











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El lince rojo, también conocido como gato montés (Lynx rufus Schreber, 1777), habita en ecosistemas desérticos, tropicales y de montaña (Gómez-Naranjo et al. 2017, CONABIO 2024), desde los 0 hasta los 3 600 msnm, y se distribuye desde el sur de Canadá hasta el sur de México (López-González et al. 2015). La presencia de este felino es indicadora de la buena calidad del hábitat, ya que está asociada a la alta cantidad de presas disponibles para su sobrevivencia (Sánchez-González et al. 2018).
En México se registra en 27 de los 32 estados (CONABIO 2009, Bárcenas y Medellín 2021), sin embargo, existen escasas investigaciones sobre su densidad poblacional a nivel nacional. Al respecto, Vega-Flores y Farías-González (2021), en una reserva de la biosfera caracterizada por selva baja caducifolia y matorral xerófilo en el estado de Puebla; y López-González et al. (2015), evaluaron la densidad del lince rojo en Querétaro, dentro de un área protegida dominada por matorral crasicaule, bosque tropical caducifolio y bosque de encino. Un estudio relevante por la amplia cobertura es el de Bárcenas y Medellín (2021), quienes estimaron la distribución potencial y densidad del lince rojo en seis estados del noroeste, centro y sur de México.
Aunque la especie no se encuentra en ningún estado de protección a nivel nacional o internacional, se ha documentado que las poblaciones locales se ven afectadas debido a la fragmentación de su hábitat (Sánchez-González et al. 2018, Luja et al. 2021). La estructura y composición vegetal de los fragmentos resultantes del cambio de uso de suelo pueden influir en su uso y selección por el lince rojo. El uso se refiere al tiempo que pasa dentro del área, mientras que la selección, es el proceso de preferencia a cierto tipo de hábitat (Abouelezz et al. 2018). La preferencia puede estar influenciada por las características de la matriz donde se encuentra, como el clima, cobertura vegetal, alimento disponible, competencia entre especies o la influencia antropogénica (Herrera 2011). En el caso del lince rojo, se ha documentado que su presencia se encuentra condicionada principalmente por una densa cobertura vegetal que proporcione refugio y escape, así como la disponibilidad de presas (IUCN 2016).
El Matorral Espinoso Tamaulipeco (MET) es un ecosistema del noreste de México dominado en el estrato alto por especies leñosas de carácter espinoso. Este presenta una gran diversidad de especies, así como un alto grado de endemismo, en relación con sus condiciones climáticas extremas (Mata et al. 2025). Es hábitat de mamíferos como el lince rojo, que cumple su función como mesodepredador al mantener el control y estabilidad de sus presar, compuestas principalmente de lagomorfos o pequeños roedores (Aranda 2012, Luja et al. 2021). Sin embargo, en los últimos años esta comunidad vegetal ha sido altamente fragmentada por actividades agropecuarias, de minería y urbanización, por lo que hoy en día se estima un remanente del 20.4% de la superficie original (Molina-Guerra et al. 2019, CONABIO 2024). Considerando la preferencia de hábitat del lince rojo a hábitats de mayor cobertura forestal, es importante evaluar la respuesta de la especie en hábitats antropizados que se encuentran fragmentados.
Por lo anterior, el presente trabajo tiene por objetivo determinar la densidad poblacional, así como la probabilidad de ocupación y detección de Lynx rufus en fragmentos del matorral espinoso tamaulipeco del noreste de México. Las hipótesis planteadas son que se presentará una baja densidad poblacional debido a la fragmentación del hábitat, y que la variable que más influye en la probabilidad ocupación y detección de L. rufus es la cobertura arbórea. Evaluar el estado del lince rojo en este ecosistema contribuirá a comprender su respuesta ante la fragmentación, lo que permitirá elaborar estrategias de manejo que incrementen su conservación a largo plazo.
Materiales y métodos
Área de estudio
El sitio de estudio se ubica dentro del municipio de Reynosa, Tamaulipas, México (25° 13´ 15.3´´ LN y -98° 12´ 51.33´´ LO), con una superficie de 140.31 km2. De acuerdo con la clasificación de Köppen modificada por García (2004), el clima es del tipo BS1 (h’) hx’, que corresponde a semiseco cálido, con una precipitación media anual que fluctúa entre los 500 y 700 mm, y una temperatura media anual de 22 °C. Los tipos de suelo presentes son Castañozem, Vertisol y Xerosol (INEGI 2020). Los usos de suelo se clasifican como: Agricultura de Temporal Anual, Pastizal Cultivado Permanente, Pastizal Inducido, Agricultura de Riego Anual, Vegetación Secundaria de Matorral Espinoso Tamaulipeco y Matorral Espinoso Tamaulipeco (INEGI 2013). Dentro del sitio se encuentra ubicado un parque eólico que cuenta con 103 aerogeneradores, separados 1 km entre sí.
Monitoreo e identificación del lince rojo
Se realizaron recorridos dentro de los corredores de vegetación para identificar la presencia del lince rojo a través de huellas y excretas, siguiendo la metodología propuesta por Aranda (2012). Se realizaron en total 35 recorridos entre los días 15 al 19 de marzo, durante un horario de 7:00 a 13:00 horas. Una vez identificada la presencia de la especie en los corredores, se determinaron las áreas potenciales de muestreo ubicándolas cerca a los caminos y senderos, considerando su accesibilidad y la facilidad para encontrar presas (Heilbrun et al. 2003, Sirén et al. 2017). Para determinar la ubicación final de cada estación se consideró una distancia de 500 a 800 m entre ellas (UNAM-INECOL 2009), con una densidad de 0.21 estaciones km-2.
Se establecieron 30 estaciones de muestreo (Figura 1), cada una formada por dos cámaras trampa Bushnell de 30 Mega Pixeles (Core DS Low Glow Trail) ubicadas una frente a otra paralelamente al camino. Las cámaras se colocaron a una altura de 0.25 a 0.45 m del suelo, ajustadas a soportes de madera. Se programaron para trabajar 24 horas al día, tomando las fotos con un minuto de retraso entre disparos, registrando la fecha y hora de captura y omitiendo el uso de cebos o atrayentes (Sánchez-González et al. 2018).
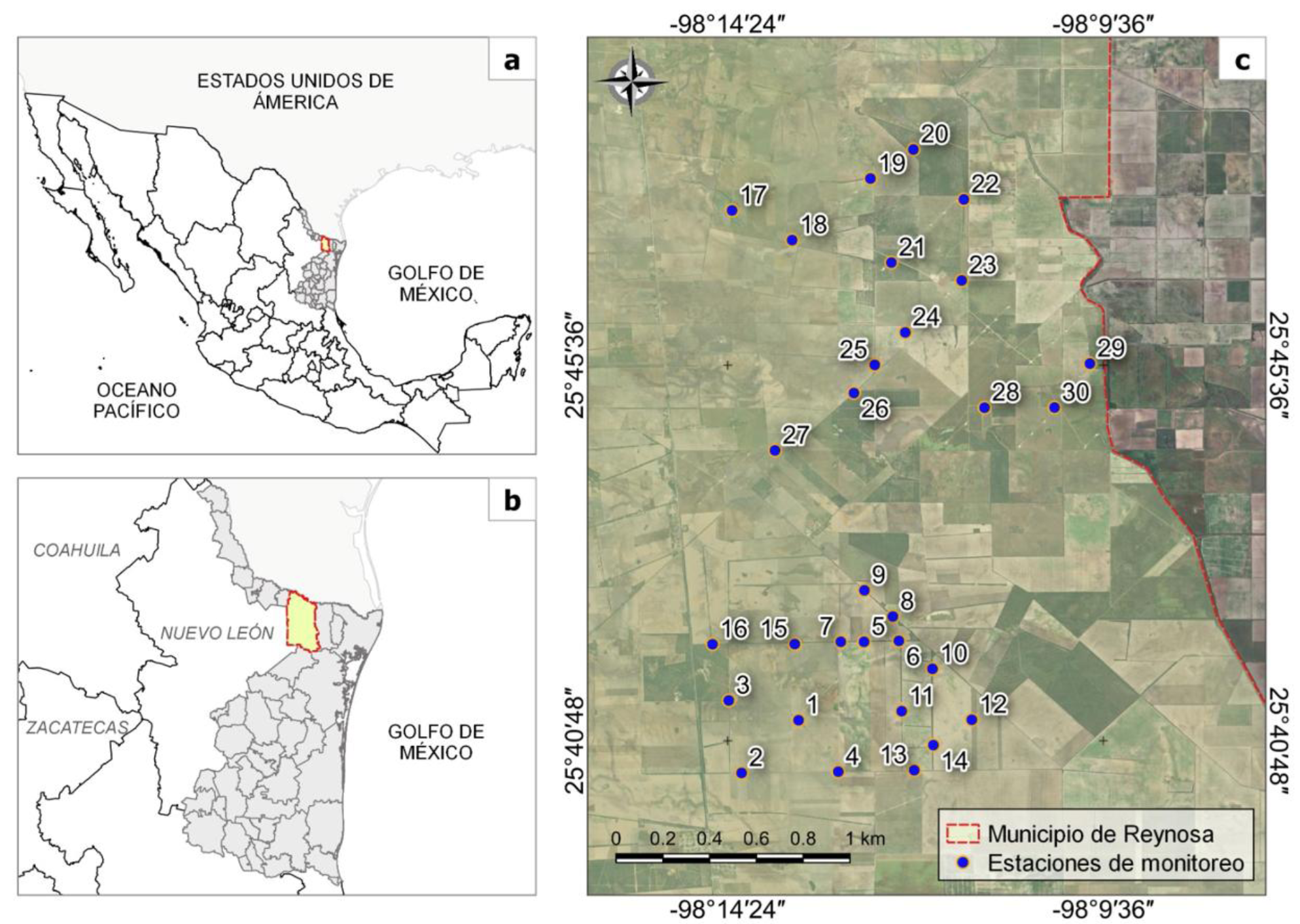
Figura 1 Ubicación de las estaciones de foto trampeo dentro del sitio de estudio. a) Mapa de México indicando el estado de Tamaulipas, b) Municipios de Tamaulipas indicando el área de estudio y c) área de estudio indicando las estaciones de foto trampeo.
El muestreo y mantenimiento de las cámaras trampa se realizó mensualmente durante 360 días, en el periodo comprendido entre el 1 de marzo del 2021 al 28 de febrero del 2022. Estas actividades consistieron en la limpieza de la estación (remoción de la vegetación en el ángulo de las fotos) e intercambio de tarjetas de memoria, así como la programación y ajuste de las cámaras.
Para el análisis de las fotografías, los individuos de lince rojo fueron identificados de acuerdo con Heilbrun et al. (2006). Se tomaron en cuenta rasgos distintivos tales como cicatrices, porte corporal, características faciales y patrones de manchas en el cuerpo, patas y orejas, considerando las posibles variaciones que pudieran presentarse por la postura del lince rojo (Figura 2). Este análisis se realizó de forma cruzada mediante un consenso de opiniones entre los autores de la presente investigación. Los individuos identificados fueron descritos y nombrados para su posterior seguimiento poblacional. Si la fotografía lo permitía, se identificó el sexo y la edad estimada. Se agruparon aquellas fotografías que presentaran un mismo flanco con el fin de obtener secuencias de los individuos en diferentes posiciones. Para evitar un sobre conteo, se consideraron registros independientes a las fotografías tomadas con una separación de 24 horas.

Estimación de la abundancia y densidad utilizando un modelo Captura-Recaptura (MCR)
Se utilizaron los registros fotográficos para generar un modelo de captura-recaptura con el fin de estimar estadísticamente la abundancia y densidad poblacional. Se llevó a cabo la clasificación de las fotografías considerando como captura a aquellos individuos que presentaran al menos una característica que no se hubiera observado en fotografías anteriores. Se descartaron las fotografías de baja calidad donde no pudieran identificarse rasgos del individuo, y se utilizaron al menos tres rasgos característicos identificados en individuos registrados para clasificarlos como recaptura (Heilbrun et al. 2003). Se elaboró una base de datos, en donde se cuantificaron los individuos y analizaron a la par sus patrones de desplazamiento y comportamiento a través del registro de capturas y recapturas por estación de muestreo.
Los datos se vaciaron en una matriz de presencia (1) o ausencia (0), donde se agregaron los individuos capturados en renglones, y los días de esfuerzo de muestreo en columnas. Se elaboraron 12 matrices correspondientes a la información de cada uno de los meses que abarcó el periodo de muestreo. Con el fin de respetar el supuesto de una población cerrada para el análisis de captura-recaptura, se descartaron los meses donde se hubieran registrado muertes o nacimientos. Por lo tanto, únicamente se analizaron los meses de noviembre del 2021 a febrero del 2022 para el análisis. Estos datos se analizaron mediante la herramienta CAPTURE dentro del programa computacional MARK, donde se determina el modelo estadístico más adecuado para la información recolectada (Cooch y White 2017). Las opciones son: el modelo nulo (Mo), de heterogeneidad a la captura (Mh), de respuesta a la captura (Mb), de variación temporal en la probabilidad de captura (Mt) o la combinación de todos estos, siendo el más apropiado el que arroje el resultado más cercano al valor 1, de acuerdo con el estimador Jacknife. Con estos datos, el programa estimó la abundancia absoluta con un error estándar y un intervalo de confianza determinados (López-González et al. 2015, Bárcenas y Medellín 2021).
Posteriormente, se calculó y promedió la distancia máxima de desplazamiento lineal (mean maximum distance moved o MMDM) recorrida por cada lince rojo entre 2 cámaras (Otis et al. 1978). Mediante el programa QGIS (QGIS Desktop 3.22.15) se ubicaron como puntos centrales las estaciones de muestreo y se agregó en cada una un buffer circular con un radio igual al valor de MMDM. Los círculos resultantes se disolvieron en un solo polígono cuya área se asignó como equivalente al Área Efectiva de Muestreo o ETA por sus siglas en inglés. Estos valores se utilizaron para calcular la densidad poblacional (Individuos km-2), al dividir la abundancia promedio resultante de los análisis de MARK CAPTURE entre el ETA (DiBitetii et al. 2006, Vega-Flores y Farías-González 2021).
Evaluación de los usos de suelo y la vegetación
Los usos de suelo dentro del sitio de estudio se clasificaron mediante el Índice de Vegetación de Diferencia Normalizada (NDVI por sus siglas en inglés), usando el software QGIS (QGIS Desktop 3.22.15). Se utilizó una imagen del 06 de enero del 2022 del satélite Sentinel-2B para realizar una clasificación supervisada de los distintos usos de suelo. Del mapa ráster obtenido, se realizó una digitalización directa en pantalla para obtener un mapa en formato shapefile con las distintas categorías de uso. La información obtenida se corroboró con los recorridos en campo posteriores. Posteriormente se calculó el porcentaje de superficie que representaban los usos de suelo agrícola, pastizal y vegetación del MET dentro del área de influencia del sitio de muestreo determinado por la superficie calculada del MMDM descrita anteriormente (veg).
Debido a que las estaciones de foto trampeo del lince rojo se ubicaron en los fragmentos y corredores de vegetación, se evaluó su composición florística y estructura mediante el establecimiento de 30 sitios de muestreo. Los sitios se ubicaron considerando el centro las estaciones de foto trampeo y presentaban una superficie de 75 m2. Se consideraron los ejemplares del estrato arbóreo-arbustivo que presentaron una altura superior a los 2 m. Las especies se identificaron taxonómicamente y se midieron las variables dendrométricas de altura total, diámetro de copa y diámetro a la altura del pecho. Con esa información se determinaron los parámetros de abundancia, cobertura y frecuencia (Alanís-Rodríguez et al. 2020).
Estimación de la probabilidad de ocupación
Con el fin de complementar el MCR, se elaboró un modelo de ocupación mediante el programa computacional PRESENCE (PRESENCE 2.13.47). El MCR se utiliza para dar un estimado del número de individuos que no fueron contados, a partir de aquellos que si fueron registrados al menos una vez. Por su parte, el modelo de ocupación se basa en estimar, a través de la probabilidad, las tazas de ocupación cuando la probabilidad de detección es menor a 1, a través de covariables y con el supuesto de que el tamaño de la población ya es conocido. Esto considerando que, el no detectar la especie en las cámaras trampa no significa que la especie no se encuentre presente en el sitio (MacKenzie et al. 2002).
Las covariables que suelen ser utilizadas en los modelos de ocupación se suelen relacionar con factores ambientales o con impactos humanos (Isasi-Catalá et al. 2016). En el presente estudio, para la probabilidad de ocupación (ψ) se utilizaron las siguientes covariables ambientales: abundancia (abun), altura promedio (alt), cobertura (cob), diversidad vegetal (div). Aquellas relacionadas con el impacto humano incluyeron la proximidad en metros lineales del aerogenerador más cercano (dist), así como el porcentaje de superficie de uso de suelo agrícola (agri), pastizal (past), o cobertura vegetal del MET (veg), calculado previamente.
Para la probabilidad de detección (p), se utilizó la temperatura promedio diaria (temp), además de las mismas covariables que ψ, considerando el impacto que pueden ejercer sobre la detección de la especie en las cámaras trampa. Se consideró el coeficiente de la variable (Beta) para evaluar el efecto de las covariables en la probabilidad de ocupación de la especie. Se analizaron y compararon únicamente aquellos modelos óptimos cuyo delta del Criterio de Información de Akaike modificado para muestras pequeñas (ΔAICc) fuera ≤ 3 (MacKenzie et al. 2006).
Resultados
Durante los 365 días del monitoreo, se identificaron 23 linces rojos mediante 308 capturas-recapturas. Se obtuvieron recapturas de todos los linces rojos en al menos dos estaciones diferentes, con excepción de dos individuos que se capturaron en una sola estación. Las estaciones 17 y 18 (Figura 1) presentaron la mayor cantidad de registros, con 41 y 40 cada una.
Referente a los patrones de actividad, las capturas muestran movimiento de los individuos a lo largo del día. No obstante, el mayor número de capturas se llevó a cabo entre las 18:00 y 07:00 horas, con picos de actividad entre 01:00 y 02:00 horas. Entre las 14:00 y 15:00 horas no se registró actividad durante todo el periodo de monitoreo. El mes con mayor número de registros corresponde a noviembre del 2021 (53), seguido de febrero (45) y enero (41) del año siguiente. Los meses con los registros más bajos fueron julio (11) y junio (13) del 2021.
Aunque no fue posible identificar el sexo de todos los individuos, se logró confirmar la identidad de dos hembras. Una de ellas se registró con tres crías durante los meses de septiembre y octubre del 2021 entre las estaciones 17 y 18. Sin embargo, no se volvió a detectar en los meses siguientes. Este fue el único individuo que se registró con cinco presas diferentes de lagomorfos y roedores de especies no identificadas.
Las mayores distancias de desplazamiento calculadas fueron de 13.81, 11.53 y 10.96 km, recorridas por tres linces rojos diferentes. Se obtuvo un valor de MMDM de 4.75 km, a partir del cual se calculó una superficie del ETA de 322.32 km2. Sin embargo, al exceder este valor la superficie del área de estudio, el valor del ETA fue ajustado a este último, dando un total de 140.31 km2. De acuerdo con MARK CAPTURE, se estimó una abundancia poblacional promedio de 12 individuos (Tabla 1), indicando una densidad promedio de 8 individuos 100 km-2. El modelo más adecuado para todos los periodos fue el de heterogeneidad a la captura para una población cerrada (Mh = 1.0).
Tabla 1 Resultados de abundancia poblacional obtenidos de la herramienta MARK-CAPTURE y Densidad estimada por periodo de muestreo. PC = Probabilidad de captura, A = Abundancia poblacional (ind), EE = Error estándar. IC = Intervalo de confianza con un 95 %.
| Periodo | Modelo | PC | A | EE | IC (95 %) | Densidad (Individuos 100 km-2) |
|---|---|---|---|---|---|---|
| 1/11/2021 al 30/11/2021 | Mh = 1.0 | 0.07 | 15 | 1.33 | 15 a 21 | 10 |
| 1/12/2021 al 31/12/2021 | Mh = 1.0 | 0.11 | 10 | 0.52 | 10 a 10 | 7 |
| 1/01/2022 al 31/01/2022 | Mh = 1.0 | 0.09 | 11 | 0.71 | 11 a 11 | 7 |
| 1/02/2022 al 30/02/2022 | Mh = 1.0 | 0.07 | 15 | 1.54 | 15 a 22 | 10 |
Dentro del área de estudio se encontraron 44.22 km2 de actividades agrícolas-ganaderas, distribuidas en la matriz paisajística y dentro de los parques eólicos. Las actividades agrícolas conformaron una superficie de 27.85 km2, mientras que la superficie de pastizal para usos ganaderos fue de 16.37 km2.
La superficie cubierta por el MET en el área estudiada fue de 28.94 km2. La composición florística del matorral se constituyó por 47 especies agrupadas en 23 familias taxonómicas. Las familias que presentaron mayor número de especies fueron Fabaceae (8), Poaceae (6) y Cactaceae (5), representadas por las especies Celtis pallida, Neltuma glandulosa, Forestiera angustifolia y Ebenopsis ebano.
Referente a las variables evaluadas dentro de los sitios de muestreo, la cobertura vegetal varía de 679 m2ha-1 (sitio 4), hasta los 4 004 m2ha-1 (sitio 18). Los valores de abundancia contienen un rango más amplio de valores, oscilando desde 1 733 (sitio 5), hasta los 18 133 Individuos ha-1 (sitio 14). Se observa que la mayor diversidad de especies vegetales se encuentra en el sitio 24 con 17 especies, mientras que el sitio 21 solo presenta 2 especies.
Se obtuvieron cuatro modelos óptimos de ocupación en base a su ΔAIC (Tabla 2). En estos se observa que se repiten las mismas siete covariables para p (veg*past*temp*cob*abun*div*alt). Por lo que, de acuerdo con los modelos, estas covariables influyen en la detección del lince rojo. De acuerdo con los resultados obtenidos en el modelo óptimo, existe mayor probabilidad de ocupación en aquellos sitios que presentan mayor cobertura arbórea (Beta = 1.30, D.E = 0.69), y que se encuentran más alejados de los aerogeneradores (Beta = 1.73, D.E = 1.40). Se estimó que los sitios con mayor probabilidad de ocupación son los enumerados como 26 (ψ = 0.9995, D.E. = 0.0025), 9 (ψ = 0.9977, D.E. = 0.0097) y 7 (ψ = 0.9955, D.E. = 0.0163).
Tabla 2 Sitios de muestreo, Número de avistamientos de Lynx rufus, número de especies vegetales, abundancia vegetal (N ha-1) y cobertura de copa (m2 ha-1) en cada sitio de muestreo.
| Sitio de muestreo | Avistamientos de Lynx rufus | Número de especies vegetales | Abundancia vegetal (N ha-1) | Cobertura de copa (m2ha-1) |
|---|---|---|---|---|
| 1 | 0 | 5 | 4 400 | 1 694 |
| 2 | 5 | 10 | 7 067 | 1 941 |
| 3 | 8 | 9 | 8 533 | 1 897 |
| 4 | 0 | 6 | 2 533 | 679 |
| 5 | 5 | 5 | 1 733 | 1 387 |
| 6 | 3 | 6 | 4 667 | 742 |
| 7 | 20 | 7 | 8 000 | 2 110 |
| 8 | 20 | 12 | 7 467 | 3 535 |
| 9 | 15 | 13 | 11 600 | 2 106 |
| 10 | 8 | 7 | 10 000 | 1 739 |
| 11 | 6 | 9 | 7 867 | 2 051 |
| 12 | 1 | 4 | 6 667 | 1 363 |
| 13 | 16 | 9 | 14 267 | 1 973 |
| 14 | 13 | 11 | 18 133 | 1 955 |
| 15 | 2 | 10 | 3 867 | 3 085 |
| 16 | 19 | 13 | 6 133 | 3 634 |
| 17 | 41 | 9 | 3 333 | 2 187 |
| 18 | 42 | 12 | 3 733 | 4 004 |
| 19 | 16 | 11 | 8 000 | 2 585 |
| 20 | 0 | 12 | 4 267 | 2 649 |
| 21 | 4 | 2 | 2 133 | 1 189 |
| 22 | 7 | 10 | 7 867 | 2 938 |
| 23 | 17 | 10 | 8 800 | 2 582 |
| 24 | 13 | 17 | 11 067 | 3 058 |
| 25 | 23 | 6 | 2 400 | 1 321 |
| 26 | 12 | 13 | 6 000 | 2 465 |
| 27 | 20 | 11 | 8 800 | 2 496 |
| 28 | 0 | 8 | 6 267 | 2 415 |
| 29 | 6 | 4 | 4 800 | 1 380 |
| 30 | 3 | 8 | 4 533 | 1 822 |
| Promedio | 12 | 9 | 6 831 | 2 166 |
La probabilidad de detección se asociada positivamente con áreas de | 1.06, D.E = 0.24) y con mayor cobertura arbórea (Beta = 0.55, D.E = 0.24). La superficie de cobertura agrícola no es una covariable que tenga efecto en la ocupación o detección de la especie, al no aparecer en estos modelos.
Discusión
La densidad poblacional estimada en este estudio fue de 6-8 individuos por cada 100 km2 para un Área Efectiva de Muestreo (AEM) de 291.505 km2. Bárcenas y Medellín (2021) evaluaron cinco localidades en México y reportaron valores entre 5-53 individuos 100 km-2, lo que demuestra una amplia variación en la densidad del Lynx rufus en México. Resultados similares se han documentado en los estados de México, Puebla y Sonora, con valores de 5.3 (AEM de 56 564 km2), 6.5 (AEM de 15 289 km2) y 7.5 individuos 100 km-2 (AEM de 26,316 km2), respectivamente (Bárcenas y Medellín 2021). Estas investigaciones, al igual que la presente, se desarrollaron en zonas con cercanía a áreas urbanas, lo que podría explicar las densidades relativamente bajas observadas en las poblaciones de Lynx rufus.
En contraste, estudios realizados en Áreas Naturales Protegidas (ANP) han reportado densidades mayores. Vega-Flores y Farías González (2021) estimaron 15.4 individuos 100 km-2 en Oaxaca, mientras que López-González et al. (2015) reportaron 17 individuos 100 km-2 en el estado de Puebla. De manera similar, en Estados Unidos, Heilbrun et al. (2003) registraron 43 individuos 100 km-2 en un refugio de vida silvestre, y Larrucea et al. (2007) documentaron 39 individuos 100 km-2 en una reserva en California. Estas diferencias pueden atribuirse a que en estas áreas destinadas a la conservación, la superficie de cobertura vegetal es mayor, proporcionando condiciones favorables para una mayor densidad del lince rojo (Morales-Delgado et al. 2021).
Los resultados del modelo óptimo (Tabla 3) muestran que la probabilidad de ocupación del lince rojo es mayor en sitios con alta cobertura arbórea y a mayor distancia de los aerogeneradores. La constante presencia de personal para mantenimiento podría explicar la tendencia de la especie a evitar dichas zonas, ya que se ha documentado que evita activamente el contacto humano (Monroy-Vilchis y Velázquez 2002, Donovan et al. 2011, Litvaitis et al. 2015). Este resultado coincide con el de Espinosa-Flores et al. (2020), que obtuvieron mayor probabilidad de ocupación del L. rufus a mayor cobertura arbórea y mayor distancia de los poblados humanos.
Tabla 3 Modelo de ocupación para Lynx rufus en el sitio de estudio. Se incluyen el Criterio de Información Akaike (AIC), delta del Criterio de Información de Akaike modificado para muestras pequeñas (ΔAIC), Peso del Modelo (AICwgt) y Número de Parámetros (No. Par). Veg = cobertura vegetal del MET, past = pastizal, temp = temperatura promedio diaria, abun = abundancia, cob = cobertura, dist = distancia al aerogenerador más cercano, div = diversidad vegetal y alt = altura promedio.
| Modelo | AIC | ΔAIC | AICwgt | No. Par. |
|---|---|---|---|---|
| ψ (cob*dist) p (veg*past*temp*cob*abun*div*alt) | 996.19 | 0.00 | 0.3204 | 11 |
| ψ (cob) p (veg*past*temp*cob*abun*div*alt) | 997.30 | 1.11 | 0.1840 | 10 |
| ψ (cob*dist*alt) p (veg*past*temp*cob*abun*div*alt) | 997.68 | 1.49 | 0.1521 | 12 |
La probabilidad de detección se asoció positivamente con áreas de pastizal y con una mayor cobertura arbórea, mientras que las superficies agrícolas no mostraron relación significativa. Este patrón concuerda con estudios previos que indican que el lince rojo evita el uso de cultivos agrícolas, prefiriendo el uso de pastizales, donde la disponibilidad de presas suele ser mayor (Monroy-Vilchis y Velázquez 2002, Flores-Morales et al. 2018).
Referente al uso del hábitat, las estaciones 17 y 18 (Figura 1) presentaron el mayor número de registros, incluyendo la observación de una hembra con crías realizando actividades de depredación en distintos días. El sitio 18, caracterizado por su alta cobertura vegetal y diversidad florística, parece ofrecer condiciones adecuadas para la caza y el refugio del lince rojo. Cercano al sitio de estudio, Hanley et al. (2025) encontraron una relación positiva entre la cobertura arbórea y el número de linces identificados al sur de Texas. Contrario a lo anterior, la estación 25 (Figura 1) presentó bajos valores de abundancia, diversidad y cobertura vegetal; sin embargo, registró 23 detecciones de lince rojo, el tercer valor más alto del estudio. Mientras que Abouelezz et al. (2018) sugieren que el lince rojo puede utilizar rutas con alto desarrollo antropogénico para llegar a cubiertas forestales densas, terrenos rocosos o zonas con alta densidad de arroyos, es decir, su hábitat de preferencia. Por otro lado Young et al. (2019) demostraron que los linces en Texas pueden encontrarse en áreas urbanizadas siempre que existan elementos como vegetación remanente o cuerpos de agua. De manera similar, mientras que Vanbianchy et al. (2018) reportaron que Lynx canadensis se desplaza entre áreas perturbadas con baja cobertura forestal, lo que resalta la importancia de considerar estos fragmentos como corredores funcionales en programas de conservación y manejo. Estos resultados subrayan la importancia de conservar la integridad estructural y funcional de los fragmentos de matorral espinoso tamaulipeco, que actúan como corredores biológicos en paisajes fragmentados.
Conclusiones
La densidad poblacional estimada de Lynx rufus es baja en comparación con la reportada en Áreas Naturales Protegidas, respaldando que la continuidad y calidad del hábitat son factores determinantes para el mantenimiento de poblaciones viables en zonas antropizadas. Confirmando la segunda hipótesis, el modelo de ocupación demostró que la cobertura arbórea es la variable con mayor efecto sobre la probabilidad de ocupación, y la segunda en la probabilidad de detección. La preferencia del lince rojo por áreas con vegetación densa y la tendencia a evitar zonas agrícolas o con presencia humana indican la necesidad de parches de vegetación que proporcionen refugio y disponibilidad de presas. La detección de individuos en sitios con menor cobertura vegetal, pero con conectividad hacia superficies de mayor cobertura vegetal o con presencia de arroyos, evidencia la capacidad del lince rojo para desplazarse en paisajes fragmentados y utilizar corredores entre parches de hábitat. Esto subraya la importancia ecológica de los fragmentos y áreas de vegetación remanente que funjan como corredores biológicos.

