











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Phytophthora capsici es un oomiceto fitopatógeno que puede causar enfermedad en una amplia gama de cultivos de importancia alimentaria en todo el mundo (Kim et al. 2012, Sang-Choon et al. 2019); afectando principalmente a las familias Solanacea y Cucurbitacea, entre las que se incluyen los cultivos de chile, pepino, jitomate, calabaza, melón, entre otros (Hausbeck y Lamour 2004, Hurtado-Gonzáles et al. 2008). Causa la pudrición de frutos, corona, raíz y hojas, y la planta en la enfermedad conocida como tizón tardío. P. capsici es un microorganismo filamentoso, constituido por hifas, que normalmente presenta una morfología colonial estrellada al crecer en medios de cultivo (Erwin y Ribeiro 1996). Su reproducción sexual se realiza mediante oosporas y asexual mediante el desarrollo de esporangios, que contienen células diferenciadas conocidas como zoosporas (Lamour et al. 2012). En el agua, los esporangios liberan las zoosporas, que pueden nadar y dispersarse fácilmente hasta encontrar un tejido vegetal, donde se enquistan para iniciar el proceso de infección (Mohsan et al. 2017). Es un patógeno destructivo que causa pérdidas económicas sustanciales cada año en los cultivos infestados (Gevens et al. 2008, Meitz et al. 2010). La enfermedad que produce es difícil de controlar debido a la rápida reproducción y la diseminación de los propágulos infecciosos (Youyou et al. 2019). Cualquier interrupción en el desarrollo de zoosporas, en su liberación o en su motilidad disminuye la posibilidad de infección (Islam et al. 2011, Kim et al. 2012).
Para el control del tizón tardío tradicionalmente se usan fungicidas de amplio espectro y acción rápida, como metalaxil, mancozeb, propamocarb, cimoxanil, dimetomorf, flumorf, pirimorf y fosetilAl (Nguyen et al. 2012, Qi et al. 2012). Sin embargo, su uso continuo ha generado una serie de problemas ecológicos y ambientales (Ramírez et al. 2020), así como preocupaciones sobre su impacto en la salud humana y el desarrollo de resistencia (Zhao et al. 2010, Sevillano-Serrano et al. 2024). Estos problemas han fomentado la búsqueda de métodos eficientes, sostenibles y seguros para controlar el tizón. El desarrollo y la prueba de nuevos compuestos con actividad inhibitoria y distintos modos de acción es un asunto urgente ante la situación agrícola y ambiental actual.
Durante las últimas dos décadas se han buscado biocidas para patógenos, seguros para el medio ambiente y fácilmente biodegradables (Mohsan et al. 2017). El uso de productos naturales, incluidos los extractos de plantas, ha mostrado eficacia en la inhibición y el control de fitopatógenos; además, los metabolitos secundarios presentes son biodegradables, considerándose como una alternativa ecológica al uso de plaguicidas organosintéticos. Los metabolitos más comunes son terpenoides, fenilpropanoides, quinonas, cumarinas, flavonoides, taninos, otros fenoles y glucósidos cianogénicos (Gulluce et al. 2007, Tian et al. 2011). Se han probado con éxito extractos crudos de col, ajo y alfalfa, con capacidad inhibitoria significativa contra P. capsici. Los extractos de hoja de muérdago han demostrado actividad antimicrobiana (Cervantes-Ortega et al. 2015) y citotóxica contra varios patógenos humanos, como Candida albicans y Acinetobacter lwoffii (Jacobo-Salcedo et al. 2011).
El muérdago es una planta hemiparásita aérea, compuesta por alrededor de 1 600 especies en todo el mundo (Nickrent et al. 2010); doce especies se distribuyen desde México hasta el norte de Argentina (Kuijt 2009). Psittacanthus calyculatus es una especie de muérdago nativa de México (Jacobo-Salcedo et al. 2011), distribuida en 25 estados, tanto en las regiones del centro como en las del sur del país. El muérdago se considera una plaga porque puede secar los árboles que parasita (Azpeitia y Lara 2006). Suele crecer sobre mezquite, huizache, durazno o en pinos (Lara et al. 2021, Quintana-Rodríguez et al. 2018), especialmente en las regiones centrales de México en Michoacán y Guanajuato (Cervantes-Ortega et al. 2015). El muérdago parasita principalmente al mezquite (Prosopis laevigata), árbol endémico del altiplano central (Aguilar-Venegas et al. 2023), lo que convierte a la planta hemiparásita en una amenaza para los mezquites (Quintana-Rodríguez et al. 2018). No obstante, el muérdago ha sido utilizado en la medicina tradicional mexicana (Ontiveros-Rodríguez et al. 2023), como remedio para prevenir infecciones cutáneas y el tratamiento de enfermedades cardiovasculares (Moustapha et al. 2011). El uso de extractos de muérdago ha demostrado afectar a tres hongos patógenos de plantas (Xoca-Orozco et al. 2022). Sin embargo, no existen estudios que evalúen el efecto de los extractos de muérdago sobre oomicetos como P. capsici. Por lo tanto, este estudio tuvo como objetivo evaluar el efecto de los extractos de hojas y flores de muérdago mexicano (P. calyculatus), que parasitaba mezquite (P. laevigata), sobre el desarrollo y la recuperación de P. capsici
Materiales y métodos
Obtención de extractos de hojas y flores de muérdago
Los extractos de hojas (EH-MM) y flores (EF-MM) de muérdago (P. calyculatus) se obtuvieron del parasitismo de esta planta en árboles de mezquite (P. laevigata), localizados en el Área Natural Protegida del Cerro del Palenque de Purísima del Rincón, Guanajuato, México.
Para obtener los extractos se siguió la metodología de extracción de metanol acidificado y acetona (Saura-Calixto et al. 2007), la cual consiste en agregar 50 mL de metanol acidificado (8 mL L−1 HCI, agua 50-50 v/v) por cada gramo de hoja y flor de muérdago pulverizado y mantener durante 60 min a temperatura ambiente. Enseguida, se centrifugó a 3 000 × g durante 5 min y se almacenó a 4 °C el sobrenadante del extracto. Por su parte, el precipitado del extracto se lavó con acetona-agua (70:30 v/v) (50 mL g−1 de muestra) durante 60 min y se centrifugó en las mismas condiciones. Enseguida, se combinaron los sobrenadantes obtenidos de cada lavado (sobrenadante y precipitado). Finalmente, se eliminaron los disolventes en presión reducida durante 120 min a 65 °C en un rotavapor. Ambos extractos se almacenaron a 4 °C en oscuridad hasta su utilización.
Evaluación de extractos de muérdago sobre el crecimiento de P. capsici
Se utilizó la cepa D3 de P. capsici proporcionada por el Dr. Mario M. Gonzáles-Chavira (Pons-Hernández et al. 2020, Sevillano-Serrano et al. 2024) para evaluar el efecto de EH-MM y EF-MM. El oomiceto creció a 28 °C en oscuridad en placas Petri con el medio V8, adicionado con cuatro concentraciones (C1, C2, C3 y C4) de EH-MM y EF-MM. Como control negativo, se utilizaron placas con disolventes de extracción (CDE); además, un segundo control consistió en placas con medio V8 sin la adición de extractos ni disolventes (SED).
La preparación del medio V8 se realizó con 200 mL de jugo V8 Herdez®, 800 mL de agua destilada, 2 g de carbonato de calcio y 15 g de agar por litro, esterilizado en autoclave a 121 °C durante 20 min (Solache-Huacuz et al. 2010). El medio V8 estéril se adicionó con los extractos a cuatro concentraciones distintas. Como control, se preparó un medio al que se adicionaron cantidades similares de disolventes de extracción para descartar el efecto de los residuos de estos en el desarrollo de P. capsici (CDE). Cada placa Petri contenía 15 mL de medio de cultivo V8, adicionado con cuatro concentraciones de EH-MM, EF-MM y disolventes de extracción, constituidos por: C1, 14 mL de medio V8 adicionado con 1 mL (6.7%) de extracto de muérdago; C2, 13 mL medio V8 y 2 mL (13.3%) de extracto; C3, 11.5 mL de medio V8 agregado con 3.5 mL (23.33%) de extracto y C4, 10 mL V8 con 5 mL (33.3%) de extracto. La preparación de las cuatro concentraciones de CDE en medio V8, como control, se realizó con la mezcla de una solución metanol acidificado: agua (1:1) y de acetona: agua (7:3) en la misma proporción, simulando la mezcla de potenciales residuos en el proceso de extracción.
Una vez preparado el medio, se dejó reposar durante 24 h a temperatura ambiente y en condiciones de oscuridad para que se evaporaran los disolventes. Posteriormente se inocularon las placas de medio con discos de agar de 1 cm de diámetro con micelio de P. capsici D3 para medir su cinética de crecimiento hasta los 7 días después de la inoculación (ddi), y así determinar el porcentaje de inhibición; los datos se sometieron a un análisis de varianza y a separación de medias (Tukey, p ≤ 0.05), mediante el software GraphPad Prism v7 y Minitab 20.3. También se evaluó el área pigmentada del medio de cultivo ocasionada por el oomiceto a los 4 y 11 ddi con ayuda del software ImageJ v1.53t.
Evaluación de la morfología colonial de P. capsici expuesto a extractos de muérdago
Se analizó la morfología colonial del patógeno mediante fotografías a los 4 y 11 ddi en medio V8 con EH-MM y EF-MM, y con disolventes de extracción. Además, se evaluaron sus características morfológicas microscópicas, constituidas por hifas y esporangios a los 4 ddi. Para inducir la producción de esporangios, las placas se colocaron en fotoestrés con luz blanca constante a temperatura ambiente durante 4 días (Lamour y Hausbeck 2000). Una vez producidos los esporangios, se tiñeron con azul de metileno al 0.1% para visualizarlos con ayuda de un microscopio óptico LEICA CTR5000 a 40×, lo que permitió describir el efecto de los extractos de muérdago en el desarrollo y la morfología de esporangios, así como identificar modificaciones en las hifas de P. capsici.
Análisis del efecto de EH-MM y EF-MM en germinación y desarrollo de zoosporas
Para conocer el efecto de EH-MM y EF-MM sobre zoosporas de P. capsici, se indujo la liberación de estas células basado en el protocolo descrito por Lamour y Hausbeck (2000), con adición de 10 mL de agua destilada estéril a la placa Petri con P. capsici expuesto durante 4 días en foto-estrés; estas placas se dejaron reposar a 4 °C por 90 min y posteriormente se colocaron a temperatura ambiente por 30 min. Después, se recuperó la suspensión de zoosporas y se cuantificó con un hematocitómetro para ajustar la concentración a 330 zoosporas µL-1.
Enseguida, se prepararon mezclas en microtubos de 1.5 mL que contenían las mismas cuatro concentraciones de EH-MM, EF-MM y CDE en relación con el medio V8, el cual se sustituyó por una suspensión de 1×105 zoosporas en agua, para incubarlas a 28 °C en oscuridad durante 48 h. Se analizó el efecto de los extractos de muérdago sobre las zoosporas con ayuda de un microscopio óptico a 40× y se contó el porcentaje de zoosporas germinadas con el hematocitómetro en dos experimentos independientes. Posteriormente, las zoosporas se lavaron con agua destilada estéril y centrifugadas a 1 500 rpm durante 3 min por dos ocasiones bajo condiciones de esterilidad, se decantó el sobrenadante, se resuspendieron en 10 µL de agua estéril, se inocularon en placas de agar V8 y se incubaron por 7 días a 28 °C en oscuridad, para evaluar su desarrollo y viabilidad después de estar en contacto con EH-MM, EF-MM y CDE.
Análisis morfológico y porcentaje de recuperación de P. capsici después del tratamiento con extractos de muérdago
En condiciones de esterilidad y con ayuda de un sacabocados estéril de 1 cm, se tomó micelio de P. capsici de 7 días de edad, creciendo en cada uno de los tratamientos, y se pasó a nuevas placas Petri con medio V8 sin adición de componentes. Enseguida, se evaluó su crecimiento a los 5 ddi, se calculó el porcentaje de recuperación del oomiceto en cada tratamiento con respecto al control SED, y se realizaron el análisis de varianza y la separación de medias (Tukey, p ≤ 0.05) con el software GraphPad Prism v7 y Minitab 20.3. Al final, se analizó la morfología colonial de recuperación del oomiceto mediante fotografías.
Resultados y discusión
Inhibición del crecimiento de P. capsici causado por extractos de muérdago
La cinética de crecimiento de P. capsici en los extractos mostró que, al aumentar la concentración de EH-MM, EF-MM y CDE, el crecimiento disminuyó con el tiempo de exposición, con diferencias en el crecimiento entre el patógeno expuesto a EH-MM y a EF-MM. Mientras que el control CDE mostró un crecimiento similar al segundo control SED, en el que no se adicionaron extractos ni disolvente (Figura 1).
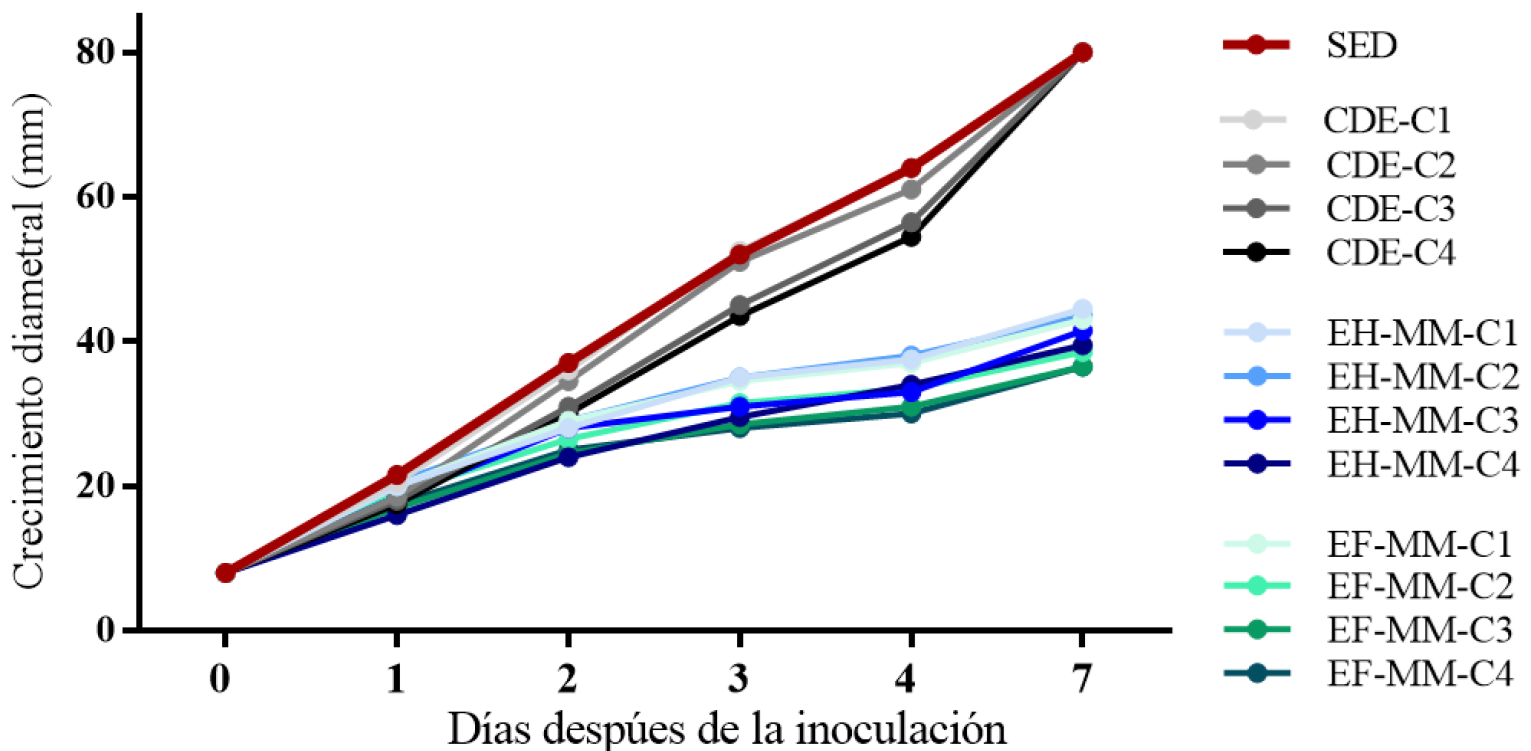
Figura 1 Cinética de crecimiento de P. capsici expuesto a extractos de muérdago. Crecimiento diametral (mm). C1, C2, C3 y C4 = concentración 1, 2, 3 y 4 (6.7 %, 13.3, 23.3 y 33.3%), de extractos de hojas, flores de muérdago, SED y CDE [solución metanol acidificado: agua (1:1) con acetona: agua (7:3)]. EH-MM, extracto de hojas de muérdago. EF-MM, extracto de flores de muérdago. SED, sin extractos y disolventes de extracción. CDE, con disolventes de extracción.
El porcentaje de inhibición del crecimiento de P. capsici a los 7 ddi en medio V8 adicionado con los extractos indicó que EH-MM y EF-MM inhibieron el crecimiento del patógeno en las cuatro concentraciones respecto al control SED (Tabla 1). En el caso del medio V8 CDE, el crecimiento fue similar al de SED, por lo que se consideró que el tiempo de evaporación fue adecuado y que los disolventes no afectaron el crecimiento del patógeno.
Tabla 1 Porcentaje de inhibición del crecimiento de P. capsici expuesto a extractos de muérdago.
| Tratamiento | Crecimiento | |||
|---|---|---|---|---|
| C1 (6.7 %) | C2 (13.3%) | C3 (23.3%) | C4 (33.3%) | |
| CDE | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| EH-MM | 44.38 ± 2.65 | 45.63 ± 2.65± | 48.13 ± 2.65± | 50.63 ± 0.88a |
| EF-MM | 46.25 ± 1.77 | 51.88 ± 0.88a, b,± | 54.38 ± 0.88a, b, c, d, ± | 54.38 ± 0.88a, b, c, d |
CDE, con disolventes de extracción. EH-MM, extracto de hojas de muérdago. EF-MM, extracto de flores de muérdago. Concentración 1, 2, 3, 4 de EH-MM y EF-MM (C1, C2, C3, C4). a, diferencia significativa con respecto a EH-MM en C1 (Turkey, p ≤ 0.05). b, diferencia significativa con respecto a EH-MM en C2 (Turkey, p ≤ 0.05). c, diferencia significativa con respecto a EH-MM en C3 (Turkey, p ≤ 0.05). d, diferencia significativa con respecto a EF-MM en C1 (Turkey, p ≤ 0.05). ±, diferencia significativa (Turkey, p ≤ 0.05) de la misma concentración de extracto obtenido de diferente parte de la planta.
En general, el extracto EF-MM presentó una mayor inhibición del crecimiento del oomiceto que EH-MM, lo que sugiere una composición química distinta entre ambos extractos. También fue significativo el porcentaje de inhibición de P. capsici entre la concentración baja (6.7%) o C1 de EH-MM y EF-MM, y la concentración alta (33.3%) o C4. El menor porcentaje de inhibición se observó en C1 de EH-MM, en comparación con todos los tratamientos evaluados. En cambio, EH-MM en C1, C2 y C3 reveló una inhibición del patógeno menor, de manera significativa, que EF-MM en C2, C3 y C4, lo que demuestra que EF-MM presentó mayor inhibición del crecimiento de P. capsici. En las últimas décadas, los productos naturales provenientes de distintas plantas han tomado gran importancia debido a que son capaces de inhibir diversos fitopatógenos como Aspergillus parasiticuas, A. niger, Passalora fulva, Alternaria solani, Stemphyllium solani, Rhizoctonia solani, Sclerotium rolfsii, Phytophthora parasitica var nicotianae, Fusarium oxysporum, F. graminearum, Phomopsis sp. entre otros (Ruiz et al. 2020, Liang et al. 2024). Lo anterior, representa una alternativa al uso de componentes químicos organosintéticos, ya que estos tienen un impacto ambiental negativo (Jáquez-Matas et al. 2022).
Los extractos de muérdago mexicano contienen una mezcla de compuestos fitoquímicos con actividad antioomicética; en el presente trabajo se observó un porcentaje de inhibición del micelio de P. capsici mayor al 50% con EH-MM y EF-MM. Esto coincide con lo reportado por Xoca-Orozco et al. (2022) para los hongos fitopatógenos C. gloesporoides, Fusarium sp. y Curvularia sp., en donde extractos de muérdago procedentes del mezquite inhibieron el crecimiento a las concentraciones de 6.7%, 13.3%, 23.3% y 33.3% EH-MM y EF-MM. También se observó que a mayor concentración de los extractos, mayor es la capacidad para inhibir el desarrollo de los tres hongos. Sin embargo, podrían realizarse estudios futuros con moléculas purificadas de los extractos de muérdago para evaluar su potencial como fuentes de moléculas de control del desarrollo de fitopatógenos. Anteriormente se había descrito la capacidad de inhibición de extractos de plantas como Parthenium hysterophorus (arrocillo), Allium sativum (ajo), Larrea tridentata (gobernadora), Syzygium aromaticum (clavo), Melia azedarach (melia), Rosmarinus officinalis (romero), Zingiber officinale (jengibre) y Solanum torvum (sosa) sobre el crecimiento de P. capsici (Mohsan et al. 2017). Sin embargo, este trabajo es el primer reporte que documenta la actividad inhibitoria de los extractos de muérdago (P. calyculatus) sobre P. capsici.
Estos resultados indican que el extracto de flores inhibe el crecimiento de P. capsici comparado con el extracto de hojas, contrario con lo reportado para hongos fitopatógenos en donde los extractos de flores y hojas de muérdago inhibieron el crecimiento de los hongos fitopatógenos Fusarium sp., C. gloesporoides y Curvularia sp., sin presentar diferencia significativa (Xoca-Orozco et al. 2022). Por ello, se considera que las flores de muérdago contienen más moléculas antioomicéticas que contribuyen a la inhibición de P. capsici. En el estudio de Moustapha et al. (2011) se analizó el perfil fitoquímico de la fracción soluble en metanol de un extracto acuoso de una muestra de muérdago (P. calyculatus) procedente de la planta hospedera Prosopsis laevigata (mesquite liso), identificándose el ácido gálico, dos flavonol-3-biósidos y el aminoácido no proteico N-metil-trans-4-hidroxi-L-prolina. El perfil fitoquímico cualitativo y cuantitativo de extractos de hojas y flores de muérdago utilizados en esta investigación se realizó en estudios anteriores reportados por Xoca-Orozco et al. (2022), los cuales mostraron que flavonoides como las antocianidinas se encontraban presentes en mayor concentración en el extracto de flores de muérdago. Diversos compuestos fenólicos, incluidos los flavonoides, han mostrado inhibir el crecimiento de fitopatógenos como Asperguillus parasiticus (Pizzolitto et al. 2015). A partir del extracto de metanol de la planta Prunella vulgaris, se aislaron los ácidos poliacetilénicos octadeca 9,11,13 triinoico y ácido trans octadec 13 eno 9,11 diinoico, los que inhibieron el crecimiento de P. capsici (Yoon et al. 2010). Además, el extracto de metanol de los rizomas de Anemarrhena asphodeloides exhibió una fuerte actividad contra P. capsici, cuya principal sustancia oomicida fue el polifenol denominado nyasol: (Z) 1,3 bis(4 hidroxifenil) 1,4 pentadieno (Park et al. 2003), es decir, compuestos fenólicos presentes en los extractos de muérdago pueden inhibir el crecimiento de P. capsici. Por lo tanto, se sugiere que ciertos compuestos fenólicos, flavonoides y demás metabolitos secundarios presentes en EH-MM y EF-MM podrían estar contribuyendo, en mayor o menor medida, a la inhibición de P. capsici.
Los flavonoides se han descrito como agentes biocidas efectivos contra una amplia variedad de organismos patógenos (Yang et al. 2019, Mohotti et al. 2020), ya que inhiben su crecimiento mediante varios mecanismos subyacentes, que incluyen la alteración de la membrana plasmática (Pinto et al. 2009, Walker y White 2005), inducción de apoptosis (Perez et al. 2003), disfunción mitocondrial, elevación de especies reactivas de oxígeno (ROS) (Peralta et al. 2015), inhibición de la formación de la pared celular, inhibición del transporte iónico y división celular (Kang et al. 2010); así como en la síntesis de ADN, ARN (Lagrouh et al. 2017) y proteínas (Abat et al. 2017). Del mismo modo, los flavonoles (miricetina, kaempferol, fisetina, quercetina, 3-hidroxi flavona y 3,7-dihidroxiflavona), una flavona (luteolina), una flavanona (naringenina) e isoflavonas (genisteína, biocanina A) inhiben el hongo filamentoso Cochliobolus lunatus mediante la inhibición de la síntesis de ácidos nucleicos (Cassetta et al. 2017). Por lo tanto, los extractos de muérdago, que contienen diversos flavonoides y polifenoles, podrían estar afectando vías metabólicas importantes en P. capsici, lo que contribuye a la inhibición de su crecimiento.
Cuando se cultivó P. capsici en las cuatro concentraciones de extractos de muérdago, se encontró que el oomiceto pigmenta el medio de cultivo al oscurecerlo sobre el sitio de crecimiento, por lo cual se midió el área pigmentada a los 4 y 11 ddi (Tabla 2). A los 4 ddi, en concentraciones bajas (C1) de ambos extractos, la pigmentación del medio de cultivo fue menor que en C2, ya que ésta mostró la mayor área pigmentada. Sin embargo, en concentraciones mayores (C3 y C4) la pigmentación del medio de cultivo tiende a disminuir proporcionalmente. Por lo cual, ambos extractos mostraron una capacidad inductora de pigmentación similar en cada concentración evaluada.
Tabla 2 Área pigmentada del medio de cultivo (mm2).
| Tratamiento | Área pigmentada | |||
|---|---|---|---|---|
| C1 (6.7%) | C2 (13.3%) | C3 (23.3%) | C4 (33.3%) | |
| EH-MM-4 | 4.07 | 14.55 | 12.76 | 12.5 |
| EH-MM-11 | 51.84 | 19.2 | 19.42 | 28.6 |
| EF-MM-4 | 4.64 | 16.29 | 12.79 | 11.84 |
| EF-MM-11 | 33.18 | 28 | 37.78 | 53.32 |
EH-MM-4, extracto de hojas de muérdago 4 días después de la inoculación (ddi). EH-MM-11, extracto de hojas de muérdago 11 ddi. EF-MM-4, extracto de flores de muérdago cuatro ddi. EF-MM-11, extracto de flores de muérdago 11 ddi. Concentraciones 1, 2, 3 y 4 de EH-MM y EF-MM (C1, C2, C3 y C4).
La pigmentación del medio de cultivo continuó aumentando con el tiempo de exposición del patógeno a los extractos de muérdago, ya que a los 11 ddi se observó un aumento del área pigmentada en cada condición evaluada con respecto a los 4 ddi. En esta segunda etapa de análisis, EF-MM a concentraciones altas provocó la mayor pigmentación del medio de cultivo en comparación con EH-MM. En ambos extractos se encontró que concentraciones bajas (C1) resultan en un área pigmentada similar a la de C4. Sin embargo, disminuyó en C2 y tiende a aumentar progresivamente en C3 y C4. Es decir, la concentración y el tiempo de exposición de P. capsici a EH-MM y EF-MM generaron cambios significativos en el área pigmentada del medio de cultivo.
Los organismos filamentosos se han estudiado por su actividad metabólica en la biosíntesis de metabolitos para la producción extracelular de pigmentos (Carvalho et al. 2003). Estos resultados indican que P. capsici pigmenta el medio de cultivo en respuesta a los extractos de muérdago. En cambio, cuando el oomiceto se pasó a medio de cultivo sin extractos de muérdago dejó de producir los pigmentos, posiblemente debido a una característica metabólica de los patógenos en respuesta al estrés causado por los componentes presentes en los extractos, similar a lo que sucede con algunas especies de Monascus sp., que producen pigmentos hidrosolubles que se difunden por el medio de cultivo con tonalidades rojas a amarillas (Blanc et al. 2001). Otros microorganismos, como los del género Paecilomyces sp., también producen pigmentos en tonalidades rojas, amarillas y violetas debido a los componentes del medio de cultivo (Cho et al. 2002).
Se ha reportado la producción de pigmentos en pruebas de antagonismo contra el oomiceto P. infestans, cuando este es inhibido por los microorganismos Trichoderma sp. y Pseudomonas sp. (Harris-Valle et al. 2020). P. capsici podría presentar un efecto similar al someterse al estrés inducido por las moléculas antagónicas de los extractos de hojas y flores de muérdago. Sin embargo, es necesario profundizar en este fenómeno para determinar si los pigmentos producidos por P. capsici en el medio de cultivo constituyen una respuesta fisiológica inducida por los extractos de muérdago.
Morfología colonial de P. capsici expuesto a extractos de muérdago
La morfología colonial de P. capsici (Figura 2) a los 4 ddi mostró asimetría del crecimiento en el borde del micelio y de tipo ralo, es decir, poca densidad de micelio aéreo comparada con el control SED (Figura 2A). Por su parte, el control CDE mostró una disminución en la definición de picos en el micelio. A los 11 ddi, P. capsici expuesto a extractos de muérdago presentó micelio ralo y bordes asimétricos, acumulándose en el sitio de inoculación y sin esporangios, en comparación con los controles SED y CDE, que mostraron evidencia de formación de esporangios y morfología estrellada típica (Figura 2B). Es decir, los extractos indujeron cambios morfológicos macroscópicos en la morfología colonial y en el desarrollo de esporangios de P. capsici. Sobre esto Flores-Vaillant et al. (2013) encontraron que el oomiceto P. nicotianae mostró micelio ralo, no algodonoso, pobre y con bordes irregulares debido a la adición de extractos de tabaco al medio de cultivo, efecto similar al observado en esta investigación tras la adición de extractos de muérdago. Estos resultados podrían deberse a la plasticidad fenotípica, definida como la capacidad de un organismo para expresar diferentes fenotipos en respuesta a cambios ambientales que conducen al proceso adaptativo. En este caso, la presencia de fitoquímicos en los extractos podría estar induciendo un proceso adaptativo en P. capsici, reflejado en el cambio de la morfología colonial de tipo estrellado a ralo. No obstante, es necesario realizar experimentos adicionales que profundicen en la potencial capacidad adaptativa del patógeno ante la presencia de los componentes de los extractos de muérdago.
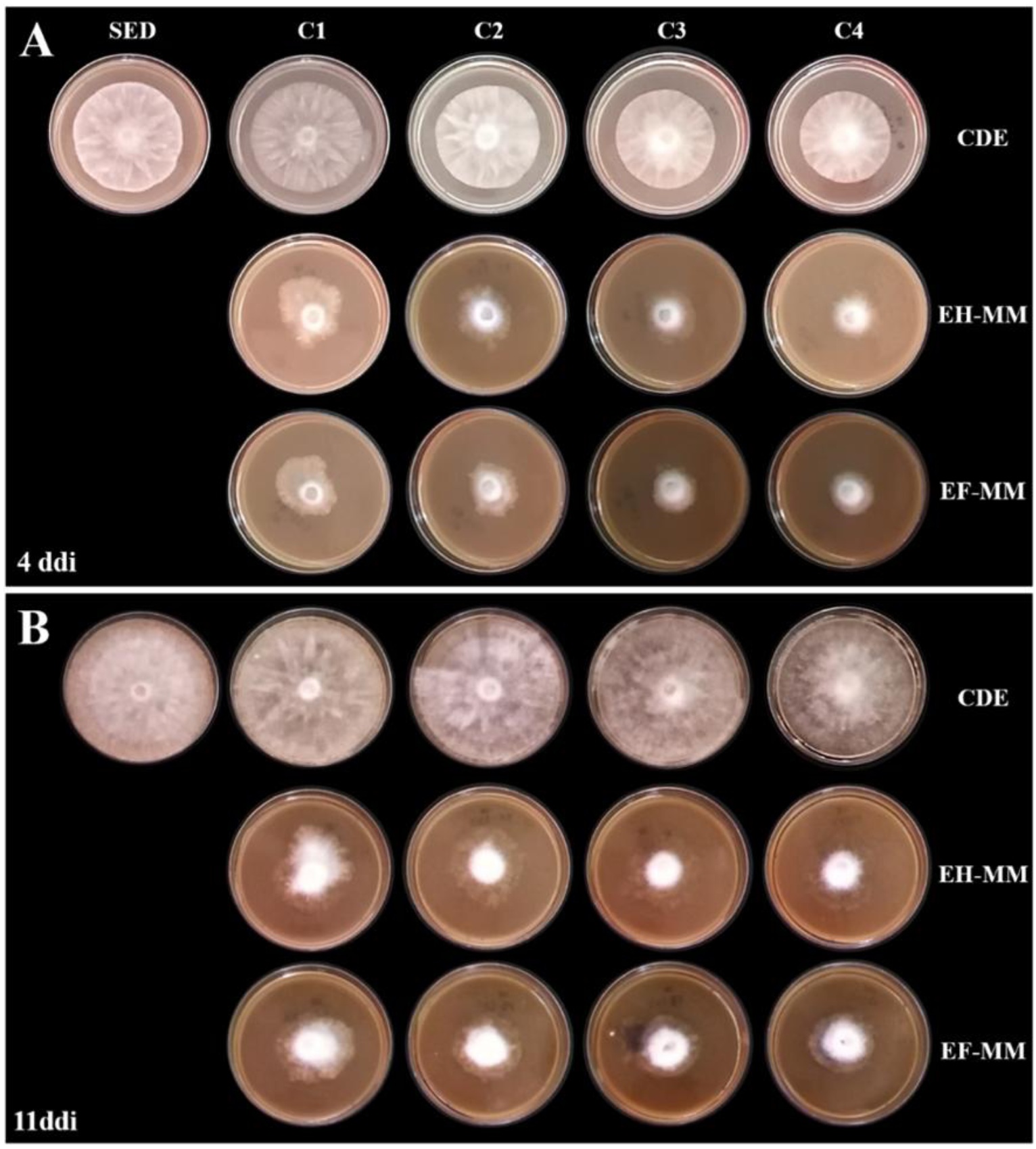
Figura 2 Morfología colonial de P. capsici expuesto a extractos de hojas y flores de muérdago a los 4 (A) y 11 (B) ddi. C1, C2, C3 y C4 concentración 1, 2, 3 y 4 (6.7%, 13.3%, 23.3% y 33.3%) respectivamente, de extractos de hojas, flores de muérdago, y CDE. EH-MM, extracto de hojas de muérdago. EF-MM, extracto de flores de muérdago. SED, sin extractos y disolventes de extracción. CDE, con disolventes de extracción.
Efecto de extractos de muérdago en hifas, esporangios y zoosporas de P. capsici
La evaluación de la morfología microscópica de hifas y esporangios de P. capsici que creció en contacto con los extractos de muérdago durante 4 días mostró que el oomiceto disminuye su capacidad de desarrollar esporangios a concentraciones altas de ambos extractos, sin cambios evidentes en la morfología (Figura 3). Sin embargo, los extractos indujeron engrosamiento de algunas hifas en concentraciones intermedias de EF-MM (C2), pérdida de componentes intracelulares en EF-MM (C3) y adelgazamiento de las hifas en EF-MM (C4).
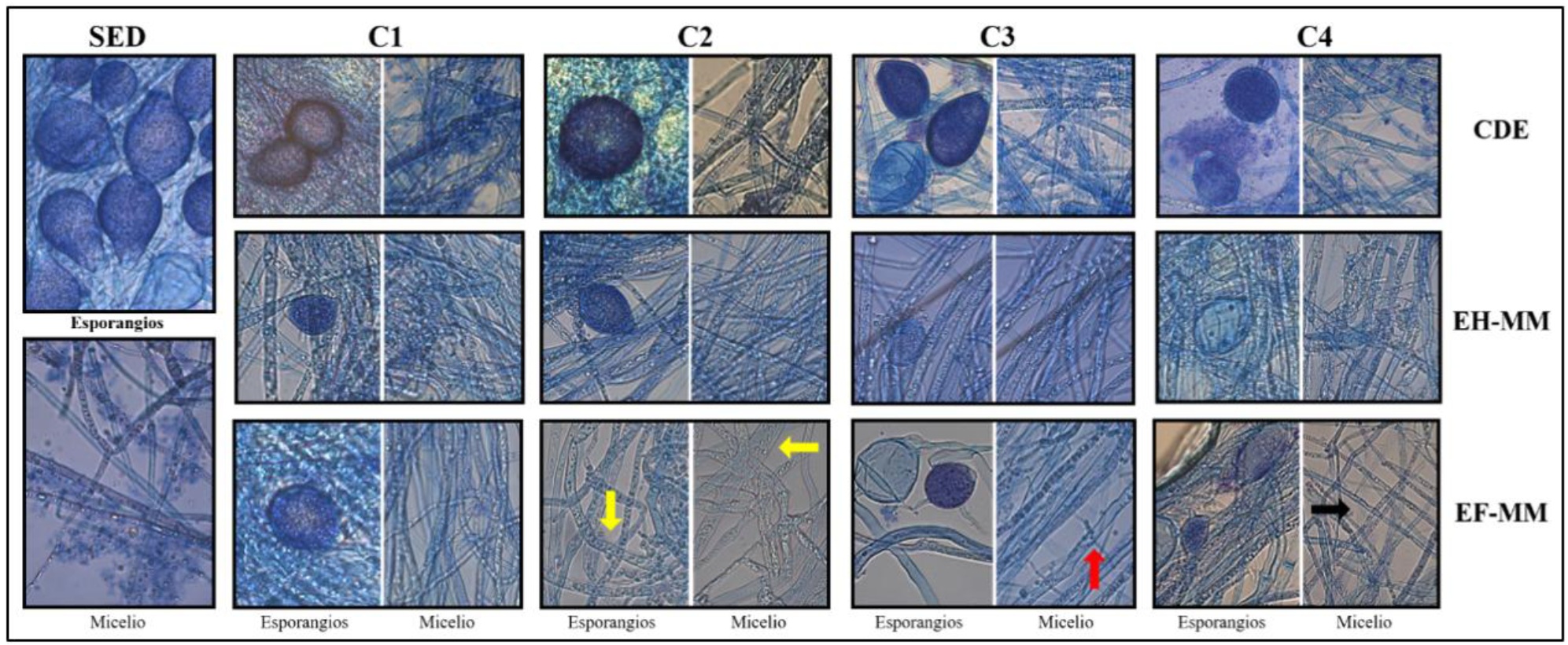
Figura 3 Morfología microscópica de P. capsici cultivado en extractos de muérdago. C1, C2, C3 y C4: concentraciones 1, 2, 3 y 4 (6.7%, 13.3%, 23.3% y 33.3%), respectivamente, de extractos de hojas, flores de muérdago y CDE. EH-MM, extracto de hojas de muérdago. EF-MM, extracto de flores de muérdago. SED, sin extractos ni disolventes de extracción. CDE, con disolventes de extracción. Flechas: amarilla, engrosamiento de hifas. Roja, pérdida de componentes intracelulares. Negra, adelgazamiento de hifas.
La interacción de zoosporas de P. capsici con EH-MM, EF-MM y CDE en C1, C2, C3 y C4 durante 48 h mostró una inhibición de la germinación (Figura 4). Las zoosporas con CDE presentaron flagelos intactos. Sin embargo, ambos extractos de muérdago provocaron daños estructurales en las células al adherirse a la pared de la zoospora en concentraciones bajas (C1, C2), pérdida de la pared celular en C3 y ruptura de la estructura celular en C4 (Figura 4). Es decir, los extractos de muérdago mostraron efectos negativos sobre la motilidad y la estructura de las zoosporas de P. capsici, lo que impidió su germinación.
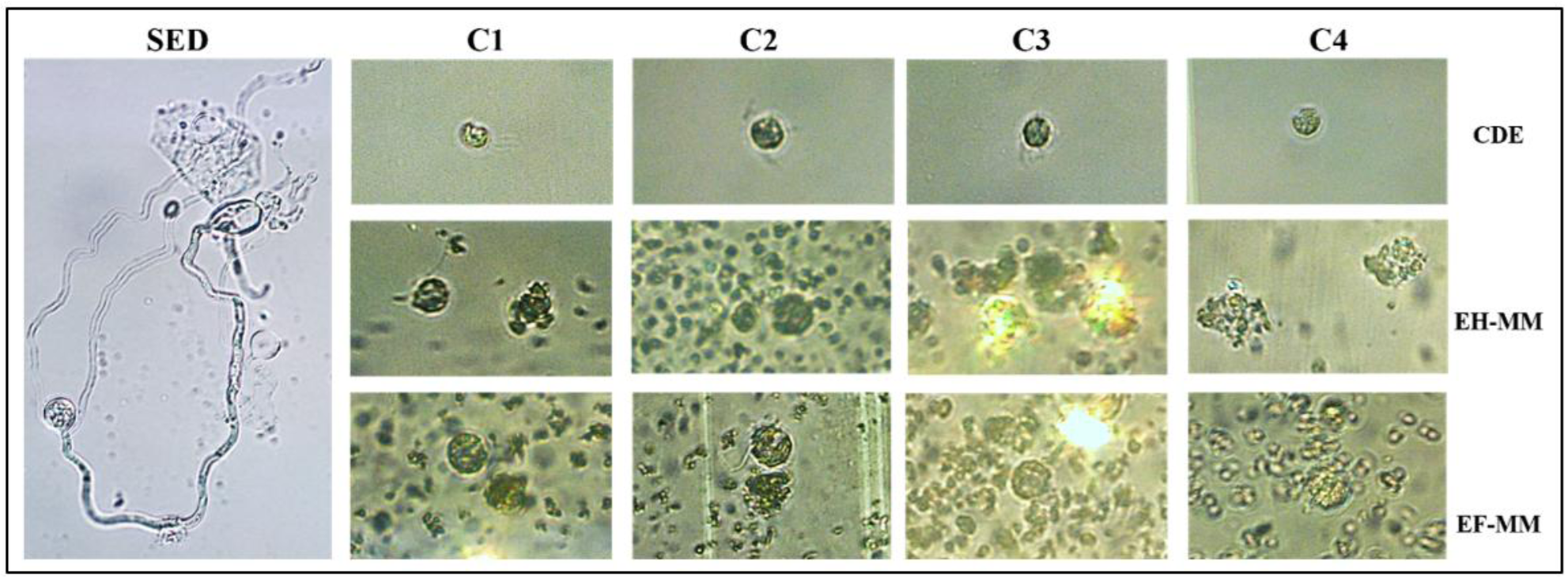
Figura 4 Efecto de los extractos de muérdago sobre las zoosporas de P. capsici. C1, C2, C3 y C4 = concentraciones 1, 2, 3 y 4 (6.7%, 13.3%, 23.3% y 33.3%) de extractos de hojas, flores de muérdago y CDE en suspensión de zoosporas. EH-MM, extracto de hojas de muérdago. EF-MM, extracto de flores de muérdago. SED, sin extractos y disolventes de extracción. CDE, con disolventes de extracción.
Los resultados indicaron que los EH-MM y EF-MM disminuyen la capacidad de producción de esporangios de manera dependiente de la concentración y causan alteraciones estructurales en las hifas de P. capsici, principalmente cuando se cultiva en medio suplementado con EF-MM. Asimismo, ocasionaron engrosamiento de las hifas, pérdida de componentes intracelulares y, en ocasiones, adelgazamiento de estas. Estos resultados son similares a los reportados por Barupal et al. (2019), quienes encontraron inhibición del crecimiento del micelio de Curvularia lunata por extractos de la planta Lawsonia inermis, los cuales indujeron un aumento del ancho de las hifas del patógeno, así como una disminución del tamaño de los conidios. En este mismo estudio, los autores propusieron que el aumento del ancho en las hifas puede deberse a la presencia de metabolitos secundarios en el extracto de la planta, entre los que se incluyen taninos, carbohidratos, saponinas y flavonoides; estos metabolitos pueden inactivar las adherencias del microorganismo, cambiar la permeabilidad de la membrana permitiendo que entre agua y/o pueden acumularse en las hifas de los patógenos de prueba, lo que da como resultado aumento en el ancho o grosor de las hifas. De manera similar, extractos de Hypoxylon anthochroum indujeron cambios morfológicos sobre la densidad en el micelio y las hifas de P. capsici (Sánchez-Fernández et al. 2020), en donde la isobenzofuranona fue considerada como la principal molécula que indujo cambios en la morfología de la colonia y de las hifas del patógeno, ya que el micelio tratado con este compuesto mostró hifas sin vacuolas, alteración de la permeabilidad de la membrana en las hifas y adelgazamiento de las hifas, la vacuolización en las hifas y pérdida del contenido de hifas, similar a lo encontrado en esta investigación.
Las zoosporas participan de manera importante en la diseminación e infección por P. capsici (Lamour et al. 2012), por lo que cualquier molécula que inhiba significativamente la producción y la germinación de zoosporas debería ser una candidata para controlar la enfermedad causada por el patógeno (Matheron y Porchas 2000). En este trabajo se determinó que los tratamientos con EH-MM y EF-MM disminuyen el desarrollo de las zoosporas. Debido a que las enfermedades del tizón por Phytophthora se originan a partir de las zoosporas y luego se propagan rápidamente bajo condiciones favorables (Li et al. 2019), la actividad de los extractos de muérdago observada en las zoosporas de P. capsici podría estudiarse a detalle con diferentes concentraciones de moléculas purificadas para establecer un potencial aplicación para el control de la enfermedad. Sin embargo, esto contrasta con lo encontrado por Xoca-Orozco et al. (2022), quienes recuperaron esporas y conidios de Fusarium sp., Curvularia sp. y C. gloesporoides de la interacción con extractos de hojas y flores de muérdago, los cuales mostraron crecimiento reducido, por lo que consideraron que los extractos de muérdago tienen un efecto fungistático sobre estos patógenos. Es posible que el efecto de EH-MM y EF-MM varíe según el tipo de fitopatógeno filamentoso que se intenta controlar.
Morfología y recuperación de P. capsici después de exponerse a extractos de muérdago
A los siete días de crecimiento en medio V8 adicionado con los extractos de muérdago, P. capsici se transfirió a medio de cultivo V8 sin extractos, lo que indicó que el oomiceto se recupera del efecto de los extractos en un 87 a 97% en cinco días (Tabla 3).
Tabla 3 Recuperación de P. capsici tras el crecimiento en medio V8 suplementado con extractos de muérdago.
| C1 (6.7%) | C2 (13.3%) | C3 (23.3%) | C4 (33.3%) | |
|---|---|---|---|---|
| EH-MM | 87.50 ± 1.77* | 91.88 ± 4.42 | 95 ± 0 | 97.5 ± 3.54* |
| EF-MM | 95.63 ± 2.65 | 93.13 ± 0.88 | 93.13 ± 0.88 | 93.13 ± 2.65 |
EH-MM, extracto de hojas de muérdago. EF-MM, extracto de flores de muérdago. * diferencia significativa p ≤ 0.05. Concentraciones 1, 2, 3 y 4 de EH-MM y EF-MM (C1, C2, C3 y C4). Como testigo, se utilizó P. capsici, que no creció en medio de cultivo adicionado con alguna concentración de EH-MM o EF-MM, y su crecimiento se consideró como el 100%.
No se encontraron diferencias significativas (Tukey, p ≤ 0.05) en la recuperación del oomiceto entre EH-MM y EF-MM en las cuatro concentraciones evaluadas. Sin embargo, hubo diferencia entre C1 y C4 en EH-MM. Sorprendentemente, en ambas condiciones evaluadas se observó que, en concentraciones bajas de los extractos (C1), el porcentaje de recuperación del crecimiento de P. capsici fue menor y aumentó progresivamente a concentraciones cada vez mayores. En cambio, la recuperación de P. capsici en EF-MM fue similar en C2, C3 y C4. No se incluyó la evaluación de la recuperación de P. capsici mediante disolventes de extracción, ya que durante la interacción estos no mostraron efecto inhibitorio sobre el crecimiento del patógeno. P. capsici pigmentó el medio de cultivo en contacto con los extractos de muérdago, por lo que la intensidad de pigmentación fue proporcional a la concentración evaluada (Figura 5A). No obstante, cuando el patógeno se pasó a placas de Petri sin extractos de muérdago, no pigmentó el medio de cultivo en ningún tratamiento (Figura 5B), lo que evidenció que los extractos de muérdago indujeron la pigmentación del medio de cultivo.
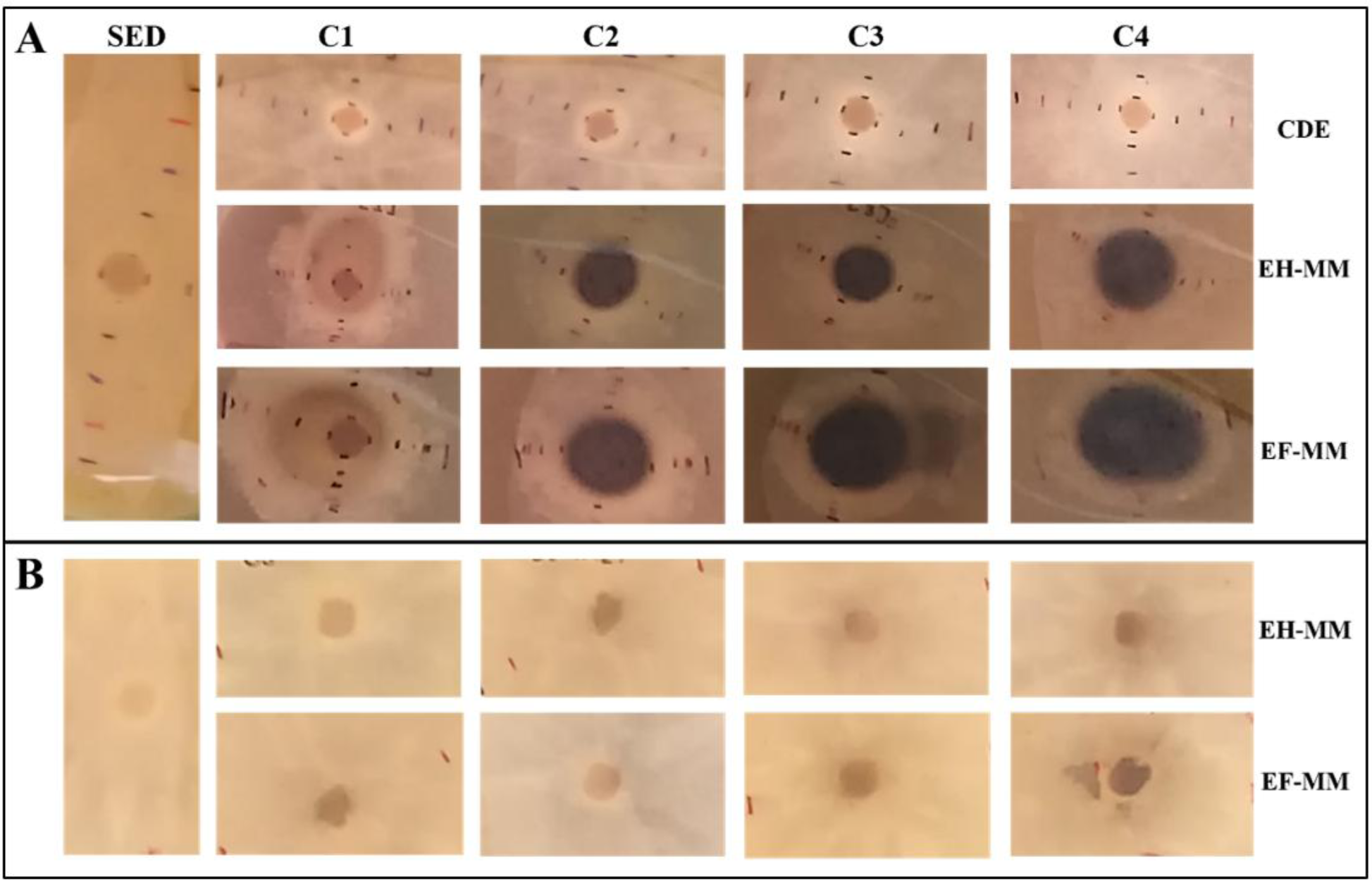
Figura 5 A, pigmentación del medio de cultivo ocasionada por P. capsici inducida por extractos de muérdago. B, ausencia de pigmentación del medio de cultivo tras el crecimiento de P. capsici con extractos de muérdago. C1, C2, C3 y C4: concentraciones 1, 2, 3 y 4 (6.7%, 13.3%, 23.3% y 33.3%), respectivamente, de extractos de hojas, flores de muérdago y CDE. EH-MM, extracto de hojas de muérdago. EF-MM, extracto de flores de muérdago. SED, sin extractos ni disolventes de extracción. CDE, con disolventes de extracción.
Se observó recuperación de la morfología, comúnmente estrellada en la mayoría de los tratamientos de P. capsici, después de la interacción con extractos de muérdago, en ocasiones con bordes asimétricos y pérdida de definición de los picos de la estrella (Figura 6). El mayor cambio morfológico se observó en EH-MM C3, con morfología rayada.
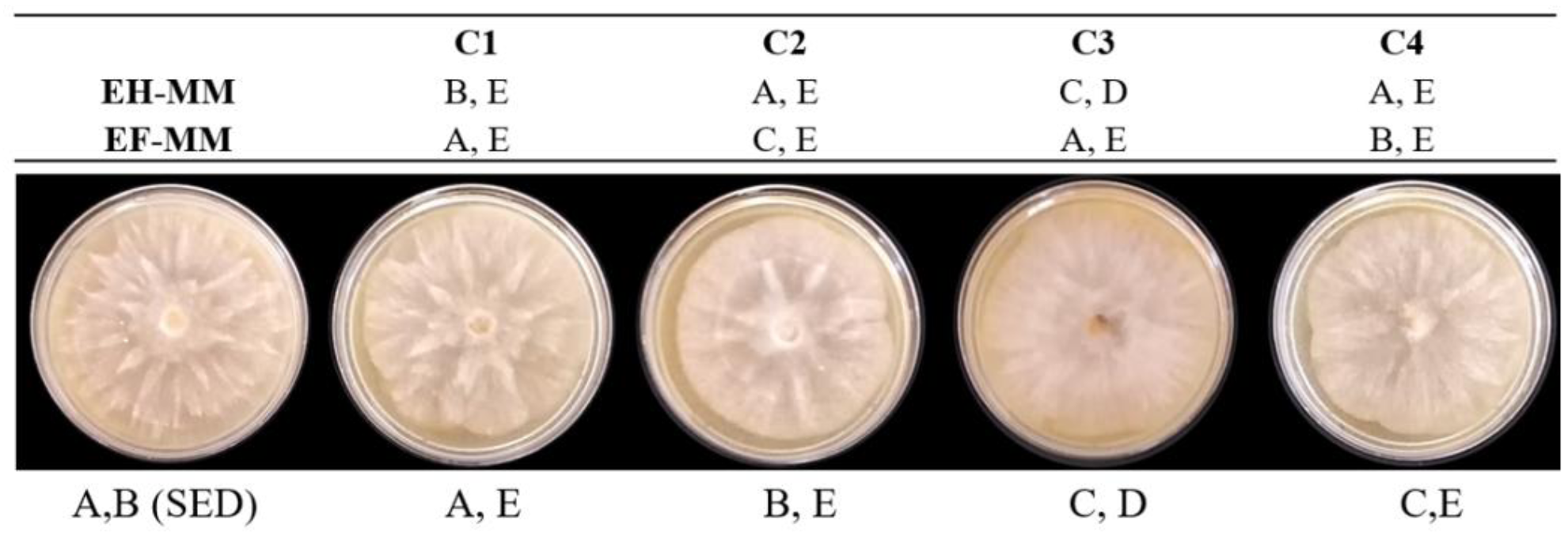
Figura 6 Morfología colonial de P. capsici tras la exposición a extractos de muérdago. C1, C2, C3 y C4 = concentraciones 1, 2, 3 y 4 (6.7%, 13.3%, 23.3% y 33.3%) de extractos de hojas y de flores de muérdago. EH-MM, extracto de hojas de muérdago. EF-MM, extracto de flores de muérdago. A, Morfología colonial estrellada. B, morfología colonial estrellada con picos no definidos. C, Morfología colonial rayada. D, bordes simétricos. E, Bordes asimétricos. SED, medio V8 sin extractos ni disolventes de extracción (A, B).
Los resultados indicaron una recuperación del 87 al 97% en la tasa de crecimiento de P. capsici después de estar expuesto a cuatro concentraciones de EH-MM y EF-MM, lo que señala la capacidad resiliente del oomiceto después de la exposición a los extractos de muérdago. En el futuro, se deben dilucidar los mecanismos moleculares por los cuales los extractos de muérdago afectan negativamente al oomiceto y su capacidad de recuperación después de la exposición a dichos extractos. Por tanto, para monitorear de manera más efectiva la resistencia de los fitopatógenos en el futuro, es crucial establecer de manera sólida la sensibilidad de referencia de un nuevo oomiceticida antes de su uso comercial generalizado (Russel 2004).
Germinación y desarrollo de zoosporas de P. capsici después de exposición a extractos de muérdago
Después de la exposición de zoosporas a los extractos de muérdago y CDE, se lavaron e inocularon en medio V8; dichas células germinan y desarrollan micelio cuando no son tratadas con extractos de muérdago y disolventes de extracción (SED) a los 7 ddi. Sin embargo, en las cuatro concentraciones de CDE, EH-MM y EF-MM, las zoosporas no germinaron.
Los extractos de hojas y flores de muérdago modificaron el crecimiento y el desarrollo de P. capsici, alteraron la morfología colonial y la estructura de las hifas y redujeron la formación de esporangios. Estos resultados evidencian el potencial de los extractos de muérdago o de las moléculas que contienen como base para el desarrollo de estrategias de control de las enfermedades causadas por Phytophthora. Además, los extractos de muérdago tienen efecto zoosporicida, por lo que podrían prevenir la propagación de la enfermedad mediante la diseminación de esporas. Sin embargo, P. capsici mostró una capacidad de recuperación relativamente rápida tras la exposición a los extractos de muérdago, lo que sugiere mecanismos adaptativos o de tolerancia.

