











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
México cuenta con aproximadamente 138 millones de hectáreas de vegetación, representando el 70% de su territorio; de esta superficie, el 17% corresponde a bosques de clima templados (CONAFOR 2014). La función ecológica de este tipo de vegetación contribuye a la regulación del clima al actuar como sumideros de gases de efecto invernadero GEI (FAO 2018). La excesiva acumulación de estos gases en la atmósfera está provocando cambios en el clima, prueba de ello, es el aumento de temperatura de 0.87°C registrado en el período de 2006 a 2015 (Allen et al. 2018). Pero en 2017 las concentraciones de GEI alcanzaron niveles globales por encima de las 405 ppm de CO2 (WMO 2018), de continuar esta tendencia, es probable alcanzar un aumento de temperatura media anual global de 1.5°C para 2030 a 2040 (IPCC 2018, Allen et al. 2018, Hansen et al. 2019). Este aumento de temperatura ira asociado de una disminución de las precipitaciones de hasta el 30% en muchas regiones del mundo (IPCC 2013). Para México se estima una reducción en precipitación del 9% para el 2060 (Sáenz-Romero et al. 2010). La alteración de la composición del ecosistema por la extracción de especies, la conversión de uso de suelo, y los incendios forestales, entre otros, provocan debilitamiento y estrés en los árboles, lo que reduce su eficiencia reguladora (Salinas et al. 2010) y favorece la incidencia de plagas forestales (Gómez-Pineda et al. 2022). Pese a todo esto, el manejo eficaz de los ecosistemas en relación con el cambio climático representa retos para los administradores, manejadores forestales y ecologistas.
En particular, destacan las especies de los géneros Pinus y Quercus, especies de importancia ecológica y económica, que se distribuyen secuencialmente desde los altos y bajas altitudes de las provincias fisiográficas de México: Abies vejarii (2 000-3 300 msnm, Farjon y Styles 1997) catalogado como amenazada (SEMARNAT 2010), Pinus pseudostrobus (1 900-3 000 msnm, Farjon y Styles 1997) es de gran importancia económica, Pinus teocote (1 500-3 000 msnm, Farjon y Styles 1997), Quercus canbyi (650-1 200 msnm, Valencia 2004), Quercus laeta (1 900-2 270 msnm, Valencia 2004), Quercus mexicana (2 230-2 700 msnm, Valencia 2004), Quercus polymorpha (1 900-2 000 msnm, Valencia 2004), Quercus rugosa (1 800-2 900 msnm, Valencia 2004), Arbutus xalapensis (2 400-3 000 msnm, Farjon y Styles 1997), Juniperus flaccida (1 600-2 000 msnm, Farjon y Styles 1997).
A pesar de la abundante diversidad de especies de plantas en México, no significa que estén exentos de alguna amenaza, uno de los más destacados es el impacto del cambio climático que se ha visto reflejado en la alteración de la distribución, tamaño, estructura y abundancia de las especies (Parmesan 2006) o en la desaparición directa de algunas especies y poblaciones (Thomas et al. 2004). Esto será más drástico para las especies de ecosistemas geográficamente confinados, sobre todo de montaña, y por ello varias especies tenderán a desplazar su distribución a latitudes y altitudes distintas de las actuales (Guitérrez y Trejo 2014). Esto es especialmente cierto para las poblaciones de especies arbóreas situadas en los límites xéricos de especies que se encuentran en el límite altitudinal inferior o sur (en el hemisferio norte) de su distribución natural (Mátyás et al. 2010). Sin embargo, las modificaciones en el clima podrían superar su capacidad para migrar o sobrevivir en las nuevas condiciones ambientales (Gavilán 2008, Sáenz-Romero et al. 2016). Por esta razón, el cambio climático presenta retos importantes para los manejadores de los bosques (Gallardo-Salazar et al. 2023). Debido a que las tasas de supervivencia de los árboles disminuyen (Sáenz-Romero et al. 2017).
Existen modelos para predecir la distribución de especies (Elith et al. 2006), y son representaciones cartográficas de la capacidad de una especie para ocupar un espacio geográfico, determinado en función de variables continuas o categóricas (Guisan y Zimmermann 2000) también llamadas bioclimáticas. Las más utilizadas son las variables climáticas (Felicísimo et al. 2012). MaxEnt es un algoritmo basado en el enfoque estadístico de máxima entropía adaptado para la construcción y predicción de modelos de distribución potencial de organismos vivos (especie de interés) utilizando datos de presencia (Phillips et al. 2006), esta técnica se aplica para priorizar áreas de conservación biológica (Ávila et al. 2014, Martínez-Sifuentes et al. 2020), evaluar los efectos del cambio climático (Manzanilla-Quijada et al. 2020a, Manzanilla-Quijada et al. 2024a), estimar y delimitar áreas potenciales para la producción de semillas (Manzanilla et al. 2019, Manzanilla-Quijada et al. 2020b) y modelar la distribución potencial de especies de pinos (Cruz-Cárdenas et al. 2016, Ramos-Dorantes et al. 2017).
La hipótesis de este estudio es que ante escenarios de cambio climático las especies de bosques templados de los géneros Pinus y Quercus presentaran modificaciones en sus áreas de distribución potencial natural. Para responder dicha hipótesis se planteó el siguiente objetivo, proyectar a través de modelos la distribución potencial actual y bajo escenarios de cambio climático de 10 especies arbóreas de clima templado del estado de Nuevo León, en México y definir áreas de conservación.
Materiales y métodos
Área de estudio y especies estudiadas
El estudio se realizó en el estado de Nuevo León, el cual se localiza en la zona noroeste de la República Mexicana, abarcando una superficie de 65 057.17 km2, representando el 3.3% del territorio nacional. Entre las coordenadas 27° 47’ y 23° 10’ latitud Norte y 98° 24’ y 101° 12’ latitud oeste (Figura 1).
Las especies de estudio (Tabla 1), además de su importancia ecológica y económica, también son de importancia forestal; estas especies se encuentran ubican en bosques templados del sistema montañoso de la Sierra Madre Oriental (SMO) principalmente, en altitudes que alcanzan hasta los 3 000 msnm, temperatura media anual de 20°C, temperatura máxima promedio de 32°C (mayo a agosto) y temperatura mínima promedio de 5°C (enero), con precipitación media estatal de 650 mm anuales (agosto y septiembre) (INEGI 2001). En el caso específico de A. vejarii es una especie que se encuentra catalogado como amenazada (SEMARNAT 2010), por lo que la hace más sensible a los efectos del cambio climático.
Origen y recopilación de los datos de presencia
Los registros de cada una de las especies, se tomaron directamente en campo (Sistema montañoso de la SMO) utilizando un GPS, los cuales fueron complementados con puntos de colectas disponibles de diversas fuentes (Tabla 2). La primera fue la base de datos del Sistema Nacional de Información sobre Biodiversidad; posteriormente se consultó el repositorio digital del Herbario Nacional de México (MEXU-UNIBIO, UNAM); seguido de la Red internacional e Infraestructura Global de Información de Biodiversidad (GBIF 2020); y de la Red social de ciencia (NATURALISTA) de la CONABIO. Todos los registros dudosos, tanto taxonómica como geográficamente, fueron cuidadosamente evaluados para su inclusión o eliminación en los análisis. Los datos obtenidos fueron depurados a través de la plataforma Niche ToolBox (Osorio-Olvera et al. 2020), excluyendo las coordenadas con información ambigua, duplicadas, mal georreferenciados y dentro de áreas urbanas, dejando una distancia de separación de 1 km2 (aprox. ~30 segundos de arco) entre cada uno de los registros (Pearson et al. 2007) (Tabla 2).
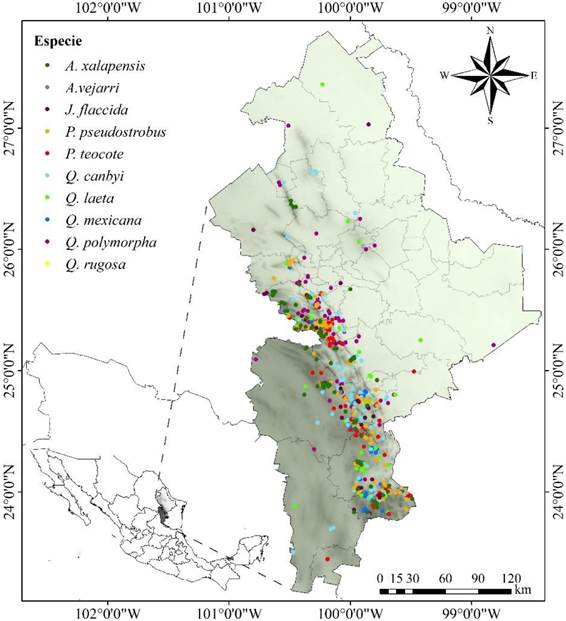
Figura 1 Área de estudio y registro de la presencia de especies arbóreas distribuidas en el estado en Nuevo León.
Tabla 1 Información biológica de las especies en estudio.
| Especie | Descripción | Altitud (msnm) | Autor |
|---|---|---|---|
| Abies vejarii Martínez | Árbol perenne de tamaño medio, alcanza los 35-40 m de altura. Las hojas son de aguja 1-2.5 cm de longitud y 1.3-2 mm de ancho, de color verde-gris con estomas arriba y dos bandas de estomas debajo. Las piñas son púrpura que al madurar tornan marrón de 6-15 cm de longitud y 4-6 cm de ancho, con unos 150-200 frutos, cada uno con una bráctea de 3-8 mm sobre el cono cerrado y dos semillas ladas; se desintegran cuando maduran para lanzar las semillas. | 2 000 - 3 300 | Farjon y Styles 1997 |
| Pinus pseudostrobus Lindl. | Árbol con altura promedio de 20 a 40 metros, (incluso se habla de ejemplares de hasta 50 metros) y un diámetro de 40 a 80 cm. Las ramas generalmente tienen ubicación horizontal, yemas oblongas-cónicas de color anaranjado. Conos de ovoides a cilíndricos de color café claro, de 10 a 15 cm de largo y curveados, estos se agrupan entre dos y tres en las puntas de la rama. | 1 900 - 3 000 | Farjon y Styles 1997 |
| Pinus teocote Schiede ex Schltdl. & Cham | Árbol perennifolio que alcanza alturas de hasta 20 m. La corteza es fisurada de color café grisáceo. Las hojas de 10 a 15 cm y en fascículos de tres, rígidas de color verde oscuro y brillante. Los frutos son conos ovoides de 3.5 a 6.5 cm de largo, de color café claro a café rojizo. | 1 500 - 3 000 | Farjon y Styles 1997 |
| Quercus canbyi Trel. | Árbol que alcanza un tamaño de entre los 4 a los 15 metros de altura, y el tronco entre 20 a 50 cm de diámetro. Las ramas son largas y flexibles, algo caídas. La corteza es de color marrón verdoso, suave, convirtiéndose en gris oscuro, verrugoso, surcado. Las hojas miden 7,5-10 x 2-3 cm, semiperennifolias o últimamente caducifolias, gruesas, correosas, lanceoladas a estrechamente elípticas. | 650 - 1 200 | Valencia 2004 |
| Quercus laeta Liebm. | Árbol de hasta 10 metros de altura con un tronco de hasta 40 centímetros (16 pulgadas) de diámetro. Las hojas son de color verde oscuro, gruesas y coriáceas, de hasta 40 cm de largo con algunos dientes poco profundos a lo largo de los bordes. | 650 - 1 200 | Valencia 2004 |
| Quercus mexicana Bonpl. | Árbol de hoja caduca que crece hasta 15 metros de altura con corteza gris. Las hojas son gruesas y coriáceas, de hasta 12 centímetros de largo, oblongo o con forma de lanza, sin dientes ni lóbulos. | 2 230 - 2 700 | Valencia 2004 |
| Quercus polymorpha Schltdl. & Cham. | Árbol caducifolio, con una altura que varía de 12 a 20 metros de altura, con copa ancha y redondeada. sus hojas son simples, alternas, ovadas, de base redonda, de 4 a 15 centímetros de largo y 3 a 8 cm. de ancho. | 1 900 - 2 000 | Valencia 2004 |
| Quercus rugosa Née | Árbol o arbusto bajo (3-8 m), aunque excepcionalmente alcanza los 30 m de altura. Corteza oscura con profundas fisuras que forman escamas; sus hojas son elíptico-obovadas de margen entero o a veces aserrado, muy rígidas al madurar y notablemente cóncavas por el envés. El fruto es una bellota ovoide pequeña (15-25 mm de largo), angosta y puntiaguda. Crece preferentemente en laderas y barrancas con clima templado subhúmedo. | 1 800 - 2 900 | Valencia 2004 |
| Arbutus xalapensis Kunth | Arbusto o árbol mediano de la familia Ericaceae; perennifolio, que alcanza los 3 m de altura. La corteza se desprende en capas delgadas. Sus hojas delgadas y resistentes de 2-18 cm son brillantes por encima y opacas por debajo. Las flores van de 7-9 mm de largo, son amarillas a blancas. El fruto es una baya roja globosa, menor a 1 cm de diámetro. | 2 400 - 3 000 | Farjon y Styles 1997 |
| Juniperus flaccida Schltdl. | Árbol o arbusto pequeños de 5 hasta 12 metros de altura, cuyo tronco se ramifica frecuentemente a una altura de 1 a 2 m y con un diámetro de 25 a 50 cm. Copa de forma globosa de hasta 10 m de ancho. Corteza, profundamente fisurada, con placas fibrosas entrelazada de color grisáceas en su exterior y café rojizas en su interior, dividida en tiras longitudinales. Ramas, extendidas formando una copa redondeada e irregular, las ramas que son secundarias por lo general péndulas y fláccidas, con un sistema terminal de ramas foliosas, dispuestas en dos filas verticales opuestas. Hojas, opuestas, con los ápices pegados al tallo y sin punta, el margen irregularmente dentado. | 1 600 - 2 000 | Farjon y Styles 1997 |
Obtención y selección de las variables climáticas actuales (período 1910-2009) y futuras (Década 2050).
Se obtuvieron 19 variables climáticas en formato ráster a una resolución de ~30 segundos de arco equivalente a 1 km2 por pixel aproximadamente (Tabla 3), información derivada de la interpolación de los registros de las estaciones climáticas mensuales de precipitación, temperatura mínima y máxima, para México del período comprendido entre los años 1910-2009, disponibles en la plataforma del Centro de Recursos Idrisi-México de la Universidad Autónoma del Estado de México, CRI-UAE-Méx (Cuervo-Robayo et al. 2014). Las variables climáticas a futuro se obtuvieron a partir del Modelo de Circulación Global (MCG) del Laboratorio de Dinámica de Fluidos Geofísicos con el componente de Dinámica de la Capa Oceánica Generalizada (GFDL_ESM2G). Además, el componente atmosférico de los ESM incluye características físicas como aerosoles (tanto naturales como antropogénicos), física de nubes y precipitación. El MCG se descargó bajo dos escenarios de trayectorias de concentración representativas de gases de efecto invernadero (RCP por sus siglas en inglés) de RCP4.5 y 8.5 para el 2050; donde el primer escenario representa un forzamiento radioactivo de condiciones actuales similares a las proyectadas a futuro considerado como conservador (Fernández et al. 2015), el segundo escenario proyecta condiciones donde las emisiones de gases de efecto invernadero se encuentran en constante aumento considerado como pesimista (Van-Vuuren et al. 2011, IPCC 2014). Dichos escenarios fueron seleccionados porque son los que se utilizan para actualizar los estudios de impacto, vulnerabilidad y adaptación en México y Centroamérica (Fernández-Eguiarte et al. 2015).
Tabla 2 Procedencia de ejemplares utilizados en la modelación de la distribución de especies.
| Clave | Nombre científico | Campo | SNIB | GBIF | IBUNAM-MEXU | Naturalista | Total |
|---|---|---|---|---|---|---|---|
| Ave | Abies vejarii Martínez | 11 | 1 | 26 | 38 | ||
| Pps | Pinus pseudostrobus Lindl. | 29 | 36 | 32 | 1 | 34 | 132 |
| Pte | Pinus teocote Schiede ex Schltdl. & Cham | 7 | 30 | 49 | 1 | 4 | 91 |
| Qca | Quercus canbyi Trel. | 5 | 38 | 71 | 3 | 10 | 127 |
| Qla | Quercus laeta Liebm. | 7 | 42 | 51 | 1 | 2 | 103 |
| Qme | Quercus mexicana Bonpl. | 7 | 27 | 40 | 10 | 84 | |
| Qpo | Quercus polymorpha Schltdl. & Cham. | 10 | 45 | 82 | 6 | 5 | 148 |
| Qru | Quercus rugosa Née | 5 | 2 | 2 | 1 | 10 | |
| Axa | Arbutus xalapensis Kunth | 16 | 56 | 1 | 13 | 86 | |
| Jfl | Juniperus flaccida Schltdl. | 20 | 11 | 14 | 45 | ||
| Total, general | 117 | 221 | 420 | 23 | 83 | 864 | |
SNIB: Sistema Nacional de Información sobre Biodiversidad; GBIF: Global Biodiversity Information Facility; IBUNAM-MEXU: Herbario Nacional de México bajo custodia del Instituto de Biología de la UNAM.
Se seleccionaron las variables climáticas que no estuvieran correlacionadas entre sí (Tabla 2). Este procedimiento se realizó a través del análisis de multicolinealidad en el programa R studio v. 4.0.3 (R Core Team 2020), empleando el factor de inflación de la varianza (VIF, por sus siglas en inglés) para seleccionar las variables menos correlacionadas a partir de una matriz con las 19 variables, donde los valores de correlación de Pearson de ≥ -0.85 o +0.85 indican problemas de multicolinealidad entre las variables. Todas las variables fueron convertidas a formato ráster ASCII y ajustadas a una resolución espacial aproximada de 1 km2 (aprox. ~30 segundos de arco) (Tabla 3).
Tabla 3 Variables climáticas empleadas para cada uno de los análisis finales en la modelación de la distribución de las especies en estudio.
| Clave | Variables bioclimáticas Unidad (en paréntesis) | Especie | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ave | Pps | Pte | Qca | Qla | Qme | Qpo | Qru | Axa | Jfl | ||
| Bio6 | Temperatura mínima del mes más frío (°C) | * | * | * | * | * | * | * | * | ||
| Bio7 | Rango de temperatura anual (Bio5 - Bio6) | * | * | * | * | * | * | * | * | * | * |
| Bio8 | Temperatura media del trimestre más húmedo | * | * | * | * | * | * | * | * | ||
| Bio9 | Temperatura media del trimestre más seco | * | * | * | * | * | |||||
| Bio10 | Temperatura media del trimestre > más caliente | * | |||||||||
| Bio11 | Temperatura media del trimestre más frío | * | |||||||||
| Bio12 | Precipitación total anual (mm) | * | * | * | * | * | * | ||||
| Bio13 | Precipitación del mes más húmedo (mm) | * | |||||||||
| Bio14 | Precipitación del mes más seco (mm) | * | * | * | |||||||
| Bio15 | Estacionalidad de la precipitación (coeficiente de variación) (mm) | * | * | * | * | * | * | * | * | * | * |
| Bio16 | Precipitación del trimestre más húmedo (mm) | * | * | * | |||||||
| Bio17 | Precipitación del trimestre más seco (mm) | * | * | * | * | ||||||
| Bio18 | Precipitación del trimestre más caliente (mm) | * | * | * | * | * | * | * | * | ||
| Bio19 | Precipitación del trimestre más frío (mm) | * | * | * | * | * | * | * | |||
| Total | 5 | 7 | 7 | 7 | 9 | 8 | 6 | 9 | 9 | 8 | |
* Selección de variables definitiva (correlación de Pearson de ≥ -0.85 o +0.85).
Área de calibración y delimitación del área accesible (M)
La delimitación del área de calibración, denotada por «M» en el diagrama de «BAM», es fundamental en la modelación de nichos y distribución de especies, así como en la generación y transferencia del modelo (Barve et al. 2011, Soberón et al. 2017) ya que se ha descrito como el espacio donde una especie esta o se asume que podría estar, con base en el conocimiento biológico y su capacidad de dispersión. Para la delimitación del área «M» se empleó el estado de Nuevo León y a partir de esta área se recortaron y ajustaron las variables ambientales a un tamaño de píxel de 1 km2 (aprox. ~30 segundos de arco) a través del programa ArcMap v. 10.5. (ESRI 2016).
Generación y validación de los modelos
Se utilizó el algoritmo de Máxima Entropía (MaxEnt) v. 3.4.3k (Phillips et al. 2006) para construir los modelos de distribución potencial de cada una de las especies. Con 100 réplicas para cada taxa, considerando la regla de aplicación umbral (equal test sensitivity and specificity), un máximo de 1 000 iteraciones, un límite de convergencia de 10-5 (valor por omisión) y un replicado interno por Bootstrap (Phillips et al. 2006). Se omitieron las opciones Extrapolate y Do clamping con la finalidad de evitar extrapolaciones artificiales en los valores extremos de las variables bioclimáticas (Elith et al. 2011). Se consideró el formato de salida del tipo logística de MaxEnt con un umbral de 0 a 1 (0 = no favorable y 1 = totalmente favorable); es decir, mientras el valor se aproxime a 1 las condiciones ambientales serán idóneas para la especie. Se incluyó el 75% de los registros geográficos de la especie para el entrenamiento del modelo y el 25% restante de los datos para la validación de los modelos. Se estimo la contribución relativa de cada variable a través de la prueba de Jackknife (Phillips et al. 2006, Cobos et al. 2019). Para la generación de los modelos de distribución proyectados a futuro del período 2050 se realizaron de acuerdo con los parámetros del modelo de distribución actual, a los cuales se aplicaron los mismos criterios de evaluación y validación efectuado en el modelo actual.
Los modelos se validaron acorde al desempeño y ajuste estadístico en las pruebas de entrenamiento y validación de la AUC (Area Under the Curve) de la prueba ROC-estándar (Receiver Operating Characteristic) (Phillips et al. 2006). Se realizó la prueba de ROC-parcial para subsanar las deficiencias del AUC, este análisis fue efectuado mediante la plataforma Niche ToolBox (Osorio-Olvera et al. 2020), utilizando un 50% de los registros re-muestreados en 1000 réplicas Bootstrap y fijando un error de omisión no mayor a 5% (1-omission threshold > 95). Además, se realizó una prueba de Z, lo que brindó de mayor robustez estadística a los modelos.
Distribución potencial actual y futura
Se generaron mapas de distribución potencial de cada especie a través del programa ArcMap v. 10.5 (ESRI 2016), reclasificando las salidas ASCII de los modelos en tres categorías de probabilidad de idoneidad (nula, media y alta) con intervalos iguales, calculando la superficie total por categoría que ocupa cada una de ellas. Para evaluar el potencial impacto del cambio climático en la distribución futura de las especies arbóreas, se obtuvo el porcentaje de cambio (superficie de distribución) por medio de la siguiente fórmula: % de cambio = [(S1-S0)/S0] *100%, donde S0 es la superficie total que ocupan en el área de estudio de acuerdo con el período de referencia actual (1910-2009) y S1 es la superficie total que se proyecta ocupará en la década centrada en el 2050. Con el fin de visualizar el aumento o reducción de la superficie donde se distribuye las especies dentro de la Sierra Madre Oriental en el estado de Nuevo León.
Áreas aptas para la conservación
La conservación de áreas potenciales de distribución de cada especie, se identificaron mediante la aplicación de un intersecto entre las áreas de distribución actual y las áreas proyectadas a futuro. Se optó por emplear solamente las especies que estimaron mayor superficie reducida en los modelos a futuro respecto a la distribución actual, con la finalidad de identificar zonas mínimas de persistencia de distribución de especies, las cuales pudieran ser utilizadas como áreas de conservación (Manzanilla-Quijada et al. 2020a, Martínez-Sifuentes et al. 2020) las cuales ayudarían a los programas de conservación y restauración.
Resultados
Al final del proceso de depuración se obtuvieron 864 registros únicos, distribuidos entre las 10 taxas, los cuales se encuentran distribuidos dentro de la Sierra Madre Oriental del estado de Nuevo León. La especie con mayor número de registros fueron Q. polymorpha (148), seguida de P. pseudostrobus (132) en contraste con Q. rugosa (10) (Tabla 2).
Validación de los modelos
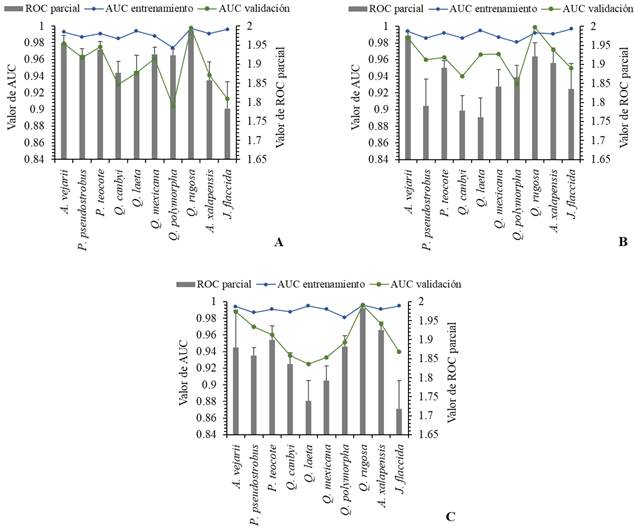
Los valores del área bajo la curva (AUC) de los datos de entrenamiento y validación en el escenario actual y futuro se estimaron por encima de 0.900 para los 10 taxones (Figura 2). Obteniendo un promedio de AUC en los datos de entrenamiento y validación para todas las especies en el escenario actual de 0.990 y 0.951; en el escenario RCP4.5 fue 0.990 y 0.963; y para el escenario RCP8.5 fue 0.991 y 0.957 respectivamente. Mientras que los valores de las pruebas ROC-parciales reportaron valores por arriba de 1.700 (Figura 2), con promedio para el escenario actual de 1.906; para el escenario RCP4.5 de 1.857; y para el escenario RCP8.5 de 1.851, respectivamente. Por lo que, el desempeño de los modelos fue excelentes, sin embargo, las pruebas del análisis de Z reafirmaron estos resultados, indicando que los modelos son estadísticamente significativos (p < 0.01).
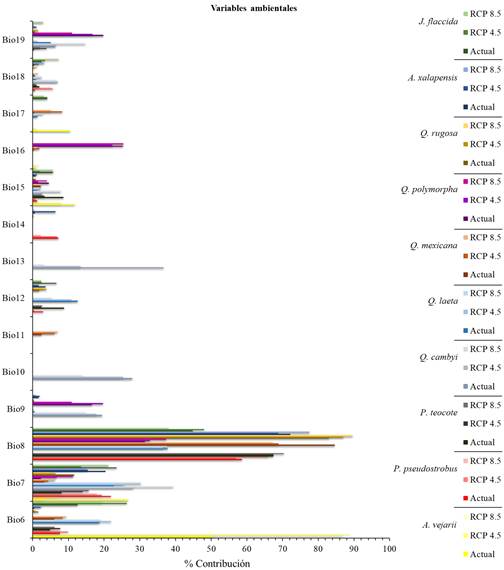
Contribución de las variables climáticas
La variable más representativa y con mayor contribución en los modelos fue la Bio8, común para ocho especies en estudio, con rangos de contribución de 36.5 a 89.6% para los escenarios actuales y futuros. La Bio7 fue relevante para las 10 especies (P. pseudostrobus, P. teocote, Q. canbyi, Q. laeta, Q. mexicana, Q. polymorpha, Q. rugosa, A. xalapensis, J. fláccida y A. vejarii) y la segunda variable con mayor contribución, con rangos de 3.2 a 30.3%. La Bio6 fue común para ocho especies (A. vejarii, P. pseudostrobus, P. teocote, Q. laeta, Q. mexicana, Q. rugosa, A. xalapensis y J. flaccida) y la tercera variable que más contribuyó, con valores que van de 0.4 a 89%, sin embargo, se aprecia que la temperatura mínima del mes más frío (°C) para A. vejarii fue la de mayor contribución tanto en el escenario actual como a futuro. La Bio19 está representada en siete especies (P. teocote, Q. canbyi, Q. laeta, Q. polymorpha, Q. rugosa, A. xalapensis y J. flaccida), siendo la cuarta variable con mayor porcentaje de contribución, con rangos de contribución de 0.1 a 19.8% (Figura 3).
Distribución potencial actual y futuro
Los resultados indican diferentes respuestas de las especies arbóreas al cambio climático. El escenario catalogado como conservador (RCP4.5) reflejan una reducción en las áreas de distribución potencial de A. vejarii 19.43%, P. pseudostrobus 15.42%, P. teocote 30.46% y Q. polymorpha 36.53%. Sin embargo, para el escenario considerado como pesimista (RCP8.5) reflejan reducciones para A. vejarii de 14.98%, P. pseudostrobus 35.76%, P. teocote 21.41% y Q. polymorpha 5.62% (Figura 4).
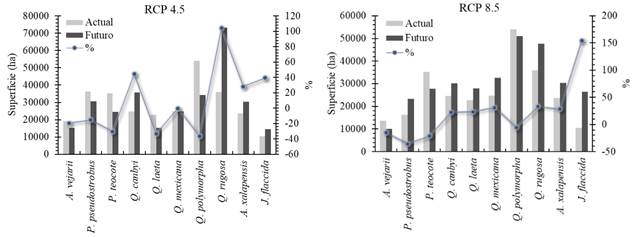
Figura 4 Área potencial de las especies analizadas de acuerdo con el escenario actual (Período 1910-2009) y bajo escenarios de cambio climático (RCP4.5 y RCP8.5 para el período 2050). Se presenta el área total que ocupan en hectáreas (ha) con sus porcentajes (%) de cambio.
Para las especies restantes correspondientes al género Quercus, Arbutus y Juniperus se observan aumentos en su distribución bajo ambos escenarios. Sin embargo, Q. rugosa correspondiente al escenario RCP4.5 estima un aumento del 104.12%, lo que supone el doble de la superficie de la distribución actual. En el caso del escenario RCP8.5 se aprecia un aumento en la especie J. flaccida del 153.73%; estos aumentos dependerán del modelo y escenario aplicado.
Se observa que, dependiendo del escenario empleado en los modelos de distribución de las especies, estas se verán afectadas diferencialmente (Figura 5). Se observa que la especie Q. laeta y Q. mexicana experimentaran reducciones de acuerdo con el escenario RCP4.5 y aumentos en el escenario RCP8.5 (Figura 5) esta irregularidad no es de esperarse, por lo que es indispensable generar diversos modelos bajo diferentes escenarios de cambio climático respecto a estas especies.
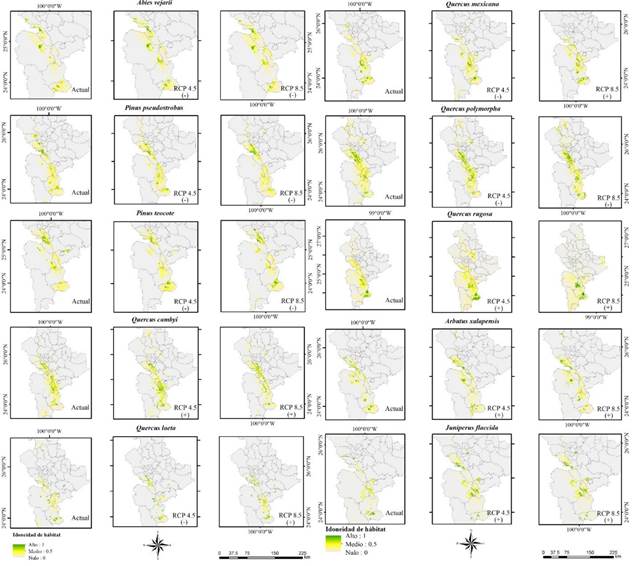
Figura 5 Distribución potencial en condiciones bioclimáticas actual (Período 1910-2009) y escenarios de cambio climático (RCP4.5 y RCP8.5 período 2050) futura de especies de clima templado en Nuevo León México. (+) aumento, (-) reducción. El tono verde en la escala de colores indica la mayor presencia de la especie, mientras que el tono más claro indica que no hay presencia.
Áreas aptas para la conservación
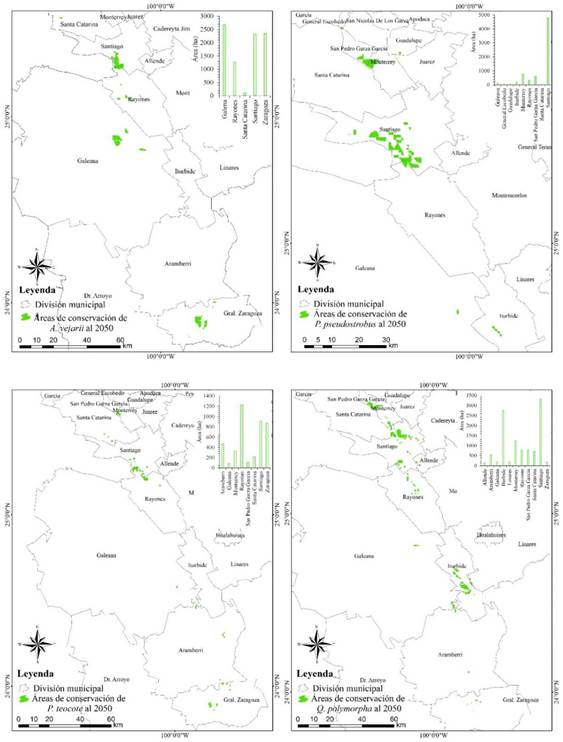
Los modelos futuros indican que cuatro de las 10 especies de clima templado sufrirán reducciones en sus áreas de distribución debido a los efectos del cambio climático, de modo que las áreas estimadas con las condiciones ambientales de idoneidad que se conservarán en la actualidad (Período 1910-2009) y se mantendrán en el futuro (década 2050) fue para A. vejarii de 8 690 ha, P. pseudostrobus de 6 852 ha, P. teocote de 4197 ha y Q. polymorpha de 10 828 ha (Figura 6). Así mismo, los resultados sugieren que ante los escenarios de cambio climático existirían algunas zonas en el estado de Nuevo León que podrían contener características ambientales para la conservación de ciertas especies. Las zonas de mayor conservación y las cuales se mantendrán para A. vejarii se encuentran principalmente en los Municipios de Galeana 2 671 ha, Gral. Zaragoza 2 347 ha y Santiago 2 311 ha. Para P. pseudostrobus en Santiago 4 787 ha. Para P. teocote en Rayones 1 225 ha, Santiago 910 ha y Gral. Zaragoza 870 ha, y Q. polymorpha en Santiago 3 313 ha, Iturbide 2 729 ha y Monterrey 1 236 ha.
Discusión
Varios autores han demostrado la utilidad de los modelos de nicho ecológico y modelos de distribución de especies para determinar los impactos de los cambios y modificaciones en la precipitación y la temperatura sobre la diversidad de especies (Sáenz-Romero et al. 2015, Gómez-Pineda et al. 2022, Manzanilla-Quijada et al. 2024a). La complementación de los registros recolectados en campo con la revisión de ejemplares de herbarios permitió obtener resultado precisos y robustos. A pesar de que Q. rugosa fue la que presento el menor número de registros, los resultados de los análisis fueron sólidos. Según Stockwell y Peterson (2002) demostraron que la modelación de cualquier especie depende de la cantidad y calidad de los datos, sugiriendo que para que la predicción sea robusta, se necesita que la ubicación geográfica sea precisa y se utilice al menos un mínimo de 50 registros. Mientras que otros requieren un muestreo mínimo de 80 observaciones (Corona-Núñez et al. 2017). Aunque se ha realizado modelación de especies con menos de seis registros (Aguirre y Duivenvoorden 2010). Sin embargo, de acuerdo con Phillips et al. (2006) y Pearson et al. (2007) es recomendable utilizar por lo menos 5 y 10 registros de presencia. Por lo que, el desempeño de los modelos no solo se encuentra relacionado con el número de observaciones, sino con la variabilidad en la distribución de la especie. Por ejemplo, Q. rugosa mostró que, pese al bajo número de registros (10), las características bioclimáticas que restringen su distribución resultaron en un modelo con buen desempeño predictivo.
Validación de los modelos
Los valores estimados señalan con exactitud las áreas de idoneidad de las especies analizadas, ya que los resultados se encuentran dentro de los rangos reportados en la literatura para ciertas especies (Aceves-Rangel et al. 2018, Manzanilla et al. 2019, González-cubas et al. 2020). Estos autores realizaron estudios sobre modelos de distribución de especies en diferentes regiones de México obteniendo resultados de AUC superiores a 0.900 para los datos de entrenamiento y validación. De acuerdo Elith et al. (2006), los modelos con valores de AUC de 0.750 son útiles para predecir la distribución de especies; en ese sentido, los valores de AUC superiores a 0.900 obtenidos en este trabajo indican que los modelos están altamente descritos por las variables bioclimáticas. Además, Ramos-Dorantes et al. (2017) obtuvieron un promedio de AUC de 0.966 en especies de clima templado en Puebla, México. Mientras que Aceves-Rangel et al. (2018) reportan valores promedio de AUC de 0.959 en especies de Pinus en México. Siendo ligeramente superiores los resultados obtenidos en este trabajo. De modo que, los modelos de distribución a través de MaxEnt, el análisis de ROC-parcial y la prueba de Z pueden considerarse fiables, ya que las predicciones son lo suficientemente precisas como para poder establecer planes de gestión.
Contribución de las variables climáticas
Si bien, la influencia de las variables climáticas difiere entre especie. Los resultados muestran que las especies evaluadas son sensibles a la oscilación de la temperatura y la precipitación. Lo anterior es similar a lo reportado por otros autores para el género (Cruz-Cárdenas et al. 2016, Guzmán-Santiago et al. 2024). Las variables de mayor contribución para las especies en estudio fue la temperatura media del trimestre más húmedo, ocupando el primer lugar en porcentaje de contribución para los modelos de ocho especies, por lo que se considera la variable bioclimática más influyente. Las variables con rango de temperatura anual (Bio5-Bio6) fue la segunda variable con diferente nivel de contribuciones para las 10 especies en estudio. Estas variables son relevantes en la distribución de las especies, por lo tanto, son de importancia en el hábitat para estos taxas. Conocer las condiciones actuales en las que se distribuyen las especies, proporciona información importante para identificar las áreas que contienen las características ambientales que estas especies requieren. Esto indica que los cambios o alteraciones en las variables más relevantes asociadas a las especies que tendrían un gran impacto en sus áreas de distribución a futuro. Por lo qué, el estudio de las variables climáticas en la distribución de las especies permite conocer las implicaciones en el desarrollo espaciotemporal de sus poblaciones a diferentes escalas, aportando información sobre el potencial efecto del cambio climático en las áreas de idoneidad donde se desarrollan estas especies y así poder decretar posibles áreas de refugio para mitigar el impacto climático a futuro.
Distribución potencial actual y futuro
El cambio climático está afectando la distribución de las especies y los ecosistemas; de acuerdo con los modelos, algunas especies se verán afectadas por el cambio climático en el año 2050. Los resultados muestran que ante el escenario optimista RCP4.5 y pesimista RCP8.5, algunas especies reducirán su área de distribución potencial (A. vejarii con una reducción de 3 707.55 ha, P. pseudostrobus con 5 592.28 ha, P. teocote con 10 737.98 ha y Q. polymorpha con 19 727.37 ha) respecto a su superficie actual. Sin embargo, para el resto de las especies (Q. canbyi, Q. laeta, Q. mexicana, Q. rugosa, A. xalapensis, J. fláccida) aumentaran su área de distribución ya que los modelos indican zonas que serán idóneas o tendrán las condiciones adecuadas para desarrollarse en el futuro. Al respecto, Hanewinkel et al. (2013) afirman que el cambio climático resultará en el reemplazo neto de bosques altamente productivos (Pinus spp.) por especies con menor productividad (Quercus spp.) para finales de siglo (Década 2100).
La capacidad de cambio y adaptación de las especies a la presencia de poblaciones en determinadas condiciones es un indicador de sus preferencias y requerimientos. Lo anterior y de acuerdo con Thuiller et al. (2011) es importante, ya que la tolerancia entre especies son diferentes y esto hace que algunas sean más vulnerables a los efectos del cambio climático. Sin embargo, es probable que los cambios en el régimen de precipitaciones, el aumento de la temperatura y la irregularidad en los eventos meteorológicos tendrá impacto en la distribución, tamaño, estructura y abundancia de las poblaciones de algunas especies arbóreas, de modo que las especies tendrán que desafiar diversas presiones fisiológicas del entorno ante el cambio climático actual y responder en tiempos relativamente cortos (IPCC 2014, Sáenz-Romero et al. 2016).
La combinación de estos factores tendrá como consecuencia regiones más cálidas y secas acorde a las proyecciones, y una tendencia hacia un aumento en la aridez, provocando estrés en las plantas al estar en condiciones que ya no son las adecuadas (Castellanos-Acuña et al. 2018, Manzanilla-Quijada et al. 2024b). Lo que lo predispondrá al ataque de plagas y enfermedades (Gómez-Pineda et al. 2022). Por ello, la permanencia de las especies ante el cambio climático dependerá de su capacidad de adaptación y de migrar a lugares con condiciones climáticas óptimas (Sáenz-Romero et al. 2016). La posibilidad de adaptación depende del nivel de variación genética y de la plasticidad fenotípica residente en las poblaciones para ajustar sus procesos a las nuevas condiciones (Tognetti et al. 2019). De acuerdo con Dawson y Spannagle (2009) sugieren que, a causa de los cambios actuales en el clima, las especies podrían desplazarse verticalmente, logrando cambiar la amplitud altitudinal de su distribución. Sin embargo, la capacidad natural de migración de las plantas a través de la dispersión y su establecimiento de nuevos hábitats es demasiado lenta para la mayoría de las especies (Soto-Correa et al. 2014). Estudios publicados, han mostrado que los procesos de migración asistida de especies de climas templados están dirigidas a porciones del territorio con mayor altitud, siendo estas zonas donde hay mayor precipitaciones y temperaturas más bajas (Mirhashemi et al. 2023). Sin embargo, Byars et al. (2009) señalan que una de las mejores opciones de movimiento de las plantas es hacia sitios con mayores elevaciones, donde se espera que ocurran las condiciones óptimas a las que están adaptadas. Al respecto Gavilán (2008), Denvir et al. (2022) señalan que es importante, reconocer que la pérdida de la biodiversidad no sólo se debe al cambio climático, sino también a otros factores asociados, que tienen efectos a corto plazo como la fragmentación del hábitat de los bosques nativos, el cambio de uso de suelo para la ganadería, la agricultura, la urbanización, especies invasoras, entre otros. Por lo que, la pérdida de especies será probablemente mucho mayor que la proyectada en este trabajo. Si bien, la fragmentación de hábitats de los bosques nativos puede propiciar una serie de efectos adversos en las especies arbóreas, estos efectos incluyen la disminución del tamaño de las poblaciones, la limitación del intercambio genético entre fragmentos aislados, el incremento de la endogamia, la reducción de la vitalidad de las plántulas e, incluso en situaciones extremas, la extinción local de las especies (Aguilar et al. 2006). La reducción del tamaño poblacional como resultado de la fragmentación de hábitats implica que las poblaciones arbóreas sean más susceptibles a la pérdida de variabilidad genética y a los efectos negativos de la deriva genética, a razón de las limitaciones del flujo génico entre fragmentos aislados, lo que dificulta el intercambio de genes y la renovación genética de las poblaciones.
El IPCC 2021 informa que la temperatura global del aire en la superficie ha aumentado aproximadamente 1°C por encima de los niveles preindustriales. Sin embargo, investigaciones publicadas por Dorji et al. (2013) y Zi et al. (2022) señalan que cada vez hay más pruebas de que el calentamiento del clima modifica el rendimiento reproductivo sexual de las plantas, incluida la fenología de la floración y el esfuerzo reproductivo (la proporción de los recursos de un organismo destinados a la reproducción, por ejemplo, el número de flores) y el éxito (el resultado final de la inversión de recursos, por ejemplo, el número de frutos y/o semillas). Lo anterior se sustenta con lo reportado por Zi et al. (2022) quienes reportan que el calentamiento climático (aumento en la temperatura) reduce el número de frutos en determinadas especies, una explicación para el efecto negativo del calentamiento sobre el número de frutos es que el estrés por altas temperaturas altera el desarrollo del embrión y el endospermo. Por lo tanto, los patrones adaptativos pueden verse afectados por el aumento en la temperatura o las alteraciones en los regímenes de precipitaciones podrían ocasionar desacoplamiento entre los ambientes y los genotipos (Sáenz-Romero et al. 2010).
Por otra parte, hay que tener en cuenta que trabajar con escenarios de cambio climático no es predecir el futuro, sino comprender mejor las incertidumbres para tomar decisiones que sean robustas ante un amplio abanico de posibles escenarios futuros. En este trabajo, bajo cualquier de los escenarios, se prevé que el área de distribución aumente o disminuya con referencia al clima actual (Figura 5). También pueden utilizarse para comparar las características biológicas de otras especies que se benefician de encontrarse con un aumento de áreas en condiciones ambientales adecuadas, en contraste con aquellas especies que experimentan una respuesta negativa.
Áreas aptas para la conservación
México enfrenta el reto de gestionar adecuadamente sus recursos forestales en un territorio privilegiado por una enorme biodiversidad y una diversidad cultural de pueblos que han participado durante siglos en el aprovechamiento de múltiples recursos bióticos (Gavito et al. 2017). Indudablemente esta transformación ambiental plantea desafíos para la conservación de la biodiversidad, generando la necesidad de implementar estrategias de manejo que reduzcan las presiones actuales sobre los ecosistemas forestales del estado de Nuevo León.
La combinación de los conocimientos derivados de los modelos de distribución potencial identificó zonas prioritarias para la conservación de las especies, que mantendrán su hábitat climático en el futuro (Década de 2050), de modo que, se podrían tomar decisiones y establecer estrategias de manejo para su conservación y/o recuperación. Algunos estudios recomiendan examinar las especies sensibles al cambio climático para definir estrategias de conservación ex situ (Viveros-Viveros et al. 2009, Sáenz-Romero et al. 2012), y planes de manejo para la adaptación de las plantas ante el cambio climático (Oliver et al. 2016). Con el desarrollo de los modelos presentados, es posible inferir los efectos de un posible cambio climático sobre la fisiología de la especie y, por tanto, sobre su distribución. El seguimiento de estas recomendaciones es de interés, ya que hay especies que se han visto afectadas y por ende sus áreas de distribución, como es el caso de A. vejarii, una de las especies que está catalogada como Casi Amenazada con una población en disminución (SEMARNAT 2010, Farjon 2013). Especie que podría desaparecer debido a su restringido intervalo óptimo térmico, que probablemente no le permitirá sobrevivir donde se encuentra actualmente. Sin embargo, esta especie podría acceder a sitios más favorables a mayor altitud donde la temperatura sería más adecuada para su desarrollo, pero esta migración depende principalmente de la dispersión de sus semillas, en el sentido de que algún vector sería capaz de transportarlas a altitudes superiores a los sitios donde se encuentra actualmente. Sin embargo, Sáenz-Romero et al. (2016) señalan que este proceso es relativamente lento y depende de la capacidad de migración altitudinal de la especie. Mientras que Sáenz-Romero et al. (2016), recomiendan reubicar a las especies cuyo hábitat se verá afectado por el cambio climático a sitios donde se prevé que en el futuro contaran con las condiciones favorables para la especie mediante Migración asistida. Es decir, la plantación de plántulas procedentes en un sitio de recolección de semillas en el presente debe tener los mismos valores climáticos que el clima que ocurrirá en un futuro previsible en un sitio a reforestar tal y como sugiere (Castellanos-Acuña et al. 2018, Manzanilla-Quijada et al. 2024b).
Conclusiones
Es evidente el efecto del cambio climático sobre la distribución de las especies estudiadas en el bosque templado del estado de Nuevo León. El análisis realizado es una aproximación para reconocer la severidad del cambio climático sobre algunas de las especies arbóreas. Permitiendo, estimar la distribución potencial actual y futura e identificando áreas de conservación que se mantendrán en el futuro. De esta manera, los tomadores de decisiones encargados de seleccionar sitios para la conservación dispongan de argumentos adicionales con mejores fundamentos ecológicos y biogeográficos. Las especies más afectadas por el cambio climático son A. vejarii, P. pseudostrobus, P. teocote y Q. polymorpha. Los resultados obtenidos pueden complementar las estrategias para la conservación de algunas especies del bosque templado en el estado Nuevo León.

