.99 ± 5508.59 | 2856.38 ± 7218.00 | 2270.04 ± 6197.39 |
Note: OM, organic matter
RESULTS
Bacteriology
With regard to the presence of V. parahaemolyticus in the hepatopancreas of shrimp, it was observed that the highest density of V. parahaemolyticus was found in both production cycles in the northern part of the state (Figure 3a). As sampling moved southward, a clear trend toward a decrease in the concentration of V. parahaemolyticus was observed. It was also observed that, in the north-central area, there was a higher concentration of bacteria during the month of October and only during the second production cycle (Figure 3b), while in the central and southern areas of the state, the month of September stood out (Figures 3c and d), and in the south-central area, the month of November stood out (Figure 3e).
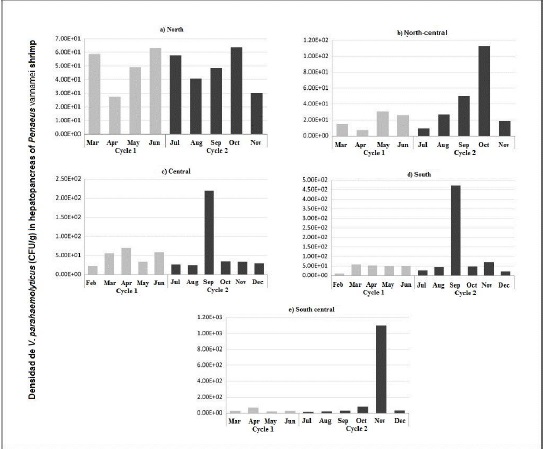
Association of physical, chemical, and biological variables with the density of V. parahaemolyticus in the hepatopancreas of shrimp
The significant effect (P<0.05) of DO, S, pH of the culture water, NO3 -, NH4 +, Alc, OM, pHs, month of culture, and density of V. parahaemolyticus (CFU/ml) in water bodies and culture ponds explained the phenomenon of V. parahaemolyticus density in the hepatopancreas of shrimp with a null deviance of 5895.2, a residual deviance of 3840.2, and an explained deviance of 34.85% (Table 2).
Table 2 Statistical analysis of the physical, chemical, and biological variables that enhance the density of V. parahaemolyticus in the hepatopancreas of Penaeus vannamei shrimp
| Variable | Coefficient Estimated | Error Standard | t Value | P |
|---|---|---|---|---|
| (Intercept) | -5.2964 | 7.7347 | -0.6847 | 0.4940 |
| CFU in water | ||||
| (Water bodies) | 0.0016 | 0.0011 | 1.4304 | 0.1537 |
| CFU in water (Farm) | -1.01E-05 | 1.94E-06 | -5.1911 | 4.05E-07 |
| CFU in Sediment (Farm) | -1.33E-08 | 3.41E-09 | -3.9066 | 0.0001 |
| Cycle: 2 | -1.7144 | 0.6305 | -2.7191 | 0.0069 |
| Month: april | -3.6409 | 3.8923 | -0.9354 | 0.3503 |
| Month: may | -3.3051 | 3.8084 | -0.8678 | 0.3862 |
| Month: june | -4.1755 | 3.8211 | -1.0927 | 0.2754 |
| Month: july | -2.4848 | 3.8558 | -0.6444 | 0.5198 |
| Month: august | -2.8677 | 3.8537 | -0.7441 | 0.4574 |
| Month: september | -4.1767 | 3.8496 | -1.0849 | 0.2788 |
| Month: october | -2.7915 | 3.8384 | -0.7272 | 0.4676 |
| Month: november | -5.8335 | 3.9263 | -1.4857 | 0.1384 |
| Month: december | -7.2182 | 4.2012 | -1.7181 | 0.0868 |
| Dissolved Oxygen (Farm) | -0.4408 | 0.1553 | -2.8375 | 0.0048 |
| Salinity (Farm) | -0.0039 | 0.0019 | -2.0667 | 0.0396 |
| pH in Water (Farm) | -0.1034 | 0.0394 | -2.6204 | 0.0092 |
| Organic matter (Farm) | 0.3915 | 0.2429 | 1.6116 | 0.1081 |
| pH in Sediment (Farm) | 0.0667 | 0.0358 | 1.8622 | 0.0636 |
| NH4 in water (Farm) | 4.4103 | 2.0688 | 2.1317 | 0.0339 |
| NO3 in Water (Farm) | -2.3772 | 1.3827 | -1.7192 | 0.0866 |
| Alkalinity in Water (Farm) | -0.0005 | 0.0001 | -2.8160 | 0.0052 |
Variables in bold P < 0.05
The model indicated that the density of V. parahaemolyticus in the hepatopancreas increases as the concentration of bacteria found in coastal waters increases (Figure 4). A similar trend in the concentration of bacteria in sediments and water showed that intermediate concentrations increased the density of V. parahaemolyticus in the hepatopancreas; however, concentrations higher than intermediate values showed a decrease (Figure 4).
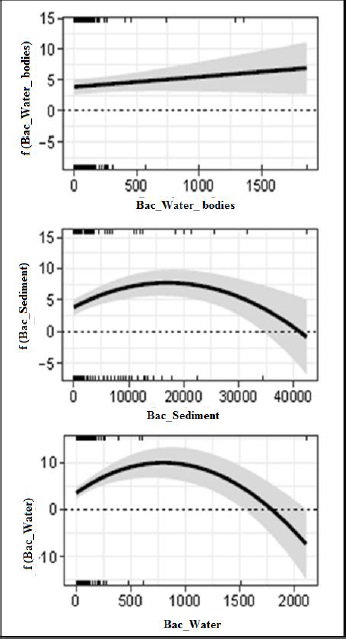
Figure 4 Effect of density (CFU/ml) of V. parahaemolyticus (Y) in coastal water bodies (a: X), in pond sediment (b: X), and in aquaculture pond water (c: X)
Regarding the cultivation cycle, cycle 1 favored V. parahaemolyticus density more than cycle 2 (Figure 5a). In general, higher values of dissolved oxygen, pH, salinity, and nitrates in the water decreased V. parahaemolyticus decreased (Figures 5b, c, d, and h, respectively), contrary to what occurred with organic matter, sediment pH, alkalinity, and ammonium, which, as their values increased, showed higher V. parahaemolyticus density (Figures 5e, f, g, e, and i, respectively).
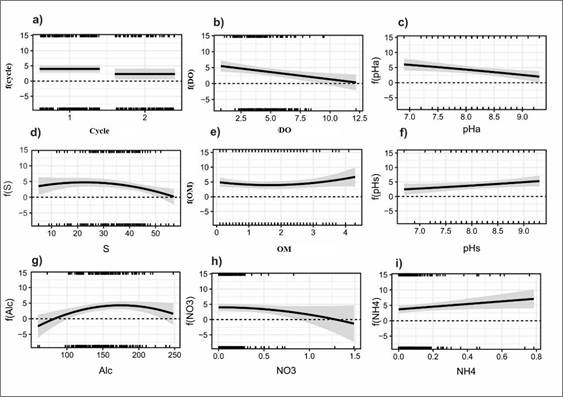
Figure 5 Influence of the culture cycle (a), dissolved oxygen (mg/L) (b), pH in the water (c), salinity (UPS) (d), organic matter (%) (e), pH of the organic matter in the ponds (f), alkalinity (g), NO3 (h), and NH4 (i) on the density of V. parahaemolyticus (CFU/g) in shrimp hepatopancreas f (Cycle, DO, pHa, S, OM, pHs, Alc, NO3, NH4)
DISCUSSION
Environmental variables play a fundamental role in shrimp development due to their physiological condition and because all biochemical processes have direct effects on food and dissolved oxygen consumption rates, ammonium excretion, and growth (Ferreira et al., 2011). In the present study, average temperature, dissolved oxygen, salinity, pH, and transparency were observed for both culture cycles within the optimal range for P. vannamei in culture, values consistent with those reported by other authors, such as temperature from 25 °C to 32 °C (Carbajal-Hernández et al., 2011; Mateka et al., 2015); dissolved oxygen from 4 to 7 mg/L (Alpuche et al., 2005); salinity of 15 to 41 UPS (Pérez-Velázquez et al., 2007; Rivas-Montaño et al., 2018); pH of 6 to 9 (Carbajal-Hernández et al., 2013) and transparency of 17 to 45 cm (García-Sánchez et al., 2018).
The results of this study showed that V. parahaemolyticus in the water supply reached a maximum of 1.35E+04 CFU/ml and, in the southern area, a maximum of 4.94E+03 CFU/ml in pond water. Therefore, it can be presumed that the water supply acts as an inoculum when it enters the cultivation pond. As mentioned by Orellana de Granados & Ayala-Mestanza (2017), these bacteria often find the conditions to grow due to combinations of multiple factors such as high concentrations of organic matter in the bottom, acidic soils, and variation in physicochemical parameters, among others. This proliferation of bacteria can be present in the hepatopancreas and hemolymph of shrimp and cause mortality (Orellana de Granados & Ayala-Mestanza (2017). This is to be expected since the shrimp farming water used in Sinaloa naturally has Vibrio sp. densities greater than 105 CFU/ml (Soto-Rodríguez et al., 2010). This information is significant when considering acute hepatopancreatic necrosis disease (AHPND), since Soto-Rodríguez et al. (2015) mention that among the multiple factors that affect the virulence of V. parahaemolyticus is density, as they found that strain M09-04 caused 93 % mortality at a density of 105 CFU/ml. With regard to the concentration of Vibrio bacteria in the hepatopancreas of shrimp, Orellana de Granados & Ayala-Mestanza (2017) recorded Vibrio sp. densities of 1.10E+03 CFU/ml, with mortalities of up to 10% in the first 30 days of culture. In this study, higher concentrations of V. parahaemolyticus of 3.00E+06 CFU/g were obtained with maximum mortalities of 81%, demonstrating the impact of Vibrio sp. concentrations on mortalities. These results are consistent with Soto-Rodríguez et al. (2015), who pointed out that the virulence of V. parahaemolyticus strains depends on the dose, with an infectious density threshold of 104 CFU/ml; no mortality was observed below that density. Similarly, Soto-Rodríguez et al. (2010) estimate reference levels for possible vibriosis in diseased shrimp at a threshold of 1.40E+05 CFU/g of Vibrio sp. in the hepatopancreas.
The survival of Vibrio spp. in marine environments depends on carbon and energy sources, dissolved oxygen, water pH, salinity, temperature, and starvation (Takemura et al., 2014). Regarding the associations of physicochemical variables, Hung-Sung et al. (2001) and Nelapati et al. (2012) mention that V. parahaemolyticus is a microorganism that has the ability to live and develop in aerobic and anaerobic conditions. However, in the present study, it was observed that at concentrations above 4.6 mg/L and close to 12.5 mg/L of DO, the density of V. parahaemolyticus decreases considerably. Concentrations lower than 4.6 mg/L but higher than 2.5 mg/L DO significantly favor an increase in the density of V. parahaemolyticus in the hepatopancreas of shrimp. These results are similar to those reported by Van Wyk et al. (1999), who suggest that corrective actions should be taken to reduce the density of V. parahaemolyticus when DO levels of 3 mg/L or lower are recorded in the culture water.
With regard to salinity, V. parahaemolyticus can tolerate a wide range of salinities, but its optimal growth conditions vary. In tropical waters, V. parahaemolyticus was found at salinities between 20-35 ppt, with higher densities at 20 ppt (Rivera et al., 1989). Experimental studies showed better adaptation at 10 and 20 UPS compared to 39 and 60 UPS (Sami et al., 2022); this is consistent with the results of this study, where it was observed that at salinities above 34.2 UPS and close to 50 UPS, the density of V. parahaemolyticus decreased, and at salinities of 10-34.2 UPS, the density of the bacteria was higher. Soto-Rodriguez et al. (2019) mention that the toxin can be expressed at different salinities and that salinity management in shrimp farming can be an important factor in controlling the infectivity of V. parahaemolyticus.
Regarding the pH in the water, it is important to note that studies on the impact of pH on the appearance of Vibrio parahaemolyticus in the hepatopancreas of farmed shrimp are scarce, so it is essential to continue with this type of research to clarify the influence of this parameter on the dynamics of Vibrio parahaemolyticus density. The results of this research indicate that V. parahaemolyticus densities (CFU/g) increased when the pH was lower than 8.2, with the highest density of V. parahaemolyticus occurring at values of 7.0. When the pH was greater than 8.2, the density of Vibrio decreased until a pH of 9.0, where the lowest density was found. These results contrast with the research of Mancilla (2005), who indicates that V. parahaemolyticus has the ability to grow in a pH range of 4.8 to 11, with an optimal growth range of 7.5 to 8.8 (FAO/WHO, 2011; Jurquiza, 2014).
Regarding the concentration of organic matter, several authors establish that it can have a direct effect on the health of organisms in culture, since it is capable of modulating bacterial populations and the production of toxic anaerobic metabolites that harm shrimp (Avnimelech & Ritvo, 2003; Nimrat et al., 2008). Merchán-Márquez (2017) establishes that the optimal percentage of organic matter in the soil of white shrimp farming ponds is 3 to 5%, since excessive accumulation of organic matter in the soil favors bacterial density. On the other hand, Díaz-Díaz & Arencibia-Carballo (1999), point out that the recommended organic matter values in pond bottom soil are 1.5% to 3%. In the present study, the average OM concentration was 2%, which is within the optimal range reported by other authors. However, it was observed that at concentrations greater than 2% and less than 0.5%, the density of V. parahaemolyticus present in the hepatopancreas of shrimp increases considerably, which could be due to the presence of organic nitrogen from the balanced feed present in the sediment, which promotes the formation of new bacterial biomass from its metabolization (Paez-Osuna & Alonso-Rodríguez, 2017), or that, when the concentration of organic matter in the bottom soil of the ponds is very high or excessive (>4%), there will be ideal conditions for microbial development, resulting in high concentrations of bacteria, mainly of the genus Vibrio (Díaz-Díaz & Arencibia-Carballo 1999; Nimrat et al., 2008).
Alkalinity refers to base acids such as bicarbonate (HCO3 -), carbonate (CO3 2-) and hydroxide (OH-) responsible for neutralizing water. Ferreira et al. (2011) mention that the level of alkalinity in culture ponds should not exceed 140 mg/L of CaCO3, which coincides with the results found, as they show that when the level of alkalinity is between 150 and 200 mg/L, the density of V. parahaemolyticus in the hepatopancreas of shrimp is higher, while at levels below 140 mg/L and above 250 mg/L, the density of V. parahaemolyticus decreases.
With regard to nitrogen compounds (NH4 +), (NO2 -) and (NO3 -), Tseng & Chen (2004), establish that high concentrations of these compounds affect the immune system of shrimp, leaving them susceptible to bacterial proliferation or causing physiological disorders such as the oxidation of hemocyanin to metahemocyanin. Camargo et al. (2005) mention that the optimal range for (NO3 -) is 0.4 to 1.3 mg/L, while for (NO2 -), Boyd (1995) suggests that an adequate concentration for shrimp development is <0.23 mg/L. However, the results show that at concentrations greater than 0.5 mg/L of NO3 -, the density of V. parahaemolyticus in the hepatopancreas of shrimp decreases considerably, contrary to what occurs with NH4 +, where the concentration of V. parahaemolyticus increases at higher concentrations of NH4 +. This behavior is similar to that reported by Joseph et al. (1982), who established that V. parahaemolyticus does not survive in waters with low nutrient concentrations, while Tantillo et al. (2004) reported that ammonium salts are the main source of nitrogen for V. parahaemolyticus, which is consistent with the present study.
Predicting the abundance of presumed Vibrio species can help prevent the onset of bacterial diseases, as it provides information on when and which environmental factors should be managed. Equito et al. (2022) used the parametric linear regression model (LRM) and the negative binomial model (NBM), as well as the generalized additive semiparametric model (GAM) to identify correlations and predict changes in Vibrio abundance with physicochemical and biological parameters of the water in biofloc ponds with Penaeus vannamei. These authors found that the abundance of presumed Vibrio species was highly correlated with alkalinity, pH, and phytoplankton density. In the present study, using the generalized linear model (GLM), it was shown that as salinity, dissolved oxygen, pH, and nitrates in the water increased, the density of Vibrio decreased (with values outside the optimal range for V. parahaemolyticus), contrary to what was observed with sediments, where higher pH, alkalinity, and ammonium the density of Vibrio increased; concluding that, with an increase in the concentration of bacteria in water and sediment, the model indicated an increase in the concentration of bacteria in the hepatopancreas of shrimp. However, Estrada-Pérez et al. (2019) and Estrada-Pérez et al. (2020), using a stochastic model of AHPND-infected shrimp production, found in their sensitivity analysis that mortality caused by the disease is significantly related to the salinity and transparency of the pond water. These differences in the results of the present study are due to the fact that the disease itself was not considered, but rather the density of V. parahaemolyticus bacteria.
In this research, temperature, transparency, and NO2 - and PO4 3- concentration had no effect on the density of V. parahaemolyticus (CFU/g) in the hepatopancreas of shrimp. According to the model used, it was indirectly detected that 65.15% of factors affected the density of V. parahaemolyticus in the hepatopancreas of shrimp. However, in the present study, it was not possible to measure or consider natural phenomena in the culture environment or management variables, so it is recommended that these be included in future studies to increase predictive capacity. It is concluded that the Generalized Linear model adequately explained the association of various factors with the concentration of V. parahaemolyticus in the hepatopancreas of shrimp, and its results are generally consistent with those published in the literature. Furthermore, the results suggest constant and effective monitoring and management to maintain shrimp crops at recommended values, preventing bacterial loads from increasing to levels that affect shrimp health.
LITERATURA CITADA
Aliaga R, Miranda J, Zevallos J. 2010. Aislamiento e identificación de Vibrio parahaemolyticus O3:K6 en pescados y moluscos bivalvos procedentes de un mercado pesquero de Lima, Perú. Revista Médica Herediana. 21: 135-149. ISSN 1729-214X. http://www.scielo.org.pe/pdf/rmh/v21n3/v21n3ao4.pdf [ Links ]
Anderson DR, Burnham KP. 2002. Avoiding pitfalls when using information-theoretic methods. The Journal of wildlife management. 66(3):912-918. https://doi.org/10.2307/3803155 [ Links ]
Alpuche J, Pereyra A, Agundis C. 2005. Respuestas bioquímicas de camarones marinos a factores ambientales. Revista Electrónica de Veterinaria. 6(5): 1-10. ISSN 1695-7504. https://www.redalyc.org/articulo.oa?id=63617216008 [ Links ]
Avnimelech Y, Ritvo G. 2003. Shrimp and fish pond soils: processes and management. Aquaculture. 220(1-4): 549-567. https://doi.org/10.1016/S00448486(02)00641-5 [ Links ]
Boyd CE. 1995. Bottom soils, sediment and pond aquaculture. Springer Science & Business Media. Pp. 348. ISBN 978-1-4613-5720-9. https://doi.org/10.1007/978-14615-1785-6 [ Links ]
Camargo JA, Alonso A. Salamanca A 2005. Nitrate toxicity to aquatic animals: a review with new data for freshwater invertebrates. Chemosphere. 58(9):1255-1267. https://doi.org/10.1016/j.chemosphere.2004.10.044 [ Links ]
Cañigral-Cárcel I. 2011. Desarrollo de métodos moleculares para la detección y caracterización de bacterias patógenas emergentes del género Vibrio en agua y alimentos. Tesis de doctorado, Escuela Técnica Superior de Ingeniería Agronómica y del Medio Natural, Universidad Politécnica de Valencia. España. Pp. 297. https://doi.org/10.4995/Thesis/10251/11799 [ Links ]
Carbajal-Hernández JJ, Sánchez-Fernández LP, Villa-Vargas LA, CarrascoOchoa JA, Martínez-Trinidad JF. 2013. Water quality assessment in shrimp culture using an analytical hierarchical process. Ecological Indicators. 29: 148-158. https://doi.org/10.1016/j.ecolind.2012.12.017 [ Links ]
Carbajal-Hernández JJ, Sánchez-Fernández LP, Pogrebnyak O. 2011. Assessment and prediction of water quality in shrimp culture using signal processing techniques. Aquacult Int. 19:1083-1104. https://doi.org/10.1007/s10499-011-9426-z [ Links ]
Cariani A, Piano A, Consolandi C, Severgnini M, Castiglioni B, Caredda G, Candela M, Serratore P, De Bellis G, Tinti F. 2012. Detection and characterization of pathogenic vibrios in shellfish by a Ligation Detection Reaction-Universal Array approach. International journal of food microbiology. 153 (3): 474-482. https://doi.org/10.1016/j.ijfoodmicro.2011.11.010 [ Links ]
Dequito AQD, Corre VJ, Abacan EJ. 2022. Generalised additive model improves estimates of Vibrio species abundance in Penaeus vannamei Boone, 1931 biofloc production system. Asian Fisheries Science. 35: 108-116. https://doi.org/10.33997/j.afs.2022.35.2.002 [ Links ]
Díaz-Díaz DG, Arencibia-Carballo. 1999. Carbón orgánico y manejo de suelos en los estanques de la estación camaronera Tunas de Zaza Cuba. Boletín del Centro de Investigaciones Biológicas. 33(1): 39-48. https://produccioncientificaluz.org/index.php/boletin/article/view/190 [ Links ]
Dulanto GJR. 2013. Identificación rápida de especies del género Vibrio asociados con el cultivo de “langostino blanco Litopenaeus vannamei por amplified ribosomal DNA restriction analysis (ARDRA). Tesis de Licenciatura. Lima, Perú: Universidad Nacional Mayor de San Marcos. Pp. 109. https://hdl.handle.net/20.500.12672/3432 [ Links ]
Estrada-Perez N, Ruiz-Velazco JM, Hernández-Llamas A. 2020. Economic risk scenarios for semi-intensive production of Litopenaeus (Penaeus) vannamei shrimp affected by acute hepatopancreatic necrosis disease. Aquaculture Reports. 18, e100442. https://doi.org/10.1016/j.aqrep.2020.100442 [ Links ]
Estrada‐Perez N, Ruiz‐Velazco JMJ, Magallon‐Barajas FJ, Campa‐Cordova AI, Hernández‐Llamas A. 2019. Dynamic stock model for analysing semi‐intensive production of whiteleg shrimp Litopenaeus (Penaeus) vannamei affected by the acute hepatopancreatic necrosis disease: Assessment of disease severity indicators and relationships with pond water quality parameters. Aquaculture. 51(1):242-254. https://doi.org/10.1111/are.14370 [ Links ]
FAO/WHO (Food and Agriculture Organization of the United Nations/World Health Organization). 2011. Risk assessment of Vibrio parahaemolyticus in seafood: Interpretative summary and Technical report. Microbiological Risk Assessment Series No. 16. Rome. Pp. 193. ISBN 978-92-5-106874-8 (FAO). ISBN 978-92-4-154817-5 (WHO).https://iris.who.int/bitstream/handle/10665/44566/9789241548175_eng.pdf?seq uence=1 [ Links ]
Ferreira NC, Bonetti C, Seiffert WQ. 2011. Hydrological and water quality indices as management tools in marine shrimp culture. Aquaculture. 318 (3-4): 425-433. https://doi.org/10.1016/j.aquaculture.2011.05.045 [ Links ]
Galaviz-Silva L, Robles-Valdez A, Sánchez-Díaz R, Ibarra-Gamez JC, Gómez-Gil B, Molina-Garza ZJ. 2021. Cepas de Vibrioparahaemolyticus que causan necrosis hepatopancreática aguda en camarón cultivado de Sonora, México y su resistencia a antibióticos. Hidrobiológica. 31 (2): 111-123. https://doi.org/10.24275/uam/izt/dcbs/hidro/2021v31n2/Galaviz [ Links ]
García-Sánchez S, Juárez-Agis A, Olivier-Salome B, Rivas-González M, ZeferinoTorres J. 2018. Variables fisicoquímicas ambientales que inciden en el cultivo de camarón Litopenaeus vannamei, en Coyuca de Benítez, Guerrero, México. Revista Mexicana de Agroecosistemas. 5(2): 135-155. ISSN: 2007-9559. https://revistaremaeitvo.mx/index.php/remae/article/view/169/151 [ Links ]
Heitmann G, Jofre M, Hormazabal O, Olea N, Vallebuonas S, Valdés H. 2005. Revisión y recomendaciones para el manejo de diarrea por Vibrio parahaemolyticus. Revista Chilena de Infecciones. 22(2): 131-140. http://dx.doi.org/10.4067/S071610182005000200003 [ Links ]
Hernández-Ayón JM, Zirino A, Marinone SG, Canino-Herrera R, Galindo-Bect MS. 2003. Relaciones pH-densidad en agua de mar. Ciencias Marinas. 29 (4): 497-508. https://www.scielo.org.mx/pdf/ciemar/v29n4/v29n4a12.pdf [ Links ]
Hong X, Lu L, Xu D. 2016. Progress in research on acute hepatopancreatic necrosis disease (AHPND). Aquaculture International. 24:577-593. https://doi.org/10.1007/s10499-015-9948-x [ Links ]
Hung-Sung H, Fang-Hsu S, Kun-Chen C, Yuang-Ting Y, Liang-Chao Y. 2001. Relationships between disease outbreak in cultured tiger shrimp (Penaeus monodon) and the composition of Vibrio communities in pond wáter and shrimp hepatopancreas during cultivation. Aquaculture. 192:101-110. https://doi.org/10.1016/S00448486(00)00458-0 [ Links ]
Joseph SW, Colwell RR, Kaper JB. 1982. Vibrio Parahaemolyticus and Related Halophilic Vibrios. CRC Critical Reviews in Microbiology. 10 (1): 77-124. https://doi.org/10.3109/10408418209113506 [ Links ]
Jurquiza V. 2014. Aislamiento, identificación y caracterización molecular de Vibrio parahaemolyticus en muestras de agua, sedimento y moluscos bivalvos de la costa Atlántica de Buenos Aires. Tesis de maestría, Administración Nacional de Laboratorios e Institutos de Salud, Universidad Nacional de San Martín. Mar del Plata, Argentina. Pp 109. https://aquadocs.org/bitstream/handle/1834/6828/Jurquiza_2014.pdf?sequence=1&isAll owed=y [ Links ]
Kumar V, Roy S, Behera BK, Bossier P, Das BK. 2021. Acute hepatopancreatic necrosis disease (AHPND): virulence, pathogenesis and mitigation strategies in shrimp aquaculture. Toxins. 13(8): e13080524. https://doi.org/10.3390/toxins13080524 [ Links ]
Lai HC, Ng TH, Ando M, Lee CT, Chen IT, Chuang JC, Mavichak R, Chang SH, Yeh MD, Chiang YA, Takeyama H, Hamaguchi HO, Lo CF, Aoki T, Wang HC. 2015. Pathogenesis of acute hepatopancreatic necrosis disease (AHPND) in shrimp. Fish Shellfish Immunol. 47:1006-1014. https://doi.org/10.1016/j.fsi.2015.11.008 [ Links ]
Mancilla E. 2005. Intoxicación por Vibrio parahaemolyticus. Cuadernos médico sociales, Chile. 45: 43-47. ISSN 0716-1336. https://dialnet.unirioja.es/servlet/articulo?codigo=9510683 [ Links ]
Mateka HA, Tamatamah R, Kyewalyanga M. 2015. Study on the Water Quality Parameters in Semi-Intensive Coastal Shrimp Culture System in Mafia Island, Tanzania. Journal of Environment and Earth Science. 5(1):142-150. ISSN 2224-3216 (Paper) ISSN 2225-0948 (Online). https://core.ac.uk/download/pdf/234663985.pdf [ Links ]
Merchán Marquez FR. 2017. Relación de la materia orgánica con la comunidad bacteriana en suelos de piscinas de cultivo de litopenaeus vannamei (Examen complexivo). UTMACH, Unidad Académica de Ciencias Agropecuarias, Machala, Ecuador. Pp. 24. https://repositorio.utmachala.edu.ec/handle/48000/11353 [ Links ]
Nelapati S, Nelapati K, Chinnam BK. 2012. Vibrio parahaemolyticus-An emerging foodborne pathogen-A Review. Vet World. 5(1):48-62. https://doi.org/10.5455/vetworld.2012.48-62 [ Links ]
Nelder J, Wedderburn R. 1972. Generalized Linear Models. Journal of the Royal Statistical Society. 135 (3): 370-384. https://doi.org/10.2307/2344614 [ Links ]
Nimrat S, Suksawat S, Maleeweach P, Vuthiphandchai V. 2008. Effect of different shrimp pond bottom soil treatments on the change of physical characteristics and pathogenic bacteria in pond bottom soil. Aquaculture. 285: e123129. https://doi.org/10.1016/j.aquaculture.2008.08.020 [ Links ]
Nunan L, Lightner D, Pantoja C, Gómez-Jiménez S. 2014. Detection of acute hepatopancreatic necrosis disease (AHPND) in México. Diseases of aquatic organisms. 111:81-86. doi: https://doi.org/10.3354/dao02776 [ Links ]
Orellana de Granados CM, Ayala-Mestanza OA. 2017. Incidencia de parásitos y bacterias del género vibrio en el cultivo de camarón marino desarrollados en cooperativas camaroneras del municipio de Jiquilisco, departamento de Usulután. Revista tecnológica. 10:27-30. ISSN: 2070-0458. http://hdl.handle.net/10972/3020 [ Links ]
Páez-Osuna F, Alonso-Rodríguez R. 2017. Calidad de agua en la camaronicultura: importancia, efectos y manejo. El Colegio de Sinaloa. Comité Estatal de Sanidad Acuícola de Sinaloa, A.C, México. Pp. 146. ISBN: 9786077904175 [ Links ]
Pérez-Velázquez M, González-Félix ML, Jaimes-Bustamante F, Martínez-Córdova LR, Trujillo-Villalba DA. 2007. Investigation of the Effects of Salinity and Dietary Protein Level on Growth and Survival of Pacific White Shrimp, Litopenaeus vannamei. Journal of the World Aquaculture Society. 38(4): 475-485. https://doi.org/10.1111/j.17497345.2007.00121.x [ Links ]
Rivas-Montaño AM, Luis-Villaseñor IE, Piña-Valdez P, Gómez-Gil B, LizárragaPartida ML. 2018. Spatiotemporal distribution of Vibrio parahaemolyticus in relation to environmental parameters in a coastal lagoon on the Pacific coast of northwestern Mexico. Ciencias Marinas. 44(3): 141-153. https://doi.org/10.7773/cm.v44i3.2772 [ Links ]
Rivera S, Lugo T, Hazen TC. 1989. Autecology of Vibrio vulnificus and Vibrio parahaemolyticus in tropical waters. Water Research. 23(7): 923-931. https://doi.org/10.1016/0043-1354(89)90018-3 [ Links ]
Rodríguez-Camacho JC, Méndez-Gómez E, Rivas-Montaño AM, Cortés-Ruiz JA. 2014. Evaluación de la presencia de Vibrio parahaemolyticus en camarón blanco (Penaeus vannamei) silvestre estuarino en el sur de Sinaloa y norte de Nayarit, mediante análisis microbiológico y PCR. Revista Bio Ciencias. 2(4): 282-292. http://dx.doi.org/10.15741/revbio.02.04.06 [ Links ]
Sami Z, Kaouthar M, Nadia C, Hedi BM. 2022. Effect of sunlight and salinity on the survival of pathogenic and non‐pathogenic strains of Vibrio parahaemolyticus in water microcosms. Water Environment Research. 94(2): e10689. https://doi.org/10.1002/wer.10689 [ Links ]
Sokal RR, Rohlf FJ. 1986. Introducción a la Bioestadística. Barcelona: Editorial Reverté. España. Pp. 380. ISBN: 978-84291-18624. [ Links ]
Soto-Rodríguez SA, Gómez-Gil B, Lozano R. 2010. Density of Vibrios in Hemolymph and Hepatopancreas of Diseased Pacific White Shrimp, Litopenaeus vannamei, from Northwestern Mexico. Journal of the World Aquaculture Society. (41): 76-83. https://doi.org/10.1111/j.1749-7345.2009.00335.x [ Links ]
Soto-Rodríguez SA, Gómez-Gil B, Lozano-Olvera R, Betancourt-Lozano M, Morales-Covarrubias MS. 2015. Field and experimental Evidence of Vibrio parahaemolyticus as the Causative Agent of Acute Hepatopancreatic Necrosis Disease of Cultured Shrimp (Litopenaeus vannamei) in Northwestern México. Appl Environ Microbiol. 81(5):1689-1699. https://doi.org/10.1128/AEM.03610-14 [ Links ]
Soto‐Rodriguez SA, Lozano‐Olvera R, Palacios‐Gonzalez DA, Bolan‐Mejia C, Rendon‐Aguilar KG. 2019. Caracterización y condiciones de crecimiento de cepas de Vibrio parahaemolyticus con diferentes grados de virulencia que causan enfermedad de necrosis hepatopancreática aguda en Litopenaeus vannamei. Journal of the World Aquaculture Society. 50(5):1002-1015. https://doi.org/10.1111/jwas.12617 [ Links ]
Strickland JDH, Parsons TR. 1972. A practical handbook of seawater análysis. Second Edition, Bulletin. Fisheries Research Board of Canada. Ottawa, Canada. Pp. 310. http://dx.doi.org/10.25607/OBP-1791 [ Links ]
Takemura AF, Chien DM and Polz MF. 2014. Associations and dynamics of Vibrionaceae in the environment, from the genus to the population level. Frontiers in microbiology. 5:38. https://doi.org/10.3389/fmicb.2014.00038 [ Links ]
Tantillo GM, Fontanarosa M, Di Pinto A, Musti M. 2004. Updated perspectives on emerging vibrios associated with human infections. Letters in applied microbiology. 39 (2): 117-126. https://doi.org/10.1111/j.1472-765X.2004.01568.x [ Links ]
Tseng IT, Chen JC. 2004. The immune response of white shrimp Litopenaeus vannamei and its susceptibility to Vibrio alginolyticus under nitrite stress. Fish and Shellfish Immunology. 17: 325-333. https://doi.org/10.1016/j.fsi.2004.04.010 [ Links ]
Turner JW, Malayil L, Guadagnoli D, Cole D, Lipp EK. 2013. Detection of Vibrio parahaemolyticus, Vibrio vulnificus and Vibrio cholerae with respect to seasonal fluctuations in temperature and plankton abundance. Environmental Microbiology. 16 (4):1019-1028. https://doi.org/10.1111/1462-2920.12246 [ Links ]
Van WYK P, Davis-Hodgkins M, Laramore R, Main KL, Mountain J, Scarpa J. 1999. Farming marine shrimp in recirculating freshwater systems. Ft. Pierce, FL: Harbor Branch Oceanographic Institution. Pp. 128-138. https://www.researchgate.net/publication/242621708_Farming_Marine_Shrimp_in_Reci rculating_Fresh_Water_Systems [ Links ]
Walkley A, Black IA. 1934. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil science. 37(1):29-38. http://dx.doi.org/10.1097/00010694-19340100000003 [ Links ]
Yang F, You Y, Lai Q, Xu L, Li F. 2023. Vibrio parahaemolyticus becomes highly virulent by producing Tc toxins Aquaculture. 576: 739817. https://doi.org/10.1016/j.aquaculture.2023.739817 [ Links ]
Zamora-Pantoja DR, Quiroz-Santiago C, Quiñonez-Ramírez EI. 2005. Un Enemigo Marino Silencioso Vibrio parahaemolyticus. 6(4):1-9. ISSN: 1067-6079. https://www.revista.unam.mx/vol.6/num4/art33/abr_art33.pdf [ Links ]
Zou Y, Xie G, Jia T, Xu T, Wang C, Wan X, Li Y, Luo K, Bian X, Wang X, Kong J, Zhang, Q. 2020. Determination of the infectious agent of translucent post-larva disease (TPD) in Penaeus vannamei. Pathogens. 9: 741. https://doi.org/10.3390/pathogens9090741 [ Links ]
Received: July 17, 2024; Accepted: February 10, 2025
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
