Scientific articles
Diversity and updated checklist of limno-terrestrial tardigrades species in Quintana Roo, Mexico
Pérez-Pech, Wilbert A.1*
 http://orcid.org/0000-0002-2375-4468
http://orcid.org/0000-0002-2375-4468
Guidetti, Roberto2
http://orcid.org/0000-0001-6079-2539
Anguas-Escalante, Abril1
http://orcid.org/0000-0001-7909-1475
Crisanto-Rodríguez, Juana I.3
http://orcid.org/0009-0009-7556-6096
Cutz-Pool, Leopoldo Q.3
http://orcid.org/0009-0009-4962-140X
http://orcid.org/0000-0002-2375-4468Guidetti, Roberto2
http://orcid.org/0000-0001-6079-2539Anguas-Escalante, Abril1
http://orcid.org/0000-0001-7909-1475Crisanto-Rodríguez, Juana I.3
http://orcid.org/0009-0009-7556-6096Cutz-Pool, Leopoldo Q.3
http://orcid.org/0009-0009-4962-140X
-
Publication dates-
July 28, 2025
Jan-Dec , 2025
- Article in PDF
- Article in XML
- Automatic translation
- Send this article by e-mail
- Share this article +
Abstract
Introduction
Quintana Roo is the only state in the Yucatán Peninsula where the limno-terrestrial tardigrade fauna has been documented; however, the records have been limited to generic reports.
Objective
To provide an updated checklist of limno-terrestrial tardigrades for Quintana Roo and to explore the spatiotemporal variation of these animals in two rural environments.
Materials and methods
A taxonomic review was conducted on tardigrades collected between 2016 and 2018. Moreover, during two sampling campaigns in 2018, specimens were collected from the moss Aulacopilum sp. and the lichen Parmotrema sp. in a citrus plantation and an evergreen forest fragment. Data were compared using the student’s t test (P < 0.05).
Results and discussion
Kristenseniscus kofordi, Milnesium sp. 1, Milnesium cf. cassandrae , Macrobiotus alvaroi, Macrobiotus cf. polyopus, Macrobiotus cf. ocotensis, Macrobiotus sp. 1, Macrobiotus cf. semmelweisi, Minibiotus cf. intermedius, Paramacrobiotus cf. savai, Mesobiotus cf. pseudopatiens and Ramazzottius cf. baumanni were recorded. The addition of these taxa, along with previous records of Echiniscus sp. and Doryphoribius chetumalensis, increases the number of limno-terrestrial tardigrade species in Quintana Roo to 14, also the coastal mangrove forest is added to the list of environments in the entity. The highest species richness and Shannon-Wiener diversity were observed in the citrus plantation.
Conclusions.
The updated checklist of limno-terrestrial tardigrades in Quintana Roo comprises 14 species. Diversity was higher in the citrus plantation, suggesting that tardigrades adapt well to anthropogenic environments.
Keywords:
Kristenseniscus kofordi, Macrobiotus alvaroi, Milnesium sp., Rhizophora mangle
Introduction
Tardigrades inhabit marine, freshwater, and terrestrial ecosystems (Nelson et al., 2015). Research on this group has primarily focused on describing and listing limno-terrestrial species in natural environments (Bartels et al., 2016; Nelson et al., 2015). However, urban and rural environments can also host unique communities of limno-terrestrial tardigrades (González-Reyes et al., 2016; Guidetti et al., 2013; Meyer et al., 2013; Pérez-Pech et al., 2017).
-
Nelson et al., 2015Phylum Tardigrada, 2015
-
Bartels et al., 2016A global biodiversity estimates of a poorly known taxon: phylum TardigradaZoological Journal of the Linnean Society, 2016
-
Nelson et al., 2015Phylum Tardigrada, 2015
-
González-Reyes et al., 2016Evaluación de la diversidad en comunidades de tardígrados (Ecdysozoa, Tardigrada) en hábitats urbano y rural de la ciudad de Salta (Argentina)Iheringia Série Zoologia, 2016
-
Guidetti et al., 2013The morphological and molecular analyses of a new South American urban tardigrade offer new insights on the biological meaning of the Macrobiotus hufelandi group of species (Tardigrada: Macrobiotidae)Journal of Natural History, 2013
-
Meyer et al., 2013Water bears in the Anthropocene: a comparison of urban and woodland tardigrade (Phylum Tardigrada) communities in Southwestern Louisiana, USAJournal of Limnology, 2013
-
Pérez-Pech
et al., 2017Doryphoribius chetumalensis sp. nov. (Eutardigrada: Isohypsibiidae) a new tardigrade species discovered in an unusual habitat of urban areas of MexicoZootaxa, 2017
In recent years, interest in studying tardigrade fauna in Mexico has increased (Dueñas-Cedillo et al., 2020; García-Román et al., 2022; Moreno-Talamantes et al., 2015, 2019; Moreno-Talamantes & León-Espinosa, 2019; Núñez et al., 2021). In the Yucatan Peninsula, only Quintana Roo has focused on the study of this fauna. Pérez-Pech et al. (2016, 2017) recorded tardigrades in urban environments in Chetumal and described Doryphoribius chetumalensisPérez-Pech, Anguas-Escalante, Cutz-Pool & Guidetti, 2017. Moreover, Anguas-Escalante et al. (2018) and Cutz-Pool et al. (2019) studied tardigrade diversity in citrus plantations. At the national level, Quintana Roo is also a pioneer in the study of coastal marine tardigrades (Anguas-Escalante et al., 2020; Pérez-Pech et al., 2020).
-
Dueñas-Cedillo et al., 2020Tardigrades from Iztaccíhuatl Volcano (Trans-Mexican Volcanic Belt), with the description of Minibiotus citlalium sp. nov. (Eutardigrada: Macrobiotidae)Diversity, 2020
-
García-Román et al., 2022A strategy to provide a present and future scenario of Mexican biodiversity of tardigradaDiversity, 2022
-
Moreno-Talamantes et al., 2015First record of Dactylobiotus parthenogeneticus Bertolani, 1982 (Eutardigrada: Murrayidae) in MexicoCheck List, 2015
-
2019Current knowledge on Mexican tardigrades with a description of Milnesium cassandrae sp. nov. (Eutardigrada: Milnesiidae) and discussion on the taxonomic value of dorsal pseudoplates in the genus Milnesium Doyère, 1840Zootaxa, 2019
-
Moreno-Talamantes & León-Espinosa, 2019Nuevo registro de Diaforobiotus islandicus (Richters, 1904) (EUTARDIGRADA: RICHTERSIIDAE) para MéxicoArido-Ciencia, 2019
-
Núñez et al., 2021First tardigrade records from San Pedro Mártir, Baja California, MexicoCheck List, 2021
-
Pérez-Pech et al. (2016Primer registro genérico de tardígrados, habitantes del área urbana de Chetumal Quintana Roo, MéxicoEntomología Mexicana, 2016
-
2017)Doryphoribius chetumalensis sp. nov. (Eutardigrada: Isohypsibiidae) a new tardigrade species discovered in an unusual habitat of urban areas of MexicoZootaxa, 2017
-
Pérez-Pech, Anguas-Escalante, Cutz-Pool & Guidetti, 2017Doryphoribius chetumalensis sp. nov. (Eutardigrada: Isohypsibiidae) a new tardigrade species discovered in an unusual habitat of urban areas of MexicoZootaxa, 2017
-
Anguas-Escalante et al. (2018)Tardígrados asociados a una plantación de cítricos de traspatio en la comunidad de El Palmar en Quintana Roo, MéxicoInvestigación y Ciencia, 2018
-
Cutz-Pool et al. (2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Anguas-Escalante et al., 2020A new species of Tardigrada from a Caribbean reef lagoon, Florarctus yucatanensis sp. nov. (Halechiniscidae: Florarctinae)CBM - Cahiers de Biologie Marine, 2020
-
Pérez-Pech et al., 2020Marine Tardigrada from the Mexican Caribbean with the description of Styraconyx robertoi sp. nov. (Arthrotardigrada: Styraconyxidae)Zootaxa, 2020
The present study aimed to update the species checklist and records of tardigrades in Quintana Roo, and to compare the diversity of limno-terrestrial tardigrades between a citrus plantation and an evergreen forest fragment.
Materials and Methods
Variation in tardigrade diversity between an extensive temporary citrus plantation and an adjacent evergreen forest fragment was evaluated in the ejido El Palmar, municipality of Othón Pompeyo Blanco (18° 26' 48" N, 88° 31' 50" W). Collections were made during two sampling campaigns, one in February (the 'nortes' season, a meteorological phenomenon characterized by intense, cold winds) and another in September (the rainy season) of 2018. In both the plantation and vegetation fragment, a 5 000 m2 plot was established, where two transects were located in the internal and external region (Figure 1). On each transect, six trees were selected (spaced 10 m apart); from each tree, 10 cm2 samples of the moss Aulacopilum sp. and the lichen Parmotrema sp. (the only genera present in the study area) were collected at a height of 1 m. At each site, the percentage of humidity and temperature were measured using a digital thermos hygrometer (Onset HOBO© data logger).
Thumbnail
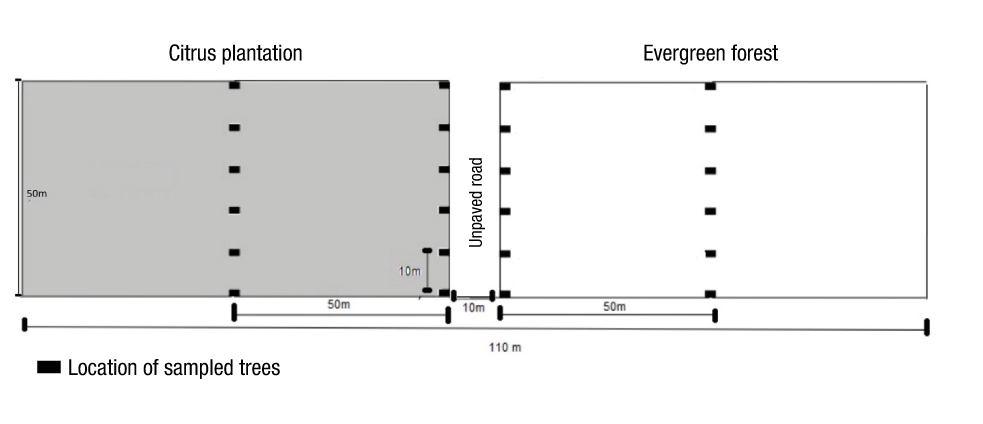
Figure 1
Sampling design of limno-terrestrial tardigrades in trees of different vegetation in the ejido El Palmar, municipality of Othón Pompeyo Blanco, Quintana Roo.
Sampling design of limno-terrestrial tardigrades in trees of different vegetation in the ejido El Palmar, municipality of Othón Pompeyo Blanco, Quintana Roo.
In the Zoology laboratory of the Tecnológico Nacional de México campus Chetumal, the samples were immersed in filtered tap water for 10 min and the pH was measured, following the methodology of Johansson et al. (2011); subsequently, the samples were sieved (mesh size 2 000 μm and 38 μm). The obtained was reviewed with a stereomicroscope Nikon C-PS ZMZ445.
-
Johansson et al.
(2011)Are urban and rural tardigrade (Tardigrada) communities distinct and determined by pH: a case study from Fresno County, CaliforniaThe Pan-Pacific Entomologist, 2011
The tardigrades were preserved in 70 % alcohol and subsequently they were mounted in glass slides using Hoyer's liquid as medium mount. Taxonomic identifications were made with Axio Lab a1 (Zeiss) and DM RB (Leica) optical microscopes (up to 100x), using differential interference contrast (DIC) and phase contrast (PhC) using dichotomous keys (Milnesium [Morek et al., 2016], Macrobiotus [Kaczmarek & Michalczyk, 2017], Mesobiotus [Kaczmarek et al., 2020] Minibiotus [Claxton, 1998] and Paramacrobiotus [Kaczmarek et al., 2017]) and primary literature (Guidetti et al., 2019; Morek et al., 2016). The slides were deposited in the Tardigrade Collection associated with the Marine Nematode Collection of the Colegio de la Frontera Sur, in the Bertolani Collection of the Universidad de Modena y Reggio Emilia (Italy), and the Tardigrade Collection related to the Mite Collection of Mexico (CNAC) of the Instituto de Biología of the Universidad Nacional Autónoma de México.
-
Morek
et al., 2016Experimental taxonomy exposes ontogenetic variability and elucidates the taxonomic value of claw configuration in Milnesium Doyère, 1840 (Tardigrada: Eutardigrada: Apochela)Contributions to Zoology, 2016
-
Kaczmarek & Michalczyk, 2017The Macrobiotus hufelandi group (Tardigrada) revisitedZootaxa, 2017
-
Kaczmarek et al., 2020Revisiting the genus Mesobiotus (Eutardigrada, Macrobiotidae)-remarks, updated dichotomous key and an integrative description of new species from MadagascarZoologischer Anzeiger, 2020
-
Claxton,
1998A revision of the genus Minibiotus (Tardigrada: Macrobiotidae) with descriptions of eleven new species from AustraliaRecords of the Australian Museum, 1998
-
Kaczmarek et al., 2017Revision of the genus Paramacrobiotus Guidetti et al., 2009 with the description of a new species, re-descriptions and a keyAnnales Zoologici, 2017
-
Guidetti et al., 2019High diversity in species, reproductive modes and distribution within the Paramacrobiotus richtersi complex (Eutardigrada, Macrobiotidae)Zoological Letters, 2019
-
Morek et
al., 2016Experimental taxonomy exposes ontogenetic variability and elucidates the taxonomic value of claw configuration in Milnesium Doyère, 1840 (Tardigrada: Eutardigrada: Apochela)Contributions to Zoology, 2016
To obtain the species checklist, samples available in the Zoology laboratory of the Tecnológico Nacional de México campus Chetumal, and the Tardigrade Collection associated with the Marine Nematode Collection of the College of the Colegio de la Frontera Sur were examined. These samples were compared with the taxa identified in the plantation and forest fragment and with specimens from samples of Rhizophora mangle L. stems from Chetumal Bay. To achieve this, 33 permanent slides from the work of Anguas-Escalante et al. (2018), conducted in an intensive citrus plantation in 2016 in El Palmar (18° 26' 48.0" N, 88° 31' 47.0" W), were reviewed. From the same locality, 20 samples (lichen and moss) collected by Cutz-Pool et al. (2019) in an extensive citrus plantation, and 48 samples (lichen and moss) collected by Pérez-Pech in 2018 in an evergreen forest fragment (unpublished data) were also reviewed. Finally, three lichen samples collected in February 2018 by Pérez-Pech (unpublished data) from coastal mangrove stems were examined (18° 31' 14.1" N, 88° 16' 09.4" W).
-
Anguas-Escalante
et al. (2018)Tardígrados asociados a una plantación de cítricos de traspatio en la comunidad de El Palmar en Quintana Roo, MéxicoInvestigación y Ciencia, 2018
-
Cutz-Pool et al. (2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
Abundance (N) and richness (S) of tardigrades were calculated. Diversity was assessed using species accumulation curves (Chao1 estimator) and the Shannon-Wiener index (H´). The difference in tardigrade diversity between the sampled environments and temporal variation was determined using Student's t-tests in Past 2.17c (Hammer et al., 2001). Environmental variables were correlated with tardigrade diversity through multidimensional scaling (MDS) analysis in Primer 6 (Clarke et al., 2006). For this, data on ecosystem type (citrus plantation and forest fragment), humidity, temperature, pH, and type of microecosystem (mosses and lichens) were evaluated.
-
Hammer et al., 2001PAST: Paleontological statistics software package education and analysisPaleontologia Electronica, 2001
-
Clarke et al., 2006PRIMER V6: User Manual/tutorial, 2006
Results
Species of the phylum tardigrada (Spallanzani, 1777)
Kristenseniscus kofordi (Schuster & Grigarick, 1966)
This species belongs to the genus Kristenseniscus Gąsiorek, Morek, Stec & Michalczyk, 2019 (Table 1). A total of 69 specimens collected from an extensive citrus plantation were analyzed. The specimens showed cuticular thickenings on the first and second pairs of plates and on the scapular and terminal plates. These features are characteristic of K. kofordi (Figure 2), a species that has been recorded in the state of Chiapas (Pilato, 2006), so there is a high probability that it belongs to the same species.
-
Pilato,
2006Remarks on the Macrobiotus polyopus group, with the description of two new species (Eutardigrada, Macrobiotidae)Zootaxa, 2006
Table 1
Updated list of limno-terrestrial tardigrades in Quintana Roo and biotopes (moss Aulacopilum sp., Lichen Parmotrema sp., and Mangrove Rhizophora mangle).
Updated list of limno-terrestrial tardigrades in Quintana Roo and biotopes (moss Aulacopilum sp., Lichen Parmotrema sp., and Mangrove Rhizophora mangle).
| Taxa | Record | Microecosystem | Source |
|---|---|---|---|
| Class: Heterotardigrada Marcus, 1927 Order: Echiniscoidea Richters, 1926 Family: Echiniscidae Thulin, 1928 | Echiniscus sp.1 | Soil in urban area | Pérez Pech et al. (2016, 2017) |
| Kristenseniscus kofordi (Schuster & Grigarick, 1966) | Lichen and moss | Cutz-Pool et al. (2019); this study | |
| Class: Eutardigrada Richters, 1926 Order: Apochela Schuster, Nelson, Grigarick & Christenberry, 1980 Family: Milnesiidae Ramazzotti, 1962 | Milnesium sp. 1 | Lichen | Anguas-Escalante et al. (2018) |
| Milnesium cf. cassandrae | Lichen, mossy and mangle | Anguas-Escalante et al. (2018); Cutz-Pool et al. (2019); this study | |
| Class: Eutardigrada Richters, 1926 Order: Apochela Schuster, Nelson, Grigarick & Christenberry, 1980 Family: Doryphoribiidae Gąsiorek, Stec, Morek & Michalczyk, 2019 |
Doryphoribius
chetumalensis |
Soil in urban area | Pérez Pech et al. (2016, 2017) |
| Class: Eutardigrada Richters, 1926 Order: Parachela Schuster, Nelson, Grigarick, Christenberry, 1980 Family: Ramazzottiidae Sands, McInnes, Marley, Goodall-Copestake, Convey & Linse, 2008 | Ramazzottius cf. baumanni | Lichen | Anguas-Escalante et al. (2018) |
| Class: Eutardigrada Richters, 1926 Order: Apochela Schuster, Nelson, Grigarick & Christenberry, 1980 Family: Macrobiotidae Thulin, 1928 | Macrobiotus cf. semmelweisi | Moss and lichen | Cutz-Pool et al. (2019); this study |
| Macrobiotus cf. polyopus | Moss and lichen | Cutz-Pool et al. (2019); this study | |
| Macrobiotus sp. 1 | Moss and lichen | Cutz-Pool et al. (2019); this study | |
| Macrobiotus alvaroiPilato & Kaczmarek 2007 | Moss and lichen | Cutz-Pool et al. (2019); this study | |
| Macrobiotus cf. ocotensis | Lichen | Cutz-Pool et al. (2019) | |
| Mesobiotus cf. pseudopatiens | Moss and lichen | Anguas-Escalante et al. (2018), Cutz-Pool et al (2019), this study | |
| Minibiotus cf. intermedius | Moss and lichen | Cutz-Pool et al. (2019); this study | |
| Paramacrobiotus cf. savai | Moss and lichen | Cutz-Pool et al. (2019); this study |
-
Pérez Pech et al.
(2016Primer registro genérico de tardígrados, habitantes del área urbana de Chetumal Quintana Roo, MéxicoEntomología Mexicana, 2016
-
2017)Doryphoribius chetumalensis sp. nov. (Eutardigrada: Isohypsibiidae) a new tardigrade species discovered in an unusual habitat of urban areas of MexicoZootaxa, 2017
-
Cutz-Pool et al.
(2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Anguas-Escalante et
al. (2018)Tardígrados asociados a una plantación de cítricos de traspatio en la comunidad de El Palmar en Quintana Roo, MéxicoInvestigación y Ciencia, 2018
-
Anguas-Escalante et
al. (2018)Tardígrados asociados a una plantación de cítricos de traspatio en la comunidad de El Palmar en Quintana Roo, MéxicoInvestigación y Ciencia, 2018
-
Cutz-Pool et al.
(2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Pérez-Pech,
Anguas-Escalante, Cutz-Pool & Guidetti,
2017Doryphoribius chetumalensis sp. nov. (Eutardigrada: Isohypsibiidae) a new tardigrade species discovered in an unusual habitat of urban areas of MexicoZootaxa, 2017
-
Pérez Pech et al.
(2016Primer registro genérico de tardígrados, habitantes del área urbana de Chetumal Quintana Roo, MéxicoEntomología Mexicana, 2016
-
2017)Doryphoribius chetumalensis sp. nov. (Eutardigrada: Isohypsibiidae) a new tardigrade species discovered in an unusual habitat of urban areas of MexicoZootaxa, 2017
-
Anguas-Escalante et al.
(2018)Tardígrados asociados a una plantación de cítricos de traspatio en la comunidad de El Palmar en Quintana Roo, MéxicoInvestigación y Ciencia, 2018
-
Cutz-Pool et al.
(2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Cutz-Pool et
al. (2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Cutz-Pool et al.
(2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Pilato & Kaczmarek
2007Macrobiotus alvaroi, a new species of eutardigrade (Tardigrada, Macrobiotidae) of the polyopus group from Costa Rica (Central America)Zootaxa, 2007
-
Cutz-Pool et
al. (2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Cutz-Pool et
al. (2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Anguas-Escalante et al.
(2018)Tardígrados asociados a una plantación de cítricos de traspatio en la comunidad de El Palmar en Quintana Roo, MéxicoInvestigación y Ciencia, 2018
-
Cutz-Pool et al (2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Cutz-Pool et al.
(2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Cutz-Pool et al.
(2019)Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
Thumbnail
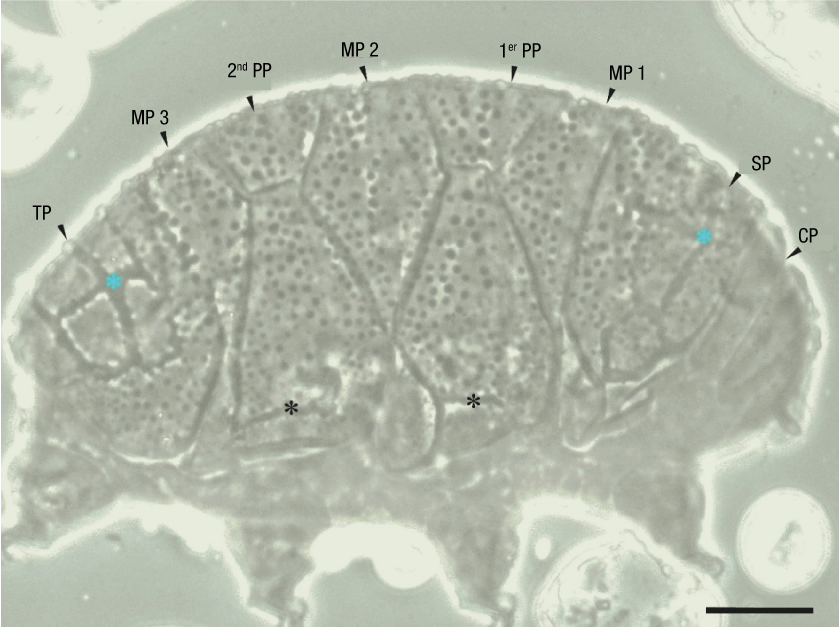
Figure 2
Dorsal region of Kristenseniscus kofordi. Blue asterisks indicate band-like thickenings that are interrupted, while the black asterisks indicate longitudinal band-like thickenings. Scapular plate (SP), cephalic plate (CP), median plate (MP), plate pair (PP), terminal plate (TP). Scale 20 µm.
Dorsal region of Kristenseniscus kofordi. Blue asterisks indicate band-like thickenings that are interrupted, while the black asterisks indicate longitudinal band-like thickenings. Scapular plate (SP), cephalic plate (CP), median plate (MP), plate pair (PP), terminal plate (TP). Scale 20 µm.
Milnesium Doyère, 1840
Milnesium sp. 1. A single individual was examined from an intensive citrus plantation. The specimen showed a [3-3]-[3-3/4] claw configuration and a smooth cuticle.
Milnesium cf. cassandrae. A total of 926 specimens were examined, collected from an extensive citrus plantation and an evergreen forest fragment; 10 specimens were collected from an intensive citrus plantation; and 17 specimens were found on lichen growing on the stems of R. mangle in the coastal area of Chetumal.
According to the identification key for the granulatum group, the specimens are closely related to Milnesium cassandraeMoreno-Talamantes, Roszkowska, García-Aranda, Flores-Maldonado & Kaczmarek, 2019, due to the presence of the [2-2]-[2-2] claw configuration. However, differences were observed in the pt index values (ratio of a character's length to the buccal tube length) at the insertion point of the support stylet, which exceeded 73 µm (ranging from 75.18 to 93.14 µm), and in the buccal tube length, which was greater than 23 µm (ranging from 30.54-49.09 µm).
-
Moreno-Talamantes,
Roszkowska, García-Aranda, Flores-Maldonado & Kaczmarek,
2019Current knowledge on Mexican tardigrades with a description of Milnesium cassandrae sp. nov. (Eutardigrada: Milnesiidae) and discussion on the taxonomic value of dorsal pseudoplates in the genus Milnesium Doyère, 1840Zootaxa, 2019
Based on the description provided for M. cassandrae (Moreno-Talamantes et al., 2019), the collected specimens match the [2-2]-[2-2] claw configuration in juveniles and the [2-3] [3-2] configuration in adults, as well as the reticular pattern of the dorsal cuticle (Figure 3).
-
Moreno-Talamantes et al., 2019Current knowledge on Mexican tardigrades with a description of Milnesium cassandrae sp. nov. (Eutardigrada: Milnesiidae) and discussion on the taxonomic value of dorsal pseudoplates in the genus Milnesium Doyère, 1840Zootaxa, 2019
Thumbnail
![Diagnostic characters of the collected specimens of
Milnesium cf. cassandrae A) Claw
configuration [2-3] on leg III and B) claw configuration [3-2]
on leg VI in adult specimens. C) Claw configuration [2-2] on leg
II and D) Claw configuration [2-2] on leg VI in juvenile
specimens. E) Reticular pattern of the dorsal cuticle in adult
specimens and F) in juveniles. Internal claw (IC), external claw
(EC). Scale = 20 µm.](/img/revistas/rcscfa/v31//2007-4018-rcscfa-31-e24013-gf3.png)
Figure 3
Diagnostic characters of the collected specimens of Milnesium cf. cassandrae A) Claw configuration [2-3] on leg III and B) claw configuration [3-2] on leg VI in adult specimens. C) Claw configuration [2-2] on leg II and D) Claw configuration [2-2] on leg VI in juvenile specimens. E) Reticular pattern of the dorsal cuticle in adult specimens and F) in juveniles. Internal claw (IC), external claw (EC). Scale = 20 µm.
Diagnostic characters of the collected specimens of Milnesium cf. cassandrae A) Claw configuration [2-3] on leg III and B) claw configuration [3-2] on leg VI in adult specimens. C) Claw configuration [2-2] on leg II and D) Claw configuration [2-2] on leg VI in juvenile specimens. E) Reticular pattern of the dorsal cuticle in adult specimens and F) in juveniles. Internal claw (IC), external claw (EC). Scale = 20 µm.
Another species morphologically similar to this taxon, is Milnesium katarzynae Kaczmarek, Michalczyk & Beasley, 2004. However, M. katarzynae was described in China, while M. cassandrae is a native species from Mexico (Morek et al., 2022).
-
Morek et al., 2022Rough backs: taxonomic value of epicuticular sculpturing in the genus Milnesium Doyère, 1840 (Tardigrada: Apochela)Scientific Reports, 2022
Ramazzottius cf. baumanni
This species belongs to the genus Ramazzottius Binda & Pilato, 1986. A total of 11 adult specimens and five eggs were examined, collected from an intensive citrus plantation. The animals exhibited a sculpted dorsal cuticle similar to that of Ramazzottius baumanni (Ramazzotti, 1962) (Figure 4A). However, the eggs of this species remain unknown. In this study, eggs were collected, but an extensive comparison with other species of the genus is necessary to determine whether it represents a new species.
Thumbnail
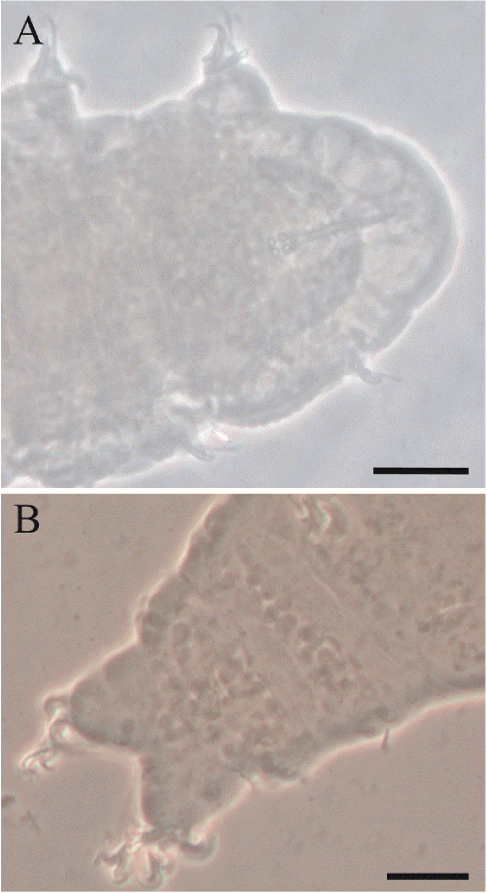
Figure 4
A) Sculpted cuticle of specimens identified as Ramazzottius cf. baumanni. B) Gibbosities of the dorsal cuticle of specimens identified as Macrobiotus cf. semmelweisi. Scale = 20 µm.
A) Sculpted cuticle of specimens identified as Ramazzottius cf. baumanni. B) Gibbosities of the dorsal cuticle of specimens identified as Macrobiotus cf. semmelweisi. Scale = 20 µm.
Macrobiotus C. A. S. Schultze 1834
Macrobiotus alvaroiPilato and Kaczmarek, 2007. A total of 194 adult specimens and 15 eggs were examined, collected from an extensive citrus plantation. This species belongs to the polyopus species complex (Marcus, 1928) of the genus Macrobiotus. The specimens collected share morphological similarities with M. alvaroi and Macrobiotus ocotensisPilato, 2006.
-
Pilato
and Kaczmarek, 2007Macrobiotus alvaroi, a new species of eutardigrade (Tardigrada, Macrobiotidae) of the polyopus group from Costa Rica (Central America)Zootaxa, 2007
-
Pilato, 2006Remarks on the Macrobiotus polyopus group, with the description of two new species (Eutardigrada, Macrobiotidae)Zootaxa, 2006
The morphology of the processes in the collected eggs matched the descriptions of M. alvaroi, which features a hexagonal prism-shaped basal portion, a conical distal portion with an apex, and egg walls with a regular reticular design composed of hexagonal meshes (Pilato & Kaczmarek, 2007). Additionally, the buccal apparatus morphometry observed in both embryonated eggs and adult specimens corresponds to that of M. alvaroi, including the thickening at the end of the buccal tube, where the pharyngeal apophyses are connected (Figure 5).
-
Pilato & Kaczmarek, 2007Macrobiotus alvaroi, a new species of eutardigrade (Tardigrada, Macrobiotidae) of the polyopus group from Costa Rica (Central America)Zootaxa, 2007
Thumbnail
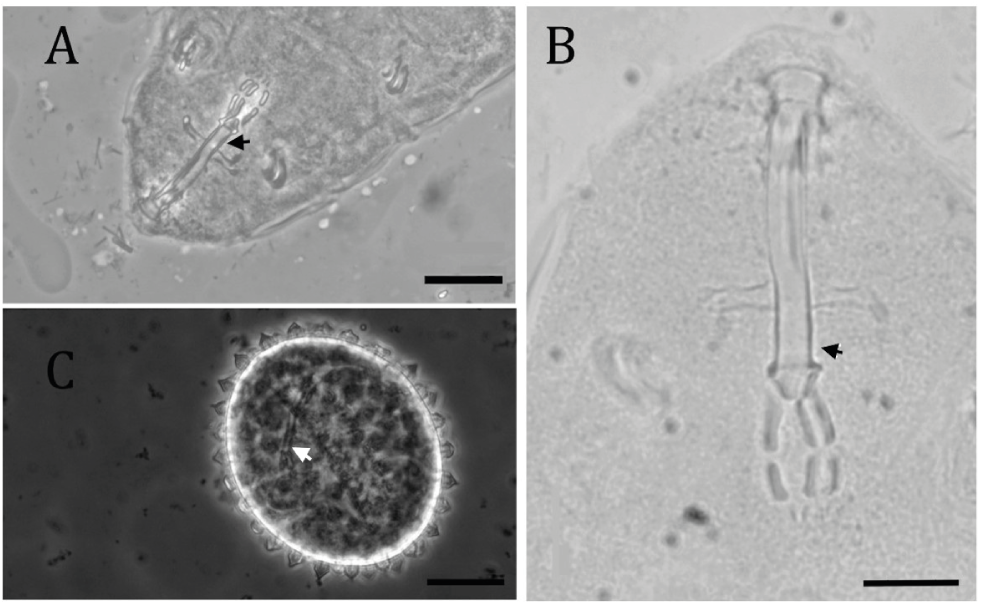
Figure 5
Buccal apparatus (A and B) and embryonated egg (C) of specimens identified as Macrobiotus alvaroi. Arrows indicate thickening at the end of the buccal tube. Scale = 20 µm.
Buccal apparatus (A and B) and embryonated egg (C) of specimens identified as Macrobiotus alvaroi. Arrows indicate thickening at the end of the buccal tube. Scale = 20 µm.
The most significant differences between M. alvaroi and M. ocotensis are the shape of the egg processes and the buccal tube. The egg processes have a hexagonal prism-shaped basal portion, a conical distal portion with an apical tip, and a shell with a regular reticular design composed of hexagonal meshes (typical of M. alvaroi). The buccal tube shows thickening. These characteristics, typical of M. alvaroi, were observed in the examined specimens (Figure 6A).
Thumbnail
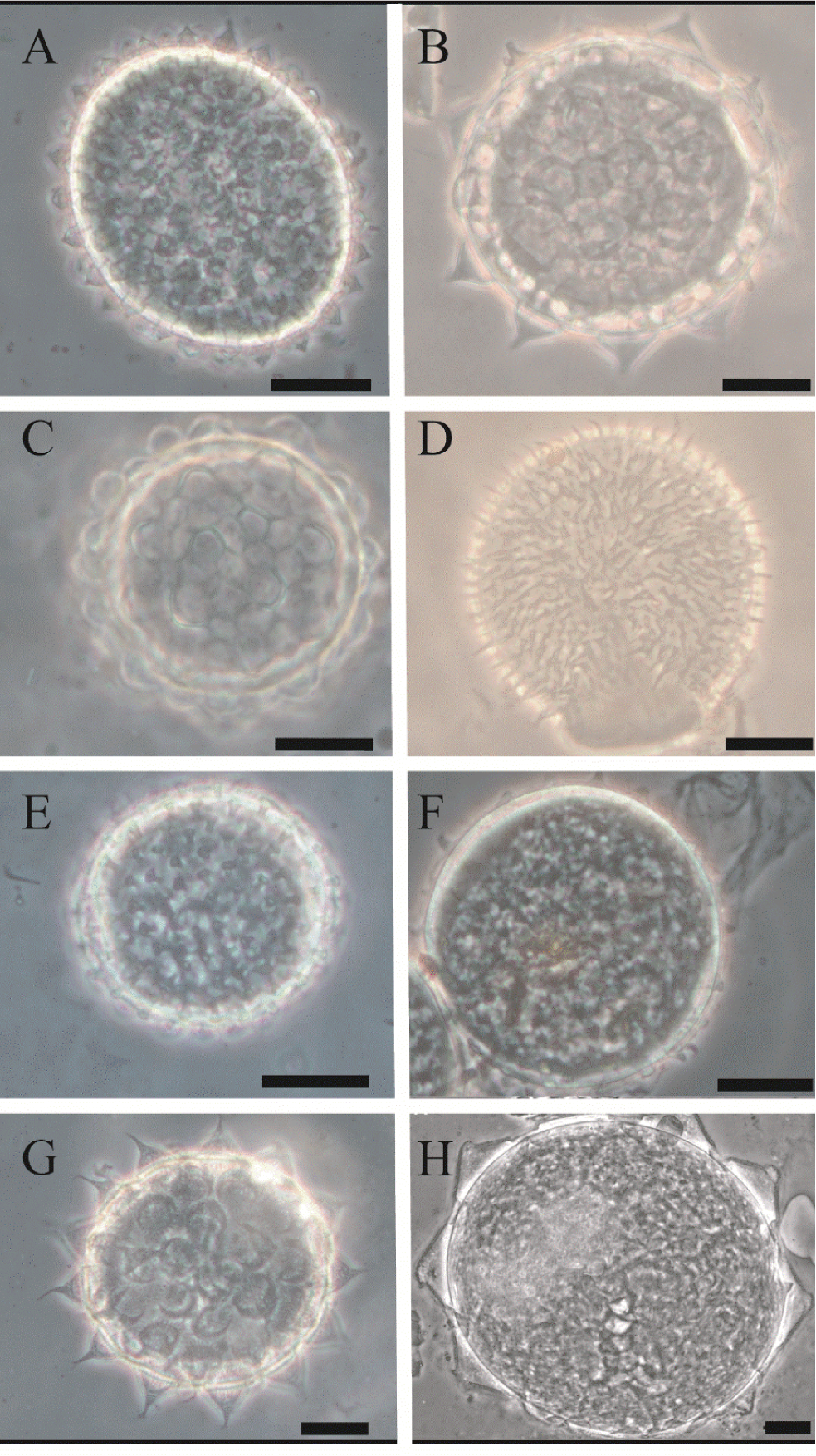
Figure 6
Eggs of the taxa identified during the study. A) Macrobiotus savai, B) Macrobiotus cf. polyopus, C) Macrobiotus cf. ocotensis, D) Macrobiotus sp. (hufelandi group), E) Minibiotus cf. intermedius, F) Macrobiotus cf. semmelweisi, G) Mesobiotus cf. pseudopatiens, H) Paramacrobiotus cf. savai. Scale 20 µm.
Eggs of the taxa identified during the study. A) Macrobiotus savai, B) Macrobiotus cf. polyopus, C) Macrobiotus cf. ocotensis, D) Macrobiotus sp. (hufelandi group), E) Minibiotus cf. intermedius, F) Macrobiotus cf. semmelweisi, G) Mesobiotus cf. pseudopatiens, H) Paramacrobiotus cf. savai. Scale 20 µm.
Macrobiotus cf. polyopus. A total of 992 adult tardigrades and 43 eggs were examined, collected from an extensive citrus plantation and an evergreen forest fragment. The morphology of both the tardigrades and the examined eggs is characteristic of the Macrobiotus polyopus group (Figure 6B). However, the shape of the egg processes differs from all known species of the group, requiring a detailed review to determine whether it represents a new species.
Macrobiotus cf. ocotensis. A single egg was examined, collected from an extensive citrus plantation. The egg diameter (excluding the processes) and the shape of the processes were similar to those documented for M. ocotensis (60 µm without processes and 80 µm with processes) (Figure 6C). However, the number of processes was lower than in M. ocotensis, with 20 processes along the circumference and 54 processes in a single hemisphere.
Macrobiotus sp. 1. A total of 899 adult individuals and four eggs were examined, collected from an intensive citrus plantation. The specimens exhibited claws, a buccal apparatus, and eggs characteristic of the hufelandi group. The buccal tube was wider than that documented in the other species in this study. The eggs were not embryonated (Figure 6D), making it difficult to associate them with the adult specimens and, consequently, preventing their identification.
Macrobiotus cf. semmelweisi. A total of 468 adults and nine eggs were examined, collected from an extensive citrus plantation. According to the identification key for the Macrobiotus hufelandi group (Kaczmarek & Michalczyk, 2017), the examined tardigrades belong to the Macrobiotus group with protuberances (Figure 4), which were similar to those documented for M. semmelweisi. However, the eggs differed in morphology from those described for this species (Figure 6F). Therefore, a thorough review is necessary to determine whether they represent a new species and to provide a formal description .
-
Kaczmarek & Michalczyk, 2017The Macrobiotus hufelandi group (Tardigrada) revisitedZootaxa, 2017
Minibiotus cf. intermedius
This species belongs to the genus Minibiotus R. O. Schuster, 1980 (Table 1). A total of 92 tardigrades and 10 eggs were examined, collected from an extensive citrus plantation. Although the observed set of characteristics in both the tardigrades and eggs corresponds to the description of M. intermedius (Plate, 1888) (Claxton, 1998; Figure 6E), it cannot be confirmed as this species. This uncertainty arises because Kaczmarek et al. (2016) classified records of M. intermedius in North America as doubtful.
-
Claxton, 1998A revision of the genus Minibiotus (Tardigrada: Macrobiotidae) with descriptions of eleven new species from AustraliaRecords of the Australian Museum, 1998
-
Kaczmarek et al. (2016)Diversity of limno-terrestrial tardigrades of the Americas in relation to the Great American Biotic Interchange hypothesis (GABI)Zoological Journal of the Linnean Society, 2016
Mesobiotus cf. pseudopatiens
This species belongs to the genus Mesobiotus Vecchi, Cesari, Bertolani, Jönsson, Rebecchi & Guidetti, 2016 (Table 1). A total of 485 individuals and 25 eggs were examined, collected from both extensive and intensive citrus plantations and an evergreen forest fragment.
Based on the identification key for Mesobiotus species by Kaczmarek et al. (2020), the specimens are close to Mesobiotus wuzhishanensis (Yin, L. Wang & X. Li, 2011) and Mesobiotus pseudopatiens Kaczmarek & Roszkowska, 2016. However, the former species is tooth-shaped in posterior lunules, while M. pseudopatiens has eyes and smooth lunules. Additionally, the egg morphology closely resembles M. pseudopatiens, a species described in Costa Rica (Figure 6G). For this reason, the specimens are more appropriately designated as Mesobiotus cf. pseudopatiens.
-
Kaczmarek et al. (2020)Revisiting the genus Mesobiotus (Eutardigrada, Macrobiotidae)-remarks, updated dichotomous key and an integrative description of new species from MadagascarZoologischer Anzeiger, 2020
Paramacrobiotus cf. savai
The genus is Paramacrobiotus Guidetti, Schill, Bertolani, Dandekar & Wolf, 2009 (Table 1). A total of 66 individuals and four eggs were examined, collected from an evergreen forest fragment.
According to the taxonomic key for Paramacrobiotus (Kaczmarek et al., 2017; Kayastha et al., 2023), the species closest to the collected specimens is P. savai (Binda & Pilato, 2001). However, these specimens differ in the smaller egg diameter without processes (90 µm in P. savai and 76.4-77.2 µm in the specimens) and the shape and size of the egg processes (12-18 µm in height in P. savai and 5.8-13.41 µm in the study specimens) (Figure 6H).
-
Kaczmarek et al., 2017The Macrobiotus hufelandi group (Tardigrada) revisitedZootaxa, 2017
-
Kayastha et al., 2023A review on the genus Paramacrobiotus (Tardigrada) with a new diagnostic keyDiversity, 2023
Ecology
A total of 4 725 tardigrades and 118 eggs, corresponding to nine species were collected from citrus plantations and an evergreen forest fragment (Tables 2 and 3). In the citrus plantation, moss and lichen were collected from the 12 selected trees during both sampling seasons. In the forest fragment, moss was collected from five trees (41 %) and lichen from four trees (33 %) in February 2018, while in September 2018, moss was collected from four trees (33 %) and lichen from six trees (50 %). For both sampled areas, the pH ranged from 5.09 to 6.80 in moss and from 4.04 to 7.36 in lichen (Table 2), the humidity ranged from 53 % to 94 %, and the temperature ranged from 26.50 °C to 36.0 °C.
Table 2
Diversity parameters of limno-terrestrial tardigrades collected in February 2018 in a citrus plantation and forest fragment in Quintana Roo.
Diversity parameters of limno-terrestrial tardigrades collected in February 2018 in a citrus plantation and forest fragment in Quintana Roo.
| Species | Citrus plantation | Forest fragment | ||
|---|---|---|---|---|
| Moss | Lichen | Moss | Lichen | |
| Macrobiotus cf. semmelweisi | 32 | 109 | 0 | 0 |
| Macrobiotus grupo hufelandi | 317 | 59 | 0 | 0 |
| Macrobiotus cf. polyopus | 524 | 28 | 0 | 2 |
| Macrobiotus alvaroi | 102 | 30 | 0 | 0 |
| Paramacrobiotus cf. savai | 0 | 0 | 4 | 0 |
| Mesobiotus cf. pseudopatiens | 76 | 51 | 0 | 10 |
| Minibiotus intermedius | 82 | 4 | 0 | 0 |
| Milnesium cf. cassandrae | 341 | 142 | 1 | 23 |
| Kristenseniscus kofordi | 212 | 11 | 0 | 0 |
| Abundancia | 1 686 | 434 | 5 | 41 |
| Riqueza | 8 | 8 | 2 | 3 |
| Número de muestras | 12 | 12 | 5 | 5 |
| Diversidad de Shannon-Wiener (H') | 1.79 | 1.73 | 0.50 | 0.79 |
| P value (t-test) | 0.07 | 0.27 | ||
Table 3
Diversity parameters of limno-terrestrial tardigrades collected in September 2018 in a citrus plantation and forest fragment in Quintana Roo.
Diversity parameters of limno-terrestrial tardigrades collected in September 2018 in a citrus plantation and forest fragment in Quintana Roo.
| Species | Citrus plantation | Forest fragment | ||
|---|---|---|---|---|
| Moss | Lichen | Moss | Lichen | |
| Macrobiotus cf. semmelweisi | 63 | 264 | 0 | 0 |
| Macrobiotus grupo hufelandi | 264 | 259 | 0 | 0 |
| Macrobiotus cf. polyopus | 331 | 107 | 0 | 0 |
| Macrobiotus alvaroi | 36 | 28 | 0 | 0 |
| Paramacrobiotus cf. savai | 0 | 0 | 1 | 61 |
| Mesobiotus cf. pseudopatiens | 98 | 209 | 1 | 29 |
| Minibiotus intermedius | 0 | 6 | 0 | 0 |
| Milnesium cf. cassandrae | 290 | 161 | 2 | 3 |
| Kristenseniscus kofordi | 265 | 81 | 0 | 0 |
| Abundance | 1 347 | 1 115 | 4 | 93 |
| Richness | 12 | 12 | 4 | 6 |
| Number of samples | 7 | 8 | 3 | 3 |
| Shannon-Wiener diversity (H') | 1.74 | 1.81 | 1.04 | 0.75 |
| P value (t-test) | 0.03 | 0.88 | ||
Nine species were recorded in both environments (citrus plantation and forest fragment) during the study. When this number was compared with the Chao1 estimator (nine species), it was determined that the sampling effort was sufficient to document all the species present in both ecosystems (Figure 7). In terms of species richness and diversity, differences were observed between the sampled areas (Tables 2 and 3), with the citrus plantation showing the highest richness and Shannon-Wiener diversity in both sampling intervals.
Thumbnail
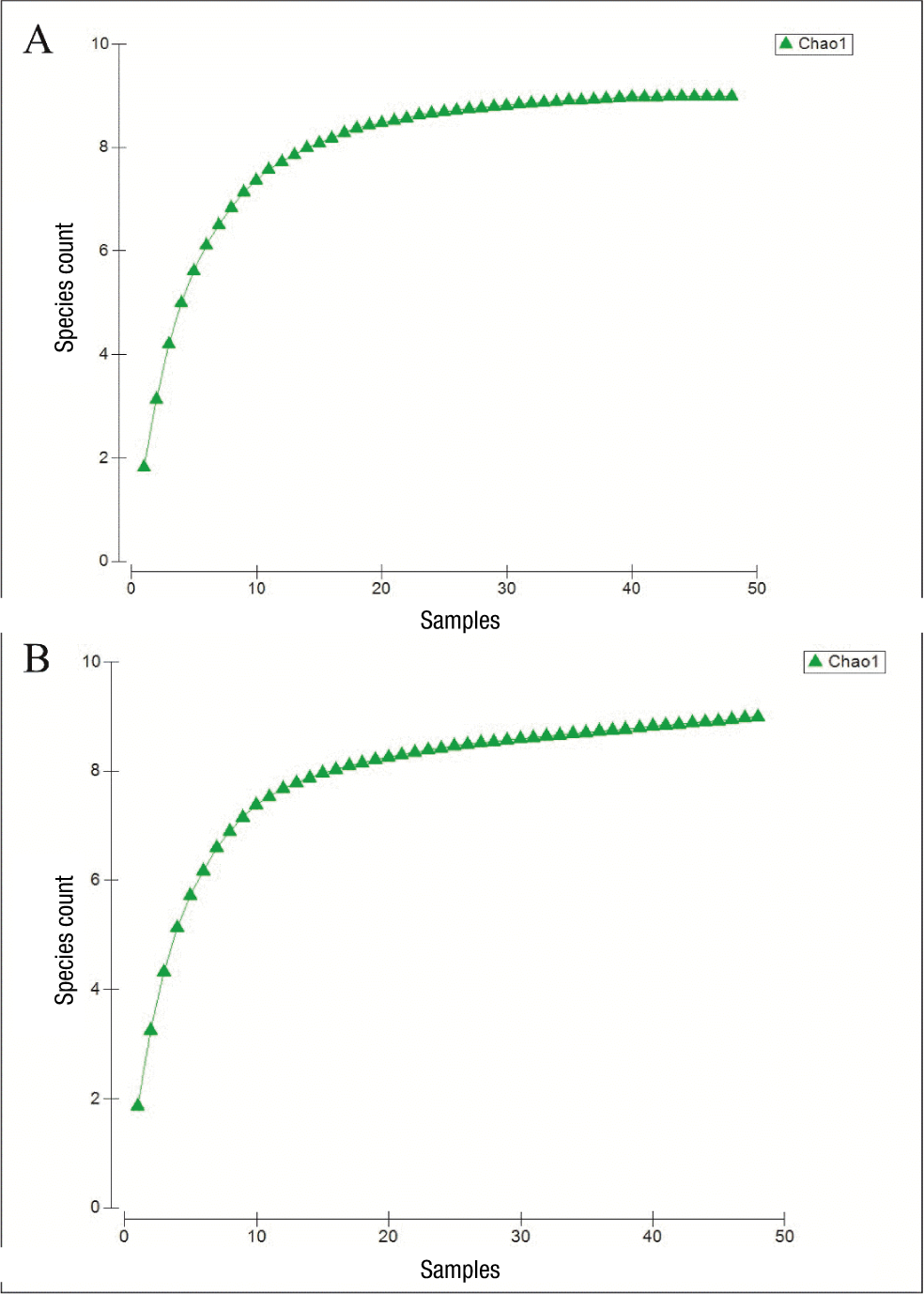
Figure 7
Accumulation curve of limno-terrestrial tardigrades species in Quintana Roo from two sampling dates: A) February 2018 and B) September 2018.
Accumulation curve of limno-terrestrial tardigrades species in Quintana Roo from two sampling dates: A) February 2018 and B) September 2018.
When the Shannon-Wiener diversity was compared between moss and lichen, in February 2018, there was no significant difference (P > 0.05). However, in September 2018, a significant difference was found (P = 0.03 < 0.05) in the citrus plantation (Table 3). On the other hand, tardigrade diversity showed variation between the study months (P < 0.05).
MDS estimated showed that diversity tended to increase toward the citrus crop during both sampling campaigns and was positively correlated with lower pH values (Figure 8A; Table 4). In September 2018, a better stratification was observed regarding the similarity of diversity among the collection sites, which was related to the type of ecosystem (citrus plantation and forest fragment; Figure 8B).
Thumbnail
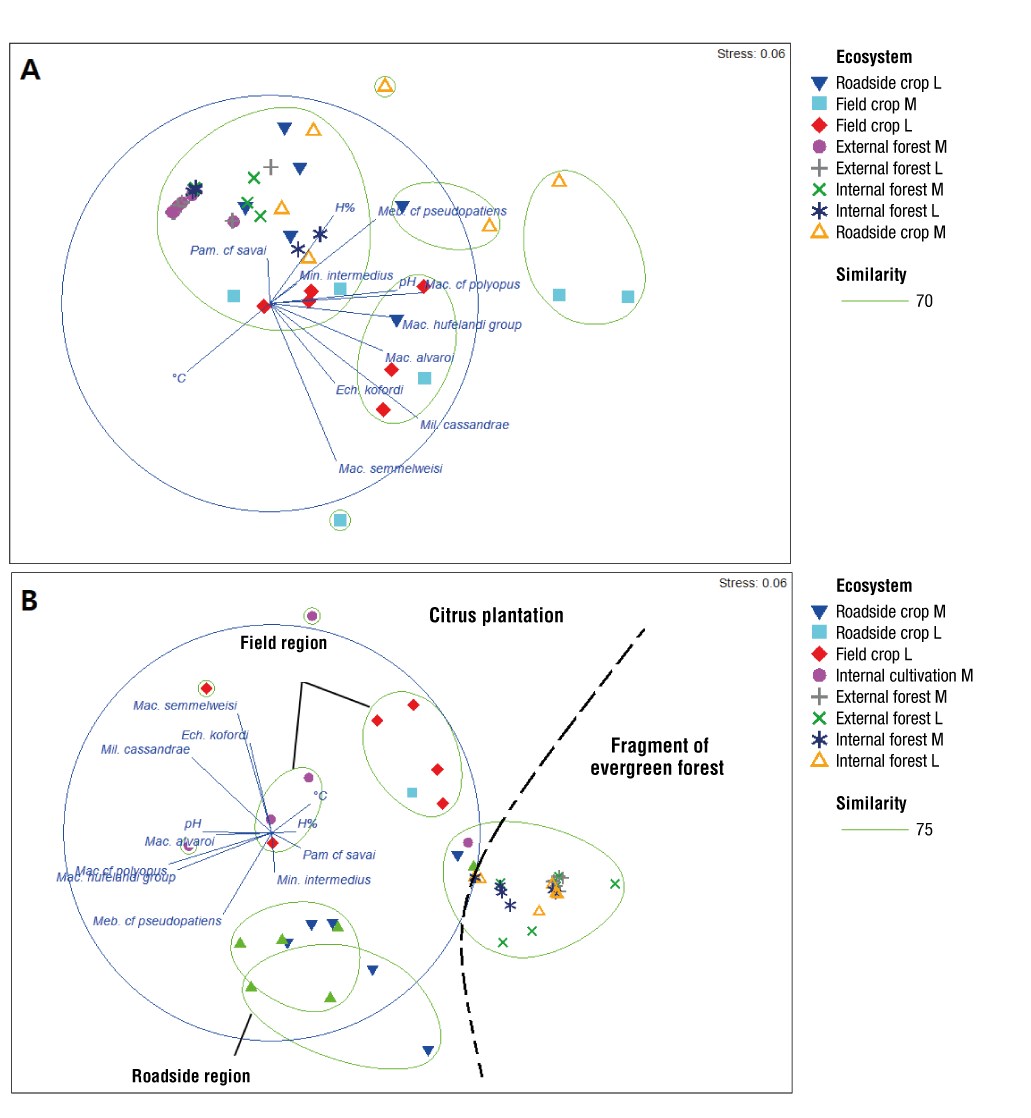
Figure 8
Multidimensional scaling analysis of pH, temperature, humidity, richness, microhabitat, and sampled environments for the collection of limno-terrestrial tardigrades on two dates: A) February 2018, B) September 2018. M: moss, L: lichen. External cultivation refers to the crop area adjacent to the unpaved road, while internal cultivation refers to the area located at the center of the cultivation site.
Multidimensional scaling analysis of pH, temperature, humidity, richness, microhabitat, and sampled environments for the collection of limno-terrestrial tardigrades on two dates: A) February 2018, B) September 2018. M: moss, L: lichen. External cultivation refers to the crop area adjacent to the unpaved road, while internal cultivation refers to the area located at the center of the cultivation site.
Table 4
Parameters measured in the sampled areas (citrus plantation and evergreen forest fragment) for the collection of limno-terrestrial tardigrades in Quintana Roo.
Parameters measured in the sampled areas (citrus plantation and evergreen forest fragment) for the collection of limno-terrestrial tardigrades in Quintana Roo.
| Parameter | Strata | February 2018 | September 2018 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Minimum | Maximum | Man | SD | Minimum | Maximum | Mean | SD | ||
| pH | Moss - citrus | 5.09 | 5.90 | 5.49 | 0.28 | 5.34 | 6.36 | 5.71 | 0.32 |
| Moss - forest | 5.80 | 6.38 | 6.04 | 0.24 | 6.03 | 6.80 | 6.39 | 0.33 | |
| Lichen - citrus | 4.04 | 5.30 | 4.93 | 0.34 | 4.89 | 6.36 | 5.27 | 0.38 | |
| Lichen - forest | 5.10 | 6.79 | 6.14 | 0.70 | 5.62 | 7.36 | 6.34 | 0.68 | |
| Humedy (%) | Moss - citrus | 53.00 | 94.00 | 71.33 | 14.24 | 57.00 | 76.00 | 69.83 | 5.47 |
| Moss - forest | 57.00 | 76.00 | 67.92 | 7.27 | 64.00 | 90.00 | 77.08 | 8.39 | |
| Lichen - citrus | 53.00 | 94.00 | 71.33 | 14.24 | 57.00 | 76.00 | 69.83 | 5.47 | |
| Lichen - forest | 57.00 | 76.00 | 67.73 | 7.27 | 64.00 | 90.00 | 77.08 | 8.39 | |
| °C | Moss- citrus | 26.50 | 32.00 | 29.68 | 2.11 | 28.50 | 34.90 | 31.58 | 2.14 |
| Moss - forest | 26.40 | 34.50 | 29.94 | 3.50 | 29.00 | 36.00 | 31.67 | 2.39 | |
| Lichen - citrus | 26.50 | 32.00 | 29.68 | 2.11 | 28.50 | 34.90 | 31.58 | 2.14 | |
| Lichen - forest | 26.40 | 34.50 | 29.88 | 3.50 | 29.00 | 36.00 | 31.67 | 2.39 | |
Discussion
Currently, the cosmopolitan distribution of certain Tardigrada species is considered questionable (Gąsiorek, 2023; Kaczmarek et al., 2016; Morek et al., 2021); indeed, Mogle et al. (2018) proposed that, in the Americas, the distribution of limno-terrestrial tardigrades over large distances may be mediated by migratory birds. Gąsiorek (2023) established that the hypothesis “everything is everywhere, but the environment selects” as a starting point for the biogeography of limno-terrestrial tardigrades has come to an end and has been replaced by a regional diversity approach.
-
Gąsiorek, 2023Catch me if you can, or how paradigms of tardigrade biogeography evolved from cosmopolitism to ‘localism’Zoological Journal of the Linnean Society, 2023
-
Kaczmarek et al., 2016Diversity of limno-terrestrial tardigrades of the Americas in relation to the Great American Biotic Interchange hypothesis (GABI)Zoological Journal of the Linnean Society, 2016
-
Morek et al., 2021“Everything is not everywhere”: Time‐calibrated phylogeography of the genus Milnesium (Tardigrada)Molecular Ecology, 2021
-
Mogle et al. (2018)Evidence of avian-mediated long distance dispersal in American tardigradesPeerJ, 2018
-
Gąsiorek (2023)Catch me if you can, or how paradigms of tardigrade biogeography evolved from cosmopolitism to ‘localism’Zoological Journal of the Linnean Society, 2023
Tardigrades are a poorly studied phylum worldwide. The number of species and the rate of their description is limited, which is reflected in the diversity estimates of the group, classifying it as species-poor (Bartels et al., 2016). Today, the systematics of the group is based on a limited number of morphological traits for species delimitation, which is why many of these species are grouped into complexes, and their cryptic diversity is only revealed through molecular studies (Gąsiorek, 2023). In this study, 66 % of the identified taxa were found to be similar to other species (Milnesium cf. cassandrae, Macrobiotus cf. polyopus, Macrobiotus cf. ocotensis, Macrobiotus cf. semmelweisi, Minibiotus cf. intermedius, Paramacrobiotus cf. savai and Ramazzottius cf. baumanni) and were considered morphospecies (Milnesium sp. 1 and Macrobiotus sp. 1). These taxa are likely new species for science. However, integrative taxonomy methods and molecular sequences are necessary to describe them (Gąsiorek, 2023).
-
Bartels et
al., 2016A global biodiversity estimates of a poorly known taxon: phylum TardigradaZoological Journal of the Linnean Society, 2016
-
Gąsiorek, 2023Catch me if you can, or how paradigms of tardigrade biogeography evolved from cosmopolitism to ‘localism’Zoological Journal of the Linnean Society, 2023
-
Gąsiorek, 2023Catch me if you can, or how paradigms of tardigrade biogeography evolved from cosmopolitism to ‘localism’Zoological Journal of the Linnean Society, 2023
M. ocotensis was described for the state of Chiapas; however, it was not possible to confirm that our specimens belong to the same species due to the variations observed in the number of processes in the single egg collected (Pilato, 2006). In the case of M. cassandrae, its distribution in Quintana Roo could be assumed, as it is a Mexican species. It is distributed in northern Mexico, an area biogeographically located in the Nearctic, which has shown differentiated diversity from the Neotropics, the region where the study area of this work is located (Kaczmarek et al., 2016). Therefore, it would be premature to assert that this species is distributed in the study area. Although the genus Milnesium was long considered to consist of cosmopolitan species, the advent of molecular biology has cast doubt on this assumption (Gąsiorek, 2023). In 2021, Morek et al. determined that Milnesium has limited dispersal capabilities, and therefore its species are likely restricted to unique zoogeographical regions. In this regard, only the integration of molecular analysis into the morphological description of the collected specimens of Macrobiotus cf. ocotensis and Milnesium cf. cassandrae could confirm the presence of these species in Quintana Roo or support the description of new species for science.
-
Pilato, 2006Remarks on the Macrobiotus polyopus group, with the description of two new species (Eutardigrada, Macrobiotidae)Zootaxa, 2006
-
Kaczmarek et al., 2016Diversity of limno-terrestrial tardigrades of the Americas in relation to the Great American Biotic Interchange hypothesis (GABI)Zoological Journal of the Linnean Society, 2016
-
Gąsiorek, 2023Catch me if you can, or how paradigms of tardigrade biogeography evolved from cosmopolitism to ‘localism’Zoological Journal of the Linnean Society, 2023
On the other hand, the species K. kofordi and M. alvaroi, previously recorded in Chiapas (Dueñas-Cedillo et al., 2020; García-Román et al., 2022), represent new records for Quintana Roo, contributing to the knowledge of the distribution of these species in Mexico. The addition of the taxa documented here, along with the previous records of Echiniscus sp. and D. chetumalensis, brings the total number of species on the list of limno-terrestrial tardigrades in Quintana Roo to 14 (Pérez-Pech et al., 2016, 2017) (Table 1). Additionally, the coastal mangrove forest is added to the list of environments for these animals in the state. Previously, limno-terrestrial tardigrades had only been reported in citrus crops (Anguas-Escalante et al., 2018; Cutz-Pool et al., 2019) and accumulated sediment along the asphalt streets of the city of Chetumal (Pérez-Pech et al., 2016, 2017).
-
Dueñas-Cedillo et al., 2020Tardigrades from Iztaccíhuatl Volcano (Trans-Mexican Volcanic Belt), with the description of Minibiotus citlalium sp. nov. (Eutardigrada: Macrobiotidae)Diversity, 2020
-
García-Román et al., 2022A strategy to provide a present and future scenario of Mexican biodiversity of tardigradaDiversity, 2022
-
Pérez-Pech et al., 2016Primer registro genérico de tardígrados, habitantes del área urbana de Chetumal Quintana Roo, MéxicoEntomología Mexicana, 2016
-
2017Doryphoribius chetumalensis sp. nov. (Eutardigrada: Isohypsibiidae) a new tardigrade species discovered in an unusual habitat of urban areas of MexicoZootaxa, 2017
-
Anguas-Escalante et al., 2018Tardígrados asociados a una plantación de cítricos de traspatio en la comunidad de El Palmar en Quintana Roo, MéxicoInvestigación y Ciencia, 2018
-
Cutz-Pool
et al., 2019Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, MéxicoAgroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria, 2019
-
Pérez-Pech et al., 2016Primer registro genérico de tardígrados, habitantes del área urbana de Chetumal Quintana Roo, MéxicoEntomología Mexicana, 2016
-
2017Doryphoribius chetumalensis sp. nov. (Eutardigrada: Isohypsibiidae) a new tardigrade species discovered in an unusual habitat of urban areas of MexicoZootaxa, 2017
The 14 limno-terrestrial tardigrade taxa for Quintana Roo, along with the record of marine tardigrade species, bring the total to 28 species (Anguas-Escalante et al., 2020; Pérez-Pech et al., 2020). This study also provides images of the tardigrade eggs collected, which are important elements for species-level identification (Kaczmarek et al., 2017, 2020; Kaczmarek & Michalczyk, 2017; Morek et al., 2016). This information is crucial for use in future studies in the Yucatán Peninsula.
-
Anguas-Escalante et al., 2020A new species of Tardigrada from a Caribbean reef lagoon, Florarctus yucatanensis sp. nov. (Halechiniscidae: Florarctinae)CBM - Cahiers de Biologie Marine, 2020
-
Pérez-Pech et al., 2020Marine Tardigrada from the Mexican Caribbean with the description of Styraconyx robertoi sp. nov. (Arthrotardigrada: Styraconyxidae)Zootaxa, 2020
-
Kaczmarek et al., 2017The Macrobiotus hufelandi group (Tardigrada) revisitedZootaxa, 2017
-
2020Revisiting the genus Mesobiotus (Eutardigrada, Macrobiotidae)-remarks, updated dichotomous key and an integrative description of new species from MadagascarZoologischer Anzeiger, 2020
-
Kaczmarek
& Michalczyk, 2017The Macrobiotus hufelandi group (Tardigrada) revisitedZootaxa, 2017
-
Morek et al.,
2016Experimental taxonomy exposes ontogenetic variability and elucidates the taxonomic value of claw configuration in Milnesium Doyère, 1840 (Tardigrada: Eutardigrada: Apochela)Contributions to Zoology, 2016
Regarding the ecology of urban and rural tardigrades, Meyer et al. (2013) suggest that environments created in the Anthropocene may support the maintenance of biodiversity with a unique composition. This was observed in the present study, as it was found that citrus crops (regardless of the biotope type), an environment altered and modified by human action, can harbor high levels of tardigrade richness, diversity, and abundance compared to vegetation fragments. In terms of diversity (H´), there was generally no variation between lichen and moss samples from the same plots, with variation only observed during the September 2018 sampling in the plantation area. This agrees with findings by Dastych (1988) in Poland, who discovered that rather than ecosystem type (e.g., forest or soil types), the microenvironment type was the factor most related to the variation in the diversity and abundance of limno-terrestrial tardigrades, not the bryophyte species to which they were associated. This has also been observed in boreal forests in Norway, where tardigrade communities varied more regarding the substrate (leaf litter, lichen, and bryophytes) than with the type of ecosystem (Guidetti et al., 2024).
-
Meyer et al. (2013)Water bears in the Anthropocene: a comparison of urban and woodland tardigrade (Phylum Tardigrada) communities in Southwestern Louisiana, USAJournal of Limnology, 2013
-
Dastych (1988)The Tardigrada of Poland, 1988
-
Guidetti et al., 2024Tardigrade diversity and community composition across Norwegian boreal forestsZoological Journal of the Linnean Society, 2024
This study also documented differences in the availability of habitats for tardigrades in the environments studied. In the case of citrus, the availability of lichen and moss was 100 % during both sampling periods, while in the forest fragment, the presence of lichen and moss was recorded in 33 % and 55 % of the sampled trees, respectively. Therefore, it is possible that the citrus crop provides greater habitat availability for tardigrades compared to the vegetation fragment. In Costa Rica, Kaczmarek et al. (2011) found similar results when comparing modified environments to natural ones and concluded that the former may harbor greater diversity. However, the researchers demonstrated that this fact was strongly correlated with humidity (and less so with plant and substrate types). In contrast, in this study, the MDS values showed little correlation between the structure of the tardigrade community and humidity levels. On the other hand, when analyzing the MDS results between physical (humidity and temperature), chemical (pH) parameters, and diversity, it was found that pH correlated better with species arrangement. These results agree with those of Utsugi (1984) and Johansson et al. (2011), who mentioned that pH is a parameter that drives the variation in limno-terrestrial tardigrade diversity. Utsugi (1984) considered that tardigrades prefer environments with a neutral pH (6.5-8.0), and Johansson et al. (2011) found that tardigrades in rural environments prefer slightly acidic pH levels (4.5 to 6.0); in this study, the values were slightly acidic and neutral (pH 5.1-7.4).
-
Kaczmarek et al. (2011)Ecological factors determining Tardigrada distribution in Costa RicaJournal of Zoological Systematics and Evolutionary Research, 2011
-
Utsugi (1984)Tardigrades found in the mosses of cities of JapanZoological Science, 1984
-
Johansson et al. (2011)Are urban and rural tardigrade (Tardigrada) communities distinct and determined by pH: a case study from Fresno County, CaliforniaThe Pan-Pacific Entomologist, 2011
-
Utsugi (1984)Tardigrades found in the mosses of cities of JapanZoological Science, 1984
-
Johansson et al. (2011)Are urban and rural tardigrade (Tardigrada) communities distinct and determined by pH: a case study from Fresno County, CaliforniaThe Pan-Pacific Entomologist, 2011
On the other hand, considering the postulation by Meyer et al. (2013) regarding the uniqueness of the tardigrade community in anthropogenic environments, it is important to add that some taxa may adapt better to these environments and, in turn, show high dominance. This could also influence the determination of the tardigrade community structure in rural environments, as documented by Gonzales-Reyes et al. (2016), who, like in the present study, identified the genus Macrobiotus as dominant. The results showed clear differentiation of tardigrade communities between the two environments in September 2018, with significant difference (P < 0.05) in overall diversity by study month. This demonstrates that the diversity of limno-terrestrial tardigrades is affected by time. This study covered the rainy season and the 'nortes' season; therefore, the observed changes in diversity between the studied environments and time periods could be affected by changes in climatic variables.
-
Meyer et al. (2013)Water bears in the Anthropocene: a comparison of urban and woodland tardigrade (Phylum Tardigrada) communities in Southwestern Louisiana, USAJournal of Limnology, 2013
Conclusions
The updated list of limno-terrestrial tardigrades in Quintana Roo consists of 14 species; furthermore, the coastal mangrove forest is added to the list of environments for these animals in the region. The richness and Shannon-Wiener diversity were higher in the citrus plantation than in the evergreen forest fragment, suggesting that tardigrades adapt well to anthropogenic environments. The results indicate that the patterns of tardigrade diversity in modified and natural environments are influenced by multifactorial processes. At the landscape level, the type of ecosystem is a factor that determines habitat availability for tardigrades. Additionally, microenvironmental conditions such as pH, and to a lesser extent humidity, also influence the structure and composition of limno-terrestrial tardigrade communities.
Acknowledgments
The first author thanks the Secretaría de Ciencia, Humanidades, Tecnología e Innovación for the funding provided during the development of the study (CVU 77783). The fifth author thanks the institutional project 5805.19-P from the Tecnológico Nacional de México/Instituto Tecnológico de Chetumal for the funding provided. The authors also thank Dr. Alberto de Jesús Navarrete and the Colegio de la Frontera Sur for providing the facilities and equipment from Laboratorio de Pesquerías Sección Meiofauna, during the identification of the taxa; thanks to M. C. Héctor Ortiz León for the support given at the Laboratorio de Zoología del Tecnológico Nacional de México, campus Chetumal, during sample separation; and to the anonymous reviewers for their feedback on this article.
References
- Anguas-Escalante, A., de Jesús-Navarrete, A., Demilio, E., Perez-Pech, W. A., & Hansen, J. G. (2020). A new species of Tardigrada from a Caribbean reef lagoon, Florarctus yucatanensis sp. nov. (Halechiniscidae: Florarctinae).CBM - Cahiers de Biologie Marine, 61, 377‒385. https://doi.org/10.21411/CBM.A.CD1B185A Links
- Anguas-Escalante, A., Pérez-Pech, W. A., Guidetti, R., Cutz-Pool, L. Q., & Ortiz León, H. (2018). Tardígrados asociados a una plantación de cítricos de traspatio en la comunidad de El Palmar en Quintana Roo, México. Investigación y Ciencia, 73, (26), 20-26. https://doi.org/10.33064/iycuaa201873203 Links
- Bartels, P. J., Apodaca, J. J., Mora, C., & Nelson, D. R. (2016). A global biodiversity estimates of a poorly known taxon: phylum Tardigrada. Zoological Journal of the Linnean Society, 178(4), 730-736. https://doi.org/10.1111/zoj.12441 Links
- Clarke, K. R., & Gorley, R. N. (2006). PRIMER V6: User Manual/tutorial. PRIMER-E. Plymouth. Links
- Claxton, S. K. (1998). A revision of the genus Minibiotus (Tardigrada: Macrobiotidae) with descriptions of eleven new species from Australia. Records of the Australian Museum, 50(2), 125-160. https://doi.org/10.3853/j.0067-1975.50.1998.1276 Links
- Cutz-Pool, L., Crisanto, J., Pérez-Pech, W. A., Anguas-Escalante, A., & Guidetti, R. (2019). Caracterización de la fauna de tardígrados (Ecdysozoa: Tardigrada) de liquen y musgo en dos sitios con diferente uso de suelo en Quintana Roo, México. En W. Cetzal-Ix, F. Casanova-Lugo, A. J. Chay-Canul, & J. F. Martínez-Puc (Eds.), Agroecosistemas tropicales: Conservación de recursos naturales y seguridad alimentaria (1.a ed., pp. 193-200). Tecnológico Nacional de México, Instituto Tecnológico de Chiná - Instituto Tecnológico de la Zona Maya. Links
- Dastych, H. (1988). The Tardigrada of Poland. Wyd. Naukowe. Links
- Dueñas-Cedillo, A., Martínez-Méndez, E., García-Román, J., Armendáriz-Toledano, F., & Ruiz, E. (2020). Tardigrades from Iztaccíhuatl Volcano (Trans-Mexican Volcanic Belt), with the description of Minibiotus citlalium sp. nov. (Eutardigrada: Macrobiotidae). Diversity, 12(271), 2‒24. https://doi.org/10.3390/d12070271 Links
- Gąsiorek, P. (2023). Catch me if you can, or how paradigms of tardigrade biogeography evolved from cosmopolitism to ‘localism’. Zoological Journal of the Linnean Society, 202(2), zlad191. https://doi.org/10.1093/zoolinnean/zlad191 Links
- García-Román, J., Dueñas-Cedillo, A., Cervantes-Espinoza, M., Flores-Martínez, J. J., Vargas-Mendoza, C. F., Ruiz, E. A., & Armendáriz-Toledano, F. (2022). A strategy to provide a present and future scenario of Mexican biodiversity of tardigrada. Diversity, 14(4), 280. https://doi.org/10.3390/d14040280 Links
- González-Reyes, A., Acosta, X., Corronca, J., Rocha, M., Doma I., & Repp, E. Y. (2016). Evaluación de la diversidad en comunidades de tardígrados (Ecdysozoa, Tardigrada) en hábitats urbano y rural de la ciudad de Salta (Argentina). Iheringia Série Zoologia, 106, 1‒8. https://doi.org/10.1590/1678-4766e2016026 Links
- Guidetti, R., Cesari, M., Bertolani, R., Altiero, T., & Rebecchi, L. (2019). High diversity in species, reproductive modes and distribution within the Paramacrobiotus richtersi complex (Eutardigrada, Macrobiotidae). Zoological Letters, 5(1), 2‒28. https://doi.org/10.1186/s40851-018-0113-z Links
- Guidetti, R., Peluffo, J. R., Rocha, A. M., Cesari, M., & de Peluffo, M. C. M. (2013). The morphological and molecular analyses of a new South American urban tardigrade offer new insights on the biological meaning of the Macrobiotus hufelandi group of species (Tardigrada: Macrobiotidae). Journal of Natural History, 47, 2409‒2426. https://doi.org/10.1080/00222933.2013.800610 Links
- Guidetti, R., Ingemar Jönsson, K., Kaczmarek, Ł., Meier, T., Speed, J. D., Prestø, T., Stur, E. Topstad, L., Cesari, M., Roszkowska, M., Zawierucha, K., hassel, K., & Ekrem, T. (2024). Tardigrade diversity and community composition across Norwegian boreal forests. Zoological Journal of the Linnean Society, 200(1), 156‒171. https://doi.org/10.1093/zoolinnean/zlad136 Links
- Hammer, O., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological statistics software package education and analysis. Paleontologia Electronica, 4(9), 2‒9. https://palaeo-electronica.org/2001_1/past/past.pdf Links
- Johansson, C., Calloway, S., Miller, W. R., & Linder, E. T. (2011). Are urban and rural tardigrade (Tardigrada) communities distinct and determined by pH: a case study from Fresno County, California. The Pan-Pacific Entomologist, 87, 86‒98. https://doi.org/10.3956/2010-27.1 Links
- Kaczmarek, Ł., Bartylak, T., Stec, D., Kulpa, A., Kepel, M., Kepel, A., & Roszkowska, M. (2020). Revisiting the genus Mesobiotus (Eutardigrada, Macrobiotidae)-remarks, updated dichotomous key and an integrative description of new species from Madagascar. Zoologischer Anzeiger, 287, 121‒146. https://doi.org/10.1016/j.jcz.2020.05.003 Links
- Kaczmarek, Ł., Gawlak, M., Bartels P. J., Nelson, D. R., & Roszkowska, M. (2017). Revision of the genus Paramacrobiotus Guidetti et al., 2009 with the description of a new species, re-descriptions and a key. Annales Zoologici, 67, 627‒657. https://doi.org/10.3161/00034541ANZ2017.67.4.001 Links
- Kaczmarek, Ł., Gołdyn, B., Mcinnes, S. J., & Michalczyk, Ł. (2016). Diversity of limno-terrestrial tardigrades of the Americas in relation to the Great American Biotic Interchange hypothesis (GABI). Zoological Journal of the Linnean Society, 178(4), 737‒746. https://doi.org/10.1111/zoj.12422 Links
- Kaczmarek, Ł., Gołdyn, B., Wełnicz, W., & Michalczyk, Ł. (2011). Ecological factors determining Tardigrada distribution in Costa Rica. Journal of Zoological Systematics and Evolutionary Research, 49, 78-83. https://doi.org/10.1111/j.1439-0469.2010.00603.x Links
- Kaczmarek, Ł., & Michalczyk, Ł. (2017). The Macrobiotus hufelandi group (Tardigrada) revisited. Zootaxa, 4363, 101‒123. https://doi.org/10.11646/zootaxa.4363.1.4 Links
- Kayastha, P., Mioduchowska, M., Warguła, J., & Kaczmarek, Ł. (2023). A review on the genus Paramacrobiotus (Tardigrada) with a new diagnostic key. Diversity, 15(9), 977. https://doi.org/10.3390/d15090977 Links
- Meyer, H. A., Hinton, J. G., & Samletzka, C. A. (2013). Water bears in the Anthropocene: a comparison of urban and woodland tardigrade (Phylum Tardigrada) communities in Southwestern Louisiana, USA. Journal of Limnology, 72, 123-127. https://doi.org/10.4081/jlimnol.2013.s1.e15 Links
- Mogle, M. J., Kimball, S. A., Miller, W. R., & McKown, R. D. (2018). Evidence of avian-mediated long distance dispersal in American tardigrades. PeerJ, 6, e5035. https://doi.org/10.7717/peerj.5035 Links
- Morek, W., Gąsiorek, P., Stec, D., Blagden, B., & Michalczyk, Ł. (2016). Experimental taxonomy exposes ontogenetic variability and elucidates the taxonomic value of claw configuration in Milnesium Doyère, 1840 (Tardigrada: Eutardigrada: Apochela). Contributions to Zoology, 85(2), 173-200. https://doi.org/10.1163/18759866-08502003 Links
- Morek, W., Surmacz, B., López‐López, A., & Michalczyk, Ł. (2021). “Everything is not everywhere”: Time‐calibrated phylogeography of the genus Milnesium (Tardigrada). Molecular Ecology, 30(14), 3590‒3609. https://doi.org/10.1111/mec.15951 Links
- Morek, W., Wałach, K., & Michalczyk, Ł. (2022). Rough backs: taxonomic value of epicuticular sculpturing in the genus Milnesium Doyère, 1840 (Tardigrada: Apochela). Scientific Reports, 12(1), 9857. https://doi.org/10.1038/s41598-022-10758-z Links
- Moreno-Talamantes, A., Roszkowska, M., Ríos Guayasamín, P., Flores Maldonado, J. J., & Kaczmarek, Ł. (2015). First record of Dactylobiotus parthenogeneticus Bertolani, 1982 (Eutardigrada: Murrayidae) in Mexico. Check List, 11(4), 1723. https://doi.org/10.15560/11.4.1723 Links
- Moreno-Talamantes, A., Roszkowska, M., García-Aranda, M. A., Flores-Maldonado, J. J., & Kaczmarek, Ł. (2019). Current knowledge on Mexican tardigrades with a description of Milnesium cassandrae sp. nov. (Eutardigrada: Milnesiidae) and discussion on the taxonomic value of dorsal pseudoplates in the genus Milnesium Doyère, 1840. Zootaxa, 4695, 501‒524. https://doi.org/10.11646/zootaxa.4691.5.5 Links
- Moreno-Talamantes A., & León-Espinosa G. (2019). Nuevo registro de Diaforobiotus islandicus (Richters, 1904) (EUTARDIGRADA: RICHTERSIIDAE) para México. Arido-Ciencia, 6(1), 5‒12. http://fcbujed.com/aridociencia/numeros/2019/VIVN1/articulo2.pdf Links
- Nelson, D., Guidetti, R., & Rebecchi, L. (2015). Phylum Tardigrada. En J. Thorp, & D. C. Rogers (Eds.), Freshwater invertebrates (vol. I 4th ed., pp. 347-380). San Diego: Academic Press (Elsevier Inc.). https://www.sciencedirect.com/book/9780123850263/thorp-and-covichs-freshwater-invertebrates Links
- Núñez, P. G., León-Espinosa, G. A., Vázquez, R., Peña-Salinas, M. E., Rodríguez-Almaraz, G. A., & Moreno-Talamantes, A. (2021). First tardigrade records from San Pedro Mártir, Baja California, Mexico. Check List, 17, 1131. https://doi.org/10.15560/17.4.1131 Links
- Pérez-Pech, W. A., Anguas-Escalante, A., Cutz-Pool, L. Q., & Guidetti, R. (2017). Doryphoribius chetumalensis sp. nov. (Eutardigrada: Isohypsibiidae) a new tardigrade species discovered in an unusual habitat of urban areas of Mexico. Zootaxa, 4344, 345‒356. https://doi.org/10.11646/zootaxa.4344.2.9 Links
- Pérez-Pech, W. A., Cutz-Pool, L. Q., Guidetti, R., & Blanco-Piñón, A. (2016). Primer registro genérico de tardígrados, habitantes del área urbana de Chetumal Quintana Roo, México. Entomología Mexicana, 3, 912-918. https://www.acaentmex.org/entomologia/revista/2016/SM/Em%20912-918.pdf Links
- Perez-Pech, W. A., Demilio, E., Anguas-Escalante, A., & Hansen, J. G. (2020). Marine Tardigrada from the Mexican Caribbean with the description of Styraconyx robertoi sp. nov. (Arthrotardigrada: Styraconyxidae). Zootaxa, 4731(4). https://doi.org/10.11646/zootaxa.4731.4.3 Links
- Pilato, G. (2006). Remarks on the Macrobiotus polyopus group, with the description of two new species (Eutardigrada, Macrobiotidae). Zootaxa, 1298(1), 37‒47. https://doi.org/10.5281/zenodo.5616731 Links
- Pilato, G., & Kaczmarek, Ł. (2007). Macrobiotus alvaroi, a new species of eutardigrade (Tardigrada, Macrobiotidae) of the polyopus group from Costa Rica (Central America).Zootaxa,1479(1), 1‒7. https://doi.org/10.11646/zootaxa.1479.1.1 Links
- Utsugi, K. (1984). Tardigrades found in the mosses of cities of Japan. Zoological Science 1, 999. Links