











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El fenómeno del cambio climático (CC) afecta a los ecosistemas terrestres de manera diferenciada en distintas regiones del mundo, esto, como resultado de las intensas interacciones entre los factores ambientales y socioeconómicos en el uso de la tierra, influyendo así en la vulnerabilidad y resiliencia de los sistemas productivos (Franzluebbers y Hendrickson 2024). Ante este hecho, surge la necesidad de modelar los probables impactos del CC en los sistemas de producción agrícolas, pecuarios y forestales, de tal manera que sea posible establecer medidas de mitigación y adaptación ante los cambios esperados en dichos sistemas (Calvas-Serrano et al. 2024).
Existen diferentes metodologías para la identificación de sitios, en los cuales, organismos de flora y fauna podrían establecerse y desarrollarse con éxito, es decir, métodos para modelar la posible distribución geográfica y ecológica de las especies (Antúnez et al. 2017, Gutiérrez y Trejo 2022). Estos modelos han abordado cuestionamientos a diferentes escalas, desde el efecto del CC en la distribución del hábitat (Romero-Sánchez et al. 2017), hasta la delimitación de especies y la evolución de sus nichos (Sillero et al. 2021), así como también, procesos ecológicos como los cambios de rango (especiación, expansión, invasión, extinción) e interacciones (competencia, hibridación, evolución morfológica) (González-Cubas et al. 2020), mediante los llamados Modelos de Nicho Ecológico (MNE) reportados por Soberón et al. (2017) y Jiménez y Méndez (2021).
El nicho ecológico, es aquella región del espacio ambiental donde puede existir una especie, dependiendo de variables como la dispersión e interacciones bióticas, de tal manera que pueden determinar el área donde existe una especie (nicho realizado) y el área con las condiciones abióticas aceptables para la especie (nicho fundamental) (Bradie y Leung 2017, Gutiérrez et al. 2018). En otras palabras, los MNE asocian el área abióticamente adecuada de una especie con condiciones ambientales disponibles en las localidades donde se presenta la especie (Manzanilla-Quijada et al. 2020). Por tanto, es posible obtener en un territorio geográfico, la distribución espacial de los entornos más adecuados para la presencia de una especie (Palacios et al. 2021). De tal manera que los MNE son una representación matemática de cómo las distribuciones de los organismos están limitadas por el ambiente, correlacionando puntos de presencia con variables ambientales, brindando además la posibilidad de crear mapas mediante los cuales es posible representar distribuciones potenciales o contemporáneas de una especie (Maciel-Mata et al. 2015, Soberón et al. 2017, CONABIO 2023a).
Actualmente hay una variedad de softwares que correlacionan la presencia de las especies con coberturas ambientales para estimar mapas que representan MNE y/o MDE (Modelos de Distribución de Especies) como los reportados por Soberón et al. (2017), y Melo-Merino et al. (2020). Estos programas se basan en inteligencia artificial y en algoritmos matemáticos; para obtener información confiable independientemente si el número de registro de datos usados son abundantes o limitados (Flores-Tolentino et al. 2019, Pozo-Gómez et al. 2022).
Alrededor del mundo y particularmente en México, existe una gran diversidad de especies arbóreas y arbustivas con diferentes usos en la actividad agropecuaria, y son consideradas como un recurso multipropósito importante en los agroecosistemas pecuarios, siendo el uso forrajero uno de los principales, el cual es aprovechado para la alimentación de rumiantes (Guerrero-Rodríguez et al. 2021). En este sentido, en el Noreste de México (NM), destacan dos especies: Leucaena leucocephala (Lam.) de Wit., y Acacia farnesiana (L.) Willd. (Martínez-Hernández et al. 2019, Molina et al. 2023). La primera por su aptitud para ser incorporada en sistemas agroforestales y distribución amplia en zonas tropicales, subtropicales, áridas y semiáridas (Martínez-Hernández et al. 2019); la segunda está ampliamente distribuida en el Matorral Espinoso Tamaulipeco (MET), formando parte de la vegetación ruderal y por su capacidad para adaptarse a condiciones medioambientales adversas (Molina et al. 2023). Por lo anterior, el objetivo de esta investigación fue modelar la distribución potencial de Leucaena leucocephala y Acacia farnesiana para el Noreste de México.
Materiales y métodos
Área de Estudio
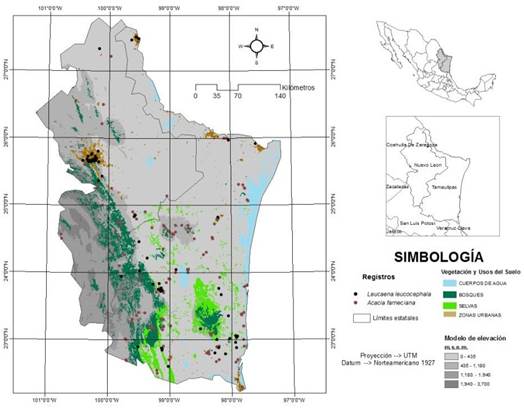
El área de estudio comprendió la región Noreste de México (NM) en la cual se encuentran amplias zonas dominadas por el MET, este ecosistema se distribuye desde el sur de Texas (Estados Unidos), hasta Coahuila, Nuevo León y Tamaulipas, con una superficie continua de 125 000 y hasta los 200 000 km2 (Figura 1), presentando también áreas con condiciones semiáridas y subhúmedas en esta región (Patiño-Flores et al. 2022). Para el alcance de esta investigación solo se consideró como NM a los Estados de Nuevo León y Tamaulipas, los cuales comparten la Gran Llanura de Norteamérica y la Llanura Costera del Golfo Norte; además presentan similitudes en fuentes productoras de biomasa, principalmente en especies del estrato herbáceo (Guzmán-Lucio et al. 2016). Ambos Estados abarcan de manera conjunta una superficie de 14.4 millones de hectáreas, de las cuales, Tamaulipas y Nuevo León destinan 4.9 y 3.0 millones de hectáreas a las actividades agropecuarias, 63 y 47% del total Estatal, respectivamente (INEGI 2022a). En el NM predominan los climas de tipo cálido subhúmedo A(w), semicálido subhúmedos AC(w,x), semiseco muy cálido y cálido BS1(h´), seco muy cálido y cálido BS0(h´)w(w) y seco semicálido BSh (INEGI 2022b).
Descripción de las especies modeladas
Las especies modeladas fueron Acacia farnesiana (L.) Willd. y Leucaena leucocephala (Lam.) de Wit.; las cuales sobresalen de sus requerimientos ecológicos por su capacidad para tolerar la sequía, incluso durante su establecimiento y pueden soportar entre 6 y 8 meses de periodo seco (Tabla 1).
Tabla 1 Requerimientos ecológicos para Acacia farnesiana (L.) Willd., y Leucaena leucocephala (Lam.) de Wit.
| Especie | Variables Biofísicas* | Referencia | |||||
|---|---|---|---|---|---|---|---|
| Ppma (mm) | Tma (°C) | Tmmf (°C) | Tmmc (°C) | Psa (meses) | Alt (msnm) | ||
| A. farnesiana | 1 200 - 2 500 | 22 - 24 | > 18 | 26 - 28 | 0 - 6 | - | Gómez et al. (2011) |
| 250 - 5 000 | 20 - 32 | 10 - 20 | 20 - 42 | - | 0 - 2 100 | FAO (2024) | |
| 350 - 2 300 | 22 - 30 | - | - | 4 - 6 | 0 - 900 | CONABIO (2023b) | |
| L. leucocephala | 100 - 1 400 | 16 - 26 | 14 - 20 | 20 - 26 | 2 - 8 | Gómez et al. (2011) | |
| 400 - 1 400 | 20 - 32 | 7 - 20 | 32 - 42 | - | 1 300 | FAO (2024) | |
| 400 - 1 500 | 24 | 13 - 15 | > 30 | - | 0 - 1 500 | CONABIO (2023b) | |
*Ppma: Precipitación media anual; Tma: Temperatura media anual; Tmmf: Temperatura del mes más frío; Tmmc: Temperatura del mes más caliente; Psa: Periodo seco anual; Alt: Altitud.
Modelación
Los modelos de nicho ecológico para A. farnesiana y L. leucocephala para el Noreste de México, se desarrollaron con el algoritmo de máxima entropía del software Maxent Ver. 3.3.3k (Phillips et al. 2006). De los múltiples programas disponibles, destaca Maxent, el cual permite proyectar patrones de distribuciones geográficas mediante el empleo de MNE, utilizando registros observados de presencias puntuales (Pérez-García y Liria 2013, Menezes 2023). Una de las características importantes de Maxent, es que se pueden hacer predicciones a partir de información incompleta, estimando la distribución más uniforme para el área estudiada; además es considerado un algoritmo confiable por su buen desempeño (Charre-Medellín et al. 2019). Para la construcción de los modelos se utilizaron 19 variables bioclimáticas, las cuales se obtuvieron de la base de datos WorldClim 1.3 (Hijmans et al. 2005). Los registros de presencia de las especies se obtuvieron de las bases de datos Global Biodiversity Information Facility (GBIF), de la Comisión Nacional sobre la Biodiversidad (CONABIO) y se completaron con registros de campo. La base de datos final de registros de presencia fue depurada con el visualizador de Google EarthTM (Pérez-García y Liria 2013) para descartar datos erróneos, localidades inciertas y duplicación de datos. Se utilizaron 143 registros de presencia de A. farnesiana, obtenidos de 8 conjuntos de datos publicados en la base de datos GBIF (Figura 1). Las fuentes de información de la base de datos GBIF son: Catálogo electrónico de especímenes depositados en el Herbario de la Universidad de Texas en Austin, Fase IV; Inventario florístico de la Sierra de San Carlos, Tamaulipas; Lista florística preliminar de Tamaulipas; Tropicos Specimen Data; The New York Botanical Garden Herbarium; Análisis de viabilidad para poblaciones de la cactácea amenazada Ariocarpus trigonus; Actualización de las bases de datos del Herbario XAL. Fase III; Etapa final de la captura y catalogación del Herbario del Instituto de Ecología, AC, Centro Regional del Bajío; iNaturalist Research-grade Observations; Determinación del estado actual de cinco especies de cactáceas amenazadas del estado de Tamaulipas, México. Para el caso de Leucaena leucocephala, se obtuvieron 82 registros (Figura 1). Las fuentes de información para la obtención de los registros de presencia fueron las bases de datos de la CONABIO (Colecciones Enciclovida y Naturalista), Herbario Nacional (MEXU) del Instituto Nacional de Biología de la UNAM y Herbario del Instituto Nacional de Ecología (INECOL), además de registros generados en salidas de campo.
Análisis estadístico
Para determinar la validez de los modelos, se utilizó el valor AUC (Área Bajo la Curva, por sus siglas en inglés) del análisis de la curva operacional ROC (Receiver Operating Characteristic, por sus siglas en inglés) y ROC parcial, mediante el programa Tool for Partial-ROC (Narayani 2008), utilizando un 50% de los puntos de evaluación independientes re-muestreados en 1 000 réplicas (bootstrap) y fijando un error de omisión no mayor al 5% (1-omission threshold > 0.95) al modelo. Además, se utilizó el método Jackknife, instrumentado en Maxent, para estimar la contribución relativa de cada variable climática.
Resultados
Los modelos de nicho ecológico muestran una probabilidad de idoneidad desde 31 al 95% para A. farnesiana y desde 30 a 92% para L. leucocephala (Figura 2). Los resultados de la modelación sugieren que existen condiciones bioclimáticas adecuadas en amplias zonas geográficas del Noreste de México para la presencia de las especies. Representando un área mayor de idoneidad para A. farnesiana, en el estado de Tamaulipas, mientras que para L. leucocephala, el área de idoneidad es más reducida en todo el Noreste, destacando zonas más extensas para el estado de Tamaulipas con relación a Nuevo León.
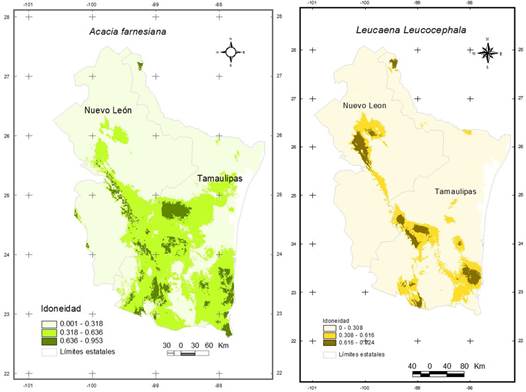
Figura 2 Modelo de Nicho Ecológico de Acacia farnesiana (L.) Willd., y Leucaena leucocephala (Lam.) de Wit., en el Noreste de México.
El análisis del Área Bajo la Curva de la función ROC, muestran resultados de 0.869 para L. leucocephala y 0.787 para A. farnesiana (Figura 3), por lo cual se consideró como un desempeño estadísticamente confiable.
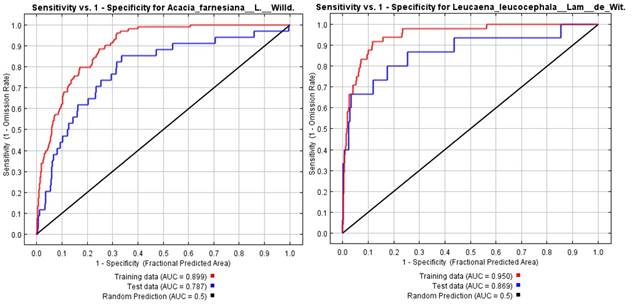
Figura 3 Área bajo la curva (AUC) de Acacia farnesiana (L.) Willd., y Leucaena leucocephala (Lam.) de Wit.
El método Jackknife instrumentado en el programa Maxent, mostró que las variables bioclimáticas con mayor contribución para la probabilidad de presencia en el MNE para L. leucocephala fueron: el rango de temperatura anual, precipitación del mes más húmedo, rango medio diurno/rango de temperatura anual, precipitación del trimestre más frío y temperatura media del trimestre más cálido. Mientras que para A. farnesiana fueron temperatura estacional, precipitación del trimestre más húmedo y precipitación del trimestre más cálido (Tabla 2).
Tabla 2 Contribución de las variables bioclimáticas en la probabilidad del modelos de nicho ecológico para Acacia farnesiana (L.) Willd., y Leucaena leucocephala (Lam.) de Wit.
| Variable | Código | Contribución (%) | |
|---|---|---|---|
| A. farnesiana | L. leucocephala | ||
| Rango medio diurno/Rango de Temperatura Anual | BIO 3 | 13.2 | |
| Temperatura Estacional | BIO 4 | 28.2 | - |
| Rango de Temperatura Anual | BIO 7 | - | 21.1 |
| Temperatura media del trimestre más cálido | BIO 10 | - | 10.6 |
| Precipitación del mes más húmedo | BIO 13 | - | 15.2 |
| Precipitación del trimestre más húmedo | BIO 16 | 18.6 | - |
| Precipitación del trimestre más cálido | BIO 18 | 10.3 | - |
| Precipitación del trimestre más frío | BIO 19 | - | 11.6 |
Discusión
Los MNE ofrecen resultados normalmente muy diferentes entre sí, por lo que la interpretación de cómo juzgar los modelos desde un punto de vista estadístico, es de gran relevancia (Melo-Merino et al. 2020). La validación se refiere a la evaluación de la exactitud en las predicciones de un modelo de distribución de especies (Sillero et al. 2021). El método del valor AUC del análisis ROC ha sido aplicado extensamente con tal propósito (Menezes 2023). Este método se ha convertido en una herramienta dominante para evaluar la precisión de los modelos que predicen la distribución de especies (Melo-Merino et al. 2020, Brown y Griscom 2022). En términos comparativos, el estadístico derivado del análisis de área bajo la curva ROC y que le da soporte estadístico, es equivalente al estadístico U de Mann-Whitney (Sillero et al. 2021).
En su interpretación el AUC, varía de 0 a 1 y representa una medida del desempeño general del modelo, la cual permite discriminar áreas adecuadas contra áreas no adecuadas (Anderson y González 2011). Los valores de AUC pasan desde 0.5 para los modelos sin capacidad predictiva hasta 1.0 para los modelos que brindan predicciones más acertadas (Brown y Griscom 2022). En otras palabras, un valor de 1.0 indica que todos los casos se han clasificado correctamente y uno de 0.5 que el modelo no es diferente de clasificar los casos al azar; valores menores a 0.5 indican que el modelo es realmente desfavorable, ya que clasifica erróneamente más casos que el azar (Sillero et al. 2021).
Es conveniente hacer mención que, el análisis ROC es sujeto de cuestionamientos cuando se utilizan datos de solo presencia, ya que, sobre la base teórica, este análisis también requiere datos de ausencias. Es decir, las predicciones espaciales pueden presentar errores de omisión (falsos negativos, dejando fuera el área de distribución conocida) y errores de comisión (falsos positivos, incluidas áreas inadecuadas en la predicción). Ante esto, la modificación del ROC, para utilizar el enfoque de ROC parcial, proporciona una base más estable para la evaluación de predicciones de MNE y corregir dichos errores (Sillero et al. 2021). Al emplear el método de Jackknife, se encontró que la probabilidad de la presencia de L. leucocephala y A. farnesiana en el MNE, estuvo asociada a las variables bioclimáticas. Este comportamiento es similar a los resultados reportados por Antúnez et al. (2017), quienes señalan que las variables bioclimáticas de temperaturas (mínimas y máximas) y precipitación tienen una contribución significativa en la distribución de especies arbóreas.
El MNE mostró una reducción en el área de idoneidad para el Noreste de México en el caso de L. leucocephala, esto es debido a que las condiciones biofísicas en el Noreste de México no son del todo propicias para esta especie, en esta región del país, donde además predomina el MET como lo mencionan Molina et al. (2023), FAO (2024), CONABIO (2023b) y Gómez et al. (2011). Sin embargo, el MNE muestra que destacan zonas más extensas para esta misma especie para Tamaulipas con relación a Nuevo León. Esto puede ser posible a la capacidad de L. leucocephala, para desarrollarse y prosperar con éxito en amplias zonas tropicales, como lo indican Martínez-Hernández et al. (2019).
El resultado de la modelación muestra que son estadísticamente confiable mediante el análisis del Área Bajo la Curva de la función ROC, lo cual le da validez al modelo obtenido para L. leucocephala y A. farnesiana, como está indicado por Sillero et al. (2021). Los resultados estadísticos de 0.869 para L. leucocephala y 0.787 para A. farnesiana, representan la probabilidad de encontrar más áreas de idoneidad entre las especies aquí modeladas, específicamente con relación a los estados de Tamaulipas y Nuevo León en el Noreste de México, este desempeño es semejante a lo que indican Anderson y González (2011) y Sillero et al. (2021), con relación a la capacidad del AUC para discriminar áreas adecuadas contra áreas no adecuadas.
Las especies modeladas pueden tener un impacto positivo en el medio ambiente, principalmente por los atributos que distinguen a la familia de las leguminosas (fabaceae) como: la fijación simbiótica de nitrógeno, valor nutritivo alto, sistema radicular pivotante profundo, amplia diversidad taxonómica-genética y brindan importantes servicios ambientales (Apan-Salcedo et al. 2021). Estas características les confieren a las leguminosas una ventaja ecológica significativa sobre otras familias de plantas en los sistemas agropecuarios (Schultze-kraft et al. 2018).
Conclusiones
Existen condiciones biogeográficas adecuadas, de acuerdo a la probabilidad de idoneidad de presencia de las especies de leguminosas arbustivas Leucaena leucocephala y Acacia farnesiana en el Noreste de México, por lo que podría ser factible su aprovechamiento en los sistemas de ganadería extensiva prevalecientes en esta región, propiciando una reconversión productiva de manera sustentable, lo que permitiría aminorar los impactos poco favorables de esta actividad productiva en los agroecosistemas ganaderos y el ambiente

